

Effect of Gracilaria vermiculophylla Macroalga on Non-Alcoholic Fatty Liver Disease in Obese Rats

 ,
,  , ,
, ,  ,
,  and
and
Abstract
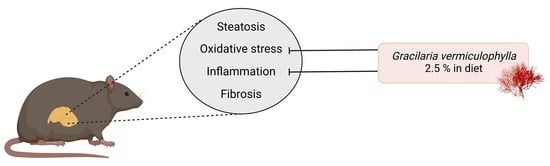
1. Introduction
2. Materials and Methods
2.1. Macroalga Obtention and Characterisation
2.2. Animals, Diets and Experimental Design
2.3. Serum Parameters
2.4. Hepatic Lipid Content
2.5. Enzyme Activities in Liver
2.6. Western Blotting for Protein Expression Measurement
2.7. Gene Expression by Real-Time PCR
2.8. Parameters Related to Oxidative Stress in Liver
2.9. Statistical Analysis
3. Results
3.1. Macroalga Composition
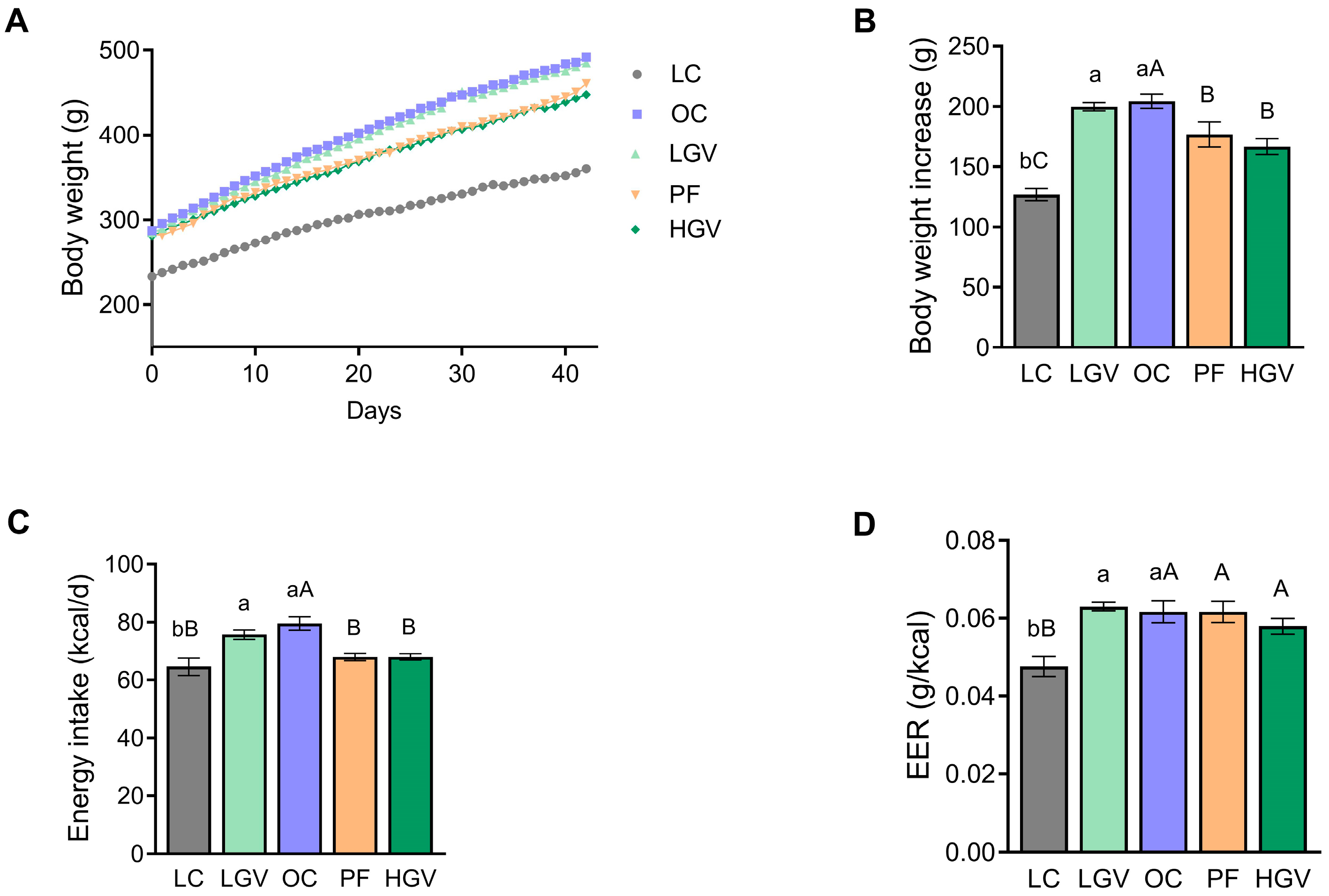
3.2. Body Weight, Energy Intake and Energy Efficiency Ratio
3.3. Serum Biochemical Parameters
3.4. Liver Weight and Hepatic Index
3.5. Hepatic Lipid Content
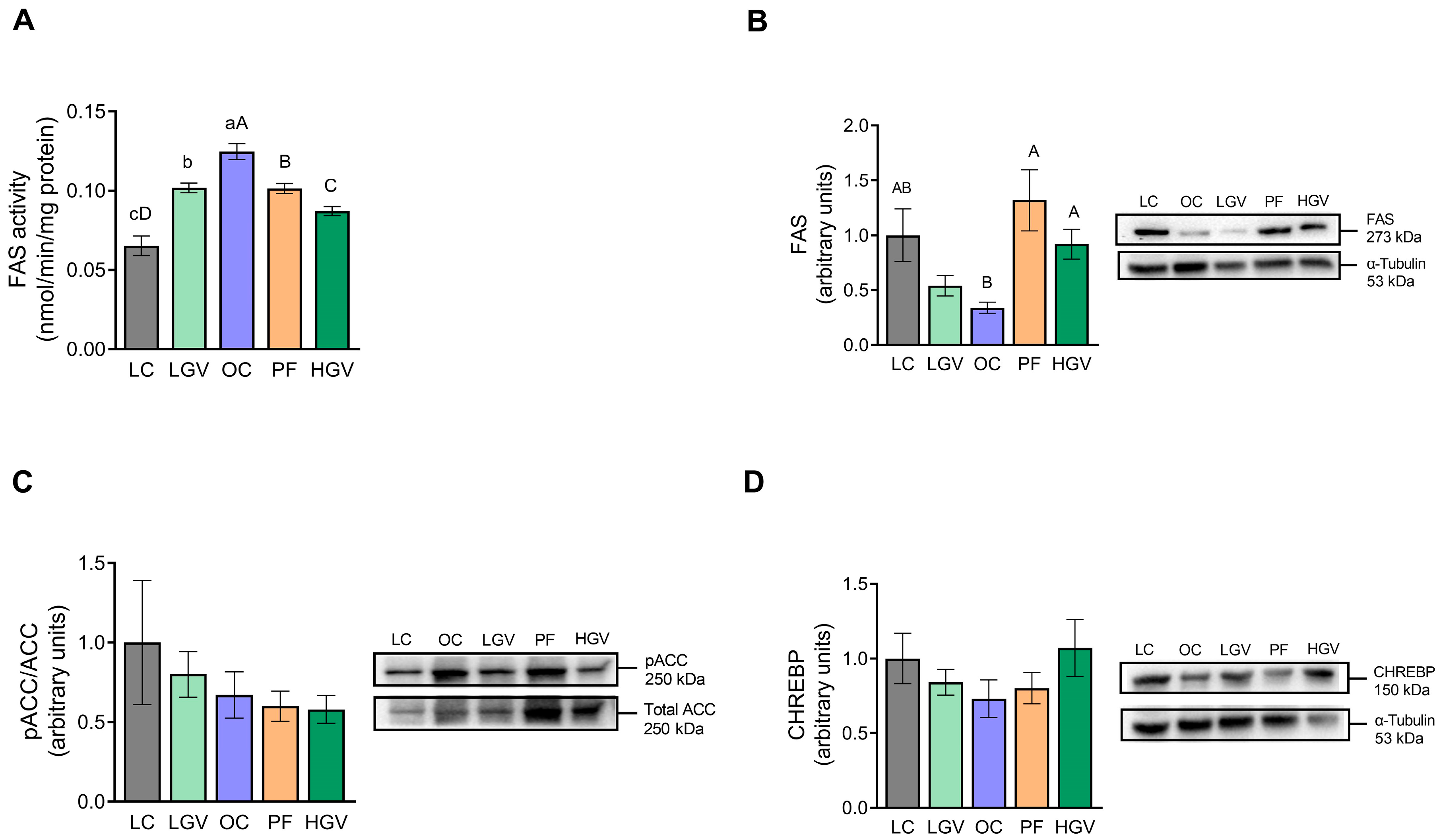
3.6. Activities of Enzymes and Expression of Proteins Involved in Fatty Acid and Triglyceride Synthesis
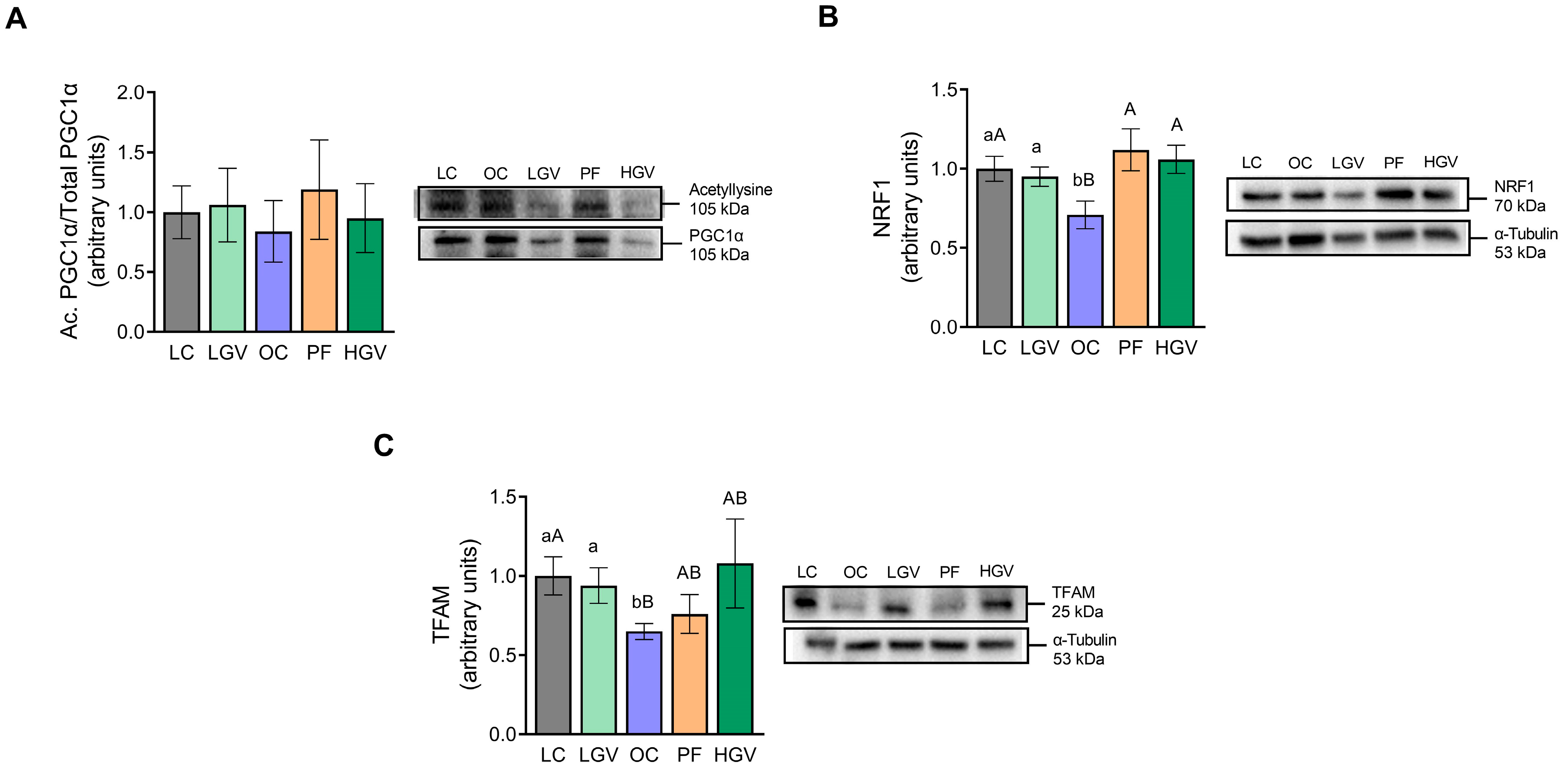
3.7. Activities of Enzymes and Expression of Proteins Involved in Fatty Acid Oxidation and Mitochondriogenesis
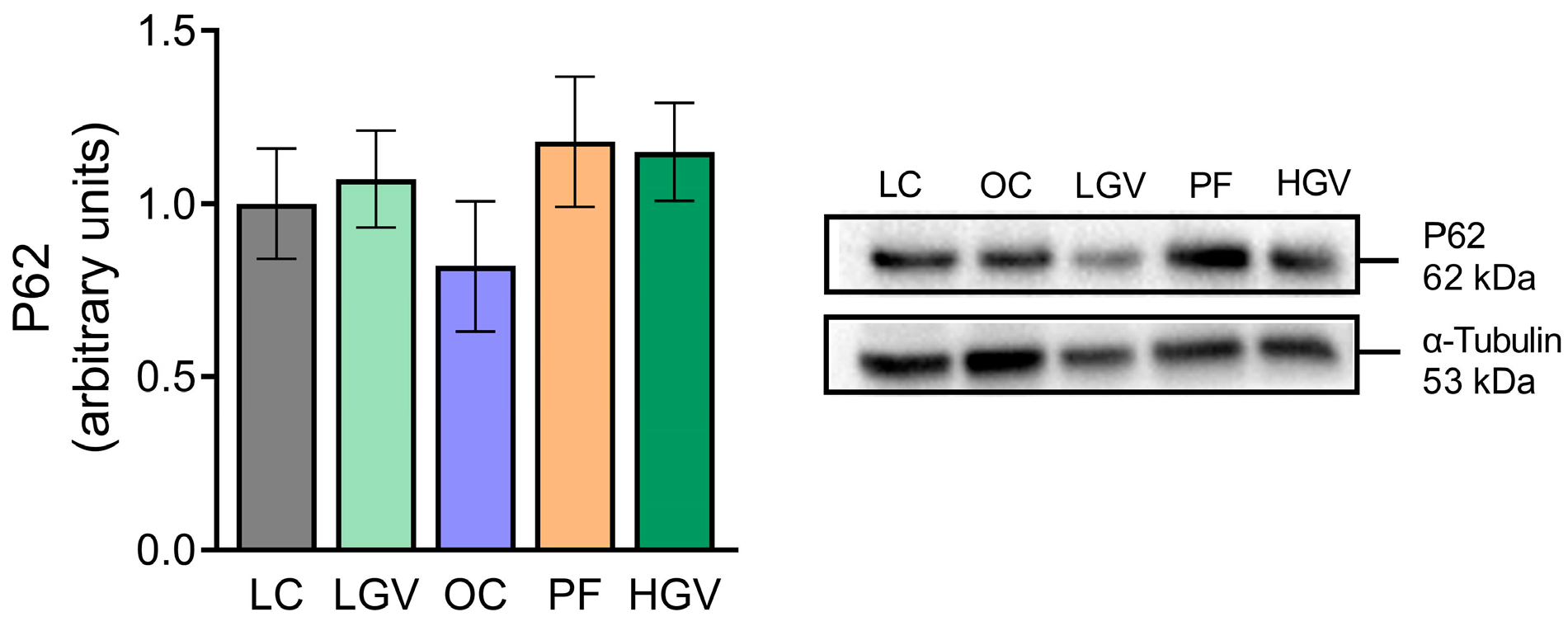
3.8. Parameters Related to Triglyceride Secretion and Autophagy
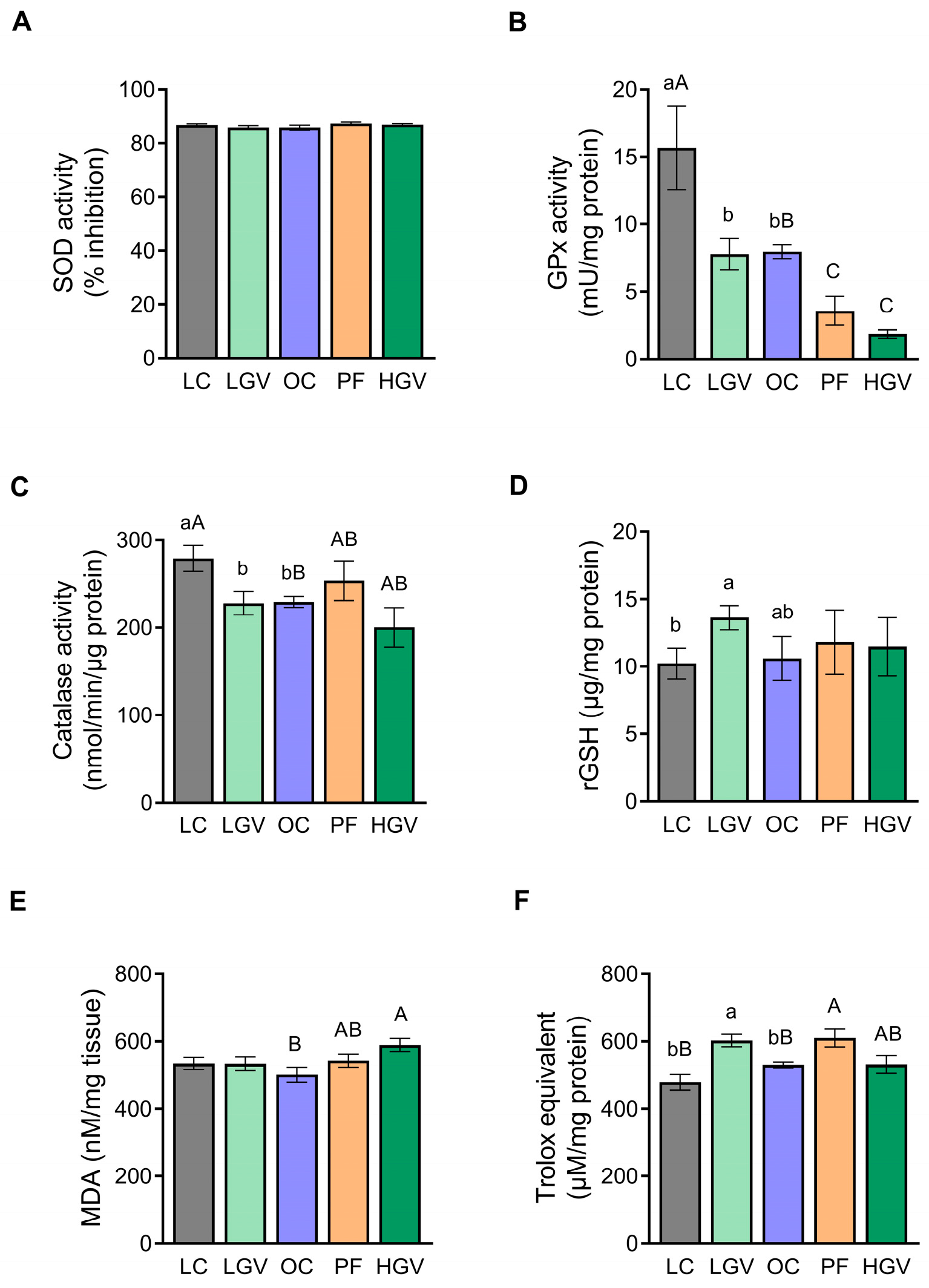
3.9. Hepatic Oxidative Stress Markers
3.10. Gene Expression of Inflammation-Related Markers in Liver
3.11. Gene Expression of Fibrogenic Markers in Liver
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Teng, M.L.; Ng, C.H.; Huang, D.Q.; Chan, K.E.; Tan, D.J.; Lim, W.H.; Yang, J.D.; Tan, E.; Muthiah, M.D. Global incidence and prevalence of nonalcoholic fatty liver disease. Clin. Mol. Hepatol. 2023, 29, S32–S42. [Google Scholar] [CrossRef]
- Ramírez-Mejía, M.M.; Qi, X.; Abenavoli, L.; Romero-Gómez, M.; Eslam, M.; Méndez-Sánchez, N. Metabolic dysfunction: The silenced connection with fatty liver disease. Ann. Hepatol. 2023, 28, 101138. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J.; Panel, I.C. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef] [PubMed]
- Nassir, F.; Rector, R.S.; Hammoud, G.M.; Ibdah, J.A. Pathogenesis and Prevention of Hepatic Steatosis. Gastroenterol. Hepatol. 2015, 11, 167–175. [Google Scholar]
- Neuschwander-Tetri, B.A. Hepatic lipotoxicity and the pathogenesis of nonalcoholic steatohepatitis: The central role of nontriglyceride fatty acid metabolites. Hepatology 2010, 52, 774–788. [Google Scholar] [CrossRef]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Arroyave-Ospina, J.C.; Wu, Z.; Geng, Y.; Moshage, H. Role of Oxidative Stress in the Pathogenesis of Non-Alcoholic Fatty Liver Disease: Implications for Prevention and Therapy. Antioxidants 2021, 10, 174. [Google Scholar] [CrossRef]
- Clare, K.; Dillon, J.F.; Brennan, P.N. Reactive Oxygen Species and Oxidative Stress in the Pathogenesis of MAFLD. J. Clin. Transl. Hepatol. 2022, 10, 939–946. [Google Scholar] [CrossRef]
- Roohinejad, S.; Koubaa, M.; Barba, F.J.; Saljoughian, S.; Amid, M.; Greiner, R. Application of seaweeds to develop new food products with enhanced shelf-life, quality and health-related beneficial properties. Food Res. Int. 2017, 99, 1066–1083. [Google Scholar] [CrossRef]
- Brown, E.S.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.; Nitecki, S.; Strain, C.R.; McSorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Biris-Dorhoi, E.S.; Michiu, D.; Pop, C.R.; Rotar, A.M.; Tofana, M.; Pop, O.L.; Socaci, S.A.; Farcas, A.C. Macroalgae-A Sustainable Source of Chemical Compounds with Biological Activities. Nutrients 2020, 12, 3085. [Google Scholar] [CrossRef]
- Kazir, M.; Abuhassira, Y.; Robin, A.; Nahor, O.; Luo, J.; Israel, A.; Golberg, A.; Livney, Y.D. Extraction of proteins from two marine macroalgae, Ulva sp. and Gracilaria sp., for food application, and evaluating digestibility, amino acid composition and antioxidant properties of the protein concentrates. Food Hydrocoll. 2019, 87, 194–203. [Google Scholar] [CrossRef]
- Thakuri, L.S.; Park, C.M.; Kim, H.A.; Kim, H.J.; Park, J.W.; Park, J.C.; Rhyu, D.Y. Gracilaria chorda subcritical water ameliorates hepatic lipid accumulation and regulates glucose homeostasis in a hepatic steatosis cell model and obese C57BL/6J mice. J. Ethnopharmacol. 2024, 320, 117395. [Google Scholar] [CrossRef]
- Huang, S.; Pang, D.; Li, X.; You, L.; Zhao, Z.; Cheung, P.C.; Zhang, M.; Liu, D. A sulfated polysaccharide from Gracilaria Lemaneiformis regulates cholesterol and bile acid metabolism in high-fat diet mice. Food Funct. 2019, 10, 3224–3236. [Google Scholar] [CrossRef]
- Li, X.; Huang, S.; Chen, X.; Xu, Q.; Ma, Y.; You, L.; Kulikouskaya, V.; Xiao, J.; Piao, J. Structural characteristic of a sulfated polysaccharide from Gracilaria Lemaneiformis and its lipid metabolism regulation effect. Food Funct. 2020, 11, 10876–10885. [Google Scholar] [CrossRef]
- Chan, P.T.; Matanjun, P.; Yasir, S.M.; Tan, T.S. Antioxidant and hypolipidaemic properties of red seaweed, Gracilaria changii. J. Appl. Phycol. 2014, 26, 987–997. [Google Scholar] [CrossRef]
- González-Arceo, M.; Trepiana, J.; Aguirre, L.; Ibarruri, J.; Martínez-Sanz, M.; Cebrián, M.; Recio, I.; Portillo, M.P.; Gómez-Zorita, S. Anti-Steatotic Effects of Chlorella vulgaris, Nannochloropsis gaditana and Gracilaria vermiculophylla Algae Extracts in AML-12 Hepatocytes. Nutrients 2023, 15, 1960. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, C.D.; Wallentinus, I. Long-term survival of an introduced red alga in adverse conditions. Mar. Biol. Res. 2009, 5, 304–308. [Google Scholar] [CrossRef]
- Rueness, J. Life history and molecular sequences of Gracilaria vermiculophylla (Gracilariales, Rhodophyta), a new introduction to European waters. Phycologia 2005, 44, 120–128. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Perseghin, G.; Caumo, A.; Caloni, M.; Testolin, G.; Luzi, L. Incorporation of the fasting plasma FFA concentration into QUICKI improves its association with insulin sensitivity in nonobese individuals. J. Clin. Endocrinol. Metab. 2001, 86, 4776–4781. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Maciá, M.; Simón, J.; González-Rellan, M.J.; Azkargorta, M.; Goikoetxea-Usandizaga, N.; Lopitz-Otsoa, F.; De Urturi, D.S.; Rodríguez-Agudo, R.; Lachiondo-Ortega, S.; Mercado-Gomez, M.; et al. Neddylation inhibition ameliorates steatosis in NAFLD by boosting hepatic fatty acid oxidation via the DEPTOR-mTOR axis. Mol. Metab. 2021, 53, 101275. [Google Scholar] [CrossRef] [PubMed]
- Lynen, F. [3] Yeast fatty acid synthase. Methods Enzymol. 1969, 14, 17–33. [Google Scholar] [CrossRef]
- Bieber, L.L.; Abraham, T.; Helmrath, T. A rapid spectrophotometric assay for carnitine palmitoyltransferase. Anal. Biochem. 1972, 50, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Lazarow, P.B. Assay of peroxisomal beta-oxidation of fatty acids. Methods Enzymol. 1981, 72, 315–319. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Aebi, H. vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Rajapakse, N.; Kim, S.K. Nutritional and digestive health benefits of seaweed. Adv. Food Nutr. Res. 2011, 64, 17–28. [Google Scholar] [CrossRef]
- Eun-Kyung, C.; Soo-Hyun, P.; Ki-Chan, H.; Soon-Ok, N.; Su-Jin, J.; Han-Jung, C.; Soo-Wan, C.; Tae-Sun, P. Clinical trial of the hypolipidemic effects of a brown alga Ecklonia cava extract in patients with hypercholesterolemia. Int. J. Pharmacol. 2015, 11, 798–805. [Google Scholar]
- Paradis, M.E.; Couture, P.; Lamarche, B. A randomised crossover placebo-controlled trial investigating the effect of brown seaweed (Ascophyllum nodosum and Fucus vesiculosus) on postchallenge plasma glucose and insulin levels in men and women. Appl. Physiol. Nutr. Metab. 2011, 36, 913–919. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kwon, Y.M.; Kim, I.S.; Kim, J.A.; Yu, D.Y.; Adhikari, B.; Lee, S.S.; Choi, I.S.; Cho, K.K. Effects of the Brown Seaweed. Front. Nutr. 2018, 5, 23. [Google Scholar] [CrossRef]
- du Preez, R.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. (Sea Grapes) Improves Cardiovascular and Metabolic Health of Rats with Diet-Induced Metabolic Syndrome. Metabolites 2020, 10, 500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Fan, X.Y.; Guo, W.L.; Cao, Y.J.; Lin, Y.C.; Cheng, W.J.; Chen, L.J.; Rao, P.F.; Ni, L.; Lv, X.C. The protective mechanisms of macroalgae Laminaria japonica consumption against lipid metabolism disorders in high-fat diet-induced hyperlipidemic rats. Food Funct. 2020, 11, 3256–3270. [Google Scholar] [CrossRef] [PubMed]
- Wanyonyi, S.; du Preez, R.; Brown, L.; Paul, N.A.; Panchal, S.K. Kappaphycus alvarezii as a Food Supplement Prevents Diet-Induced Metabolic Syndrome in Rats. Nutrients 2017, 9, 1261. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Hirazawa, C.; Mizutani, T.; Yoshikawa, R.; Ohya, T.; Ma, N.; Owaki, Y.; Owaki, T.; Ito, T.; Matsuzaki, C. The anti-obesity and anti-diabetic effects of the edible seaweed. Food Sci. Nutr. 2023, 11, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.B.; Lee, Y.; Bae, M.; Kang, H.; Hu, S.; Pham, T.X.; Lee, J.Y.; Park, Y.K. Sugar kelp (Saccharina latissima) inhibits hepatic inflammation and fibrosis in a mouse model of diet-induced nonalcoholic steatohepatitis. J. Nutr. Biochem. 2021, 97, 108799. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Hirazawa, C.; Yoshikawa, R.; Mizutani, T.; Ohya, T.; Ma, N.; Ikemori, T.; Ito, T.; Matsuzaki, C. Edible red seaweed Campylaephora hypnaeoides J. Agardh alleviates obesity and related metabolic disorders in mice by suppressing oxidative stress and inflammatory response. Nutr. Metab. 2022, 19, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gu, Y.; Wu, X.; Rayamajhi, S.; Bian, S.; Zhang, Q.; Meng, G.; Liu, L.; Wu, H.; Zhang, S.; et al. Association between consumption of edible seaweeds and newly diagnosed non-alcohol fatty liver disease: The TCLSIH Cohort Study. Liver Int. 2021, 41, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Guzzardi, M.A.; Guiducci, L.; Campani, D.; La Rosa, F.; Cacciato Insilla, A.; Bartoli, A.; Cabiati, M.; De Sena, V.; Del Ry, S.; Burchielli, S.; et al. Leptin resistance before and after obesity: Evidence that tissue glucose uptake underlies adipocyte enlargement and liver steatosis/steatohepatitis in Zucker rats from early-life stages. Int. J. Obes. 2022, 46, 50–58. [Google Scholar] [CrossRef]
- Gómez-Zorita, S.; Fernández-Quintela, A.; Macarulla, M.T.; Aguirre, L.; Hijona, E.; Bujanda, L.; Milagro, F.; Martínez, J.A.; Portillo, M.P. Resveratrol attenuates steatosis in obese Zucker rats by decreasing fatty acid availability and reducing oxidative stress. Br. J. Nutr. 2012, 107, 202–210. [Google Scholar] [CrossRef]
- Argilés, J.M. The obese Zucker rat: A choice for fat metabolism 1968–1988: Twenty years of research on the insights of the Zucker mutation. Prog. Lipid Res. 1989, 28, 53–66. [Google Scholar] [CrossRef]
- Aleixandre de Artiñano, A.; Miguel Castro, M. Experimental rat models to study the metabolic syndrome. Br. J. Nutr. 2009, 102, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.T.; Matanjun, P.; Yasir, S.M.; Tan, T.S. Histopathological studies on liver, kidney and heart of normal and dietary induced hyperlipidaemic rats fed with tropical red seaweed Gracilaria changii. J. Funct. Foods 2015, 17, 202–213. [Google Scholar] [CrossRef]
- Norziah, M.H.; Ching, C.Y. Nutritional composition of edible seaweed Gracilaria changgi. Food Chem. 2000, 68, 69–76. [Google Scholar] [CrossRef]
- Sengupta, P. The Laboratory Rat: Relating Its Age With Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Zabala, A.; Churruca, I.; Macarulla, M.T.; Rodriguez, V.M.; Fernandez-Quintela, A.; Martinez, J.A.; Portillo, M.P. The trans-10,cis-12 isomer of conjugated linoleic acid reduces hepatic triacylglycerol content without affecting lipogenic enzymes in hamsters. Br. J. Nutr. 2004, 92, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, L.; Zhu, L.; Yu, Y. Free fatty acids, not triglycerides, are associated with non-alcoholic liver injury progression in high fat diet induced obese rats. Lipids Health Dis. 2016, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Urstad, A.P.; Semenkovich, C.F. Fatty acid synthase and liver triglyceride metabolism: Housekeeper or messenger? Biochim. Biophys. Acta 2012, 1821, 747–753. [Google Scholar] [CrossRef]
- Morán-Ramos, S.; Avila-Nava, A.; Tovar, A.R.; Pedraza-Chaverri, J.; López-Romero, P.; Torres, N. Opuntia ficus indica (nopal) attenuates hepatic steatosis and oxidative stress in obese Zucker (fa/fa) rats. J. Nutr. 2012, 142, 1956–1963. [Google Scholar] [CrossRef]
- Jacobsen, C.; Sørensen, A.M.; Holdt, S.L.; Akoh, C.C.; Hermund, D.B. Source, Extraction, Characterization, and Applications of Novel Antioxidants from Seaweed. Annu. Rev. Food Sci. Technol. 2019, 10, 541–568. [Google Scholar] [CrossRef]
- El-Lakkany, N.M.; Seif El-Din, S.H.; Sabra, A.A.; Hammam, O.A.; Ebeid, F.A. Co-administration of metformin and N-acetylcysteine with dietary control improves the biochemical and histological manifestations in rats with non-alcoholic fatty liver. Res. Pharm. Sci. 2016, 11, 374–382. [Google Scholar] [CrossRef]
- Begriche, K.; Massart, J.; Robin, M.A.; Bonnet, F.; Fromenty, B. Mitochondrial adaptations and dysfunctions in nonalcoholic fatty liver disease. Hepatology 2013, 58, 1497–1507. [Google Scholar] [CrossRef]
- Chan, P.T.; Matanjun, P.; Yasir, S.M.; Tan, T.S. Oxidative stress biomarkers in organs of hyperlipidaemic and normal rats fed tropical red seaweed, Gracilaria changii. J. Appl. Phycol. 2016, 28, 1371–1378. [Google Scholar] [CrossRef]
- Jin, M.; Liu, H.; Hou, Y.; Chan, Z.; Di, W.; Li, L.; Zeng, R. Preparation, characterization and alcoholic liver injury protective effects of algal oligosaccharides from Gracilaria lemaneiformis. Food Res. Int. 2017, 100, 186–195. [Google Scholar] [CrossRef]
- Barros-Gomes, J.A.C.; Nascimento, D.L.A.; Silveira, A.C.R.; Silva, R.K.; Gomes, D.L.; Melo, K.R.T.; Almeida-Lima, J.; Camara, R.B.G.; Silva, N.B.; Rocha, H.A.O. Evaluation of the Antioxidant Activity and Protective Action of the Seaweed. Oxidative Med. Cell. Longev. 2018, 2018, 9354296. [Google Scholar] [CrossRef]
- Carmiel-Haggai, M.; Cederbaum, A.I.; Nieto, N. A high-fat diet leads to the progression of non-alcoholic fatty liver disease in obese rats. FASEB J. 2005, 19, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Wang, F.; Zhai, D.; Meng, X.; Liu, J.; Lv, X. Matrix metalloproteinases induce extracellular matrix degradation through various pathways to alleviate hepatic fibrosis. Biomed. Pharmacother. 2023, 161, 114472. [Google Scholar] [CrossRef] [PubMed]
- Kasarinaite, A.; Sinton, M.; Saunders, P.T.K.; Hay, D.C. The Influence of Sex Hormones in Liver Function and Disease. Cells 2023, 12, 1604. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Sense Primer 5′-3′ | Antisense Primer 5′-3′ |
|---|---|---|---|
| Acta2 | NM_031004.2 | GCC GAG ATC TCA CCG ACT AC | GTC CAG AGC GAC ATA GCA CA |
| Actb | NM_031144.3 | CCC GCG AGT ACA ACC TTC T | CGT CAT CCA TGG CGA ACT |
| CD206 | NM_001106123.2 | ACT GCG TGG TGA TGA AAG G | TAA CCC AGT GGT TGC TCA CA |
| Col1a1 | NM_053304.1 | TCC TGG CAA GAA CGG AGA T | CAG GAG GTC CAC GCT CAC |
| Crp | NM_017096.4 | TGT CTC TAT GCC CAC GCT GAT G | GGC CCA CCT ACT GCA ATA CTA AAC |
| F4/80 | NM_001007557.2 | CTC TTC CTG ATG GTG AGA AAC C | CCC ATG GAT GTA CAG TAG CAG A |
| Il1b | NM_031512.2 | TGT GAT GAA AGA CGG CAC AC | CTT CTT CTT TGG GTA TTG TTT GG |
| Mmp9 | NM_031055.2 | AGC CGA CGT CAC TGT AAC TG | CCA GGA AGA CGA AGG GGA AG |
| Timp1 | NM_053819.1 | CAG CAA AAG GCC TTC GTA AA | TGG CTG AAC AGG GAA ACA CT |
| Tgfb1 | NM_021578.2 | CCT GGA AAG GGC TCA ACA C | TGC CGT ACA CAG CAG TTC TT |
| Gracilaria vermiculophylla | |
|---|---|
| Energy (kcal/100 g) | 182 |
| Total lipids (g/100 g) | 1.40 |
| Saturated fatty acids (g/100 g) | 0.98 |
| Total carbohydrates (g/100 g) | 6.40 |
| Simple carbohydrates (g/100 g) | <0.50 |
| Proteins (g/100 g) | 22.10 |
| Fibre (g/100 g) | 27.90 |
| Ashes (g/100 g) | 28.8 |
| Moisture (g/100 g) | 13.4 |
| LC | LGV | OC | PF | HGV | |
|---|---|---|---|---|---|
| Glucose (mg/dL) | 75 ± 7 bB | 76 ± 12 ab | 100 ± 8 aA | 74 ± 4 B | 78 ± 9 AB |
| Insulin (ng/mL) | 0.8 ± 0.1 bC | 25.3 ± 4.1 a | 26.6 ± 1.5 aA | 17.5 ± 3.2 B | 15.8 ± 2.4 B |
| HOMA-IR | 3.5 ± 0.7 bC | 119.5 ± 28.2 a | 174.2 ± 13.3 aA | 70.8 ± 13.6 B | 77.1 ± 15.4 B |
| TG (mg/dL) | 26 ± 3 bB | 151 ± 27 a | 206 ± 50 aA | 199 ± 33 A | 219 ± 50 A |
| NEFA (mg/dL) | 8.7 ± 0.9 bB | 35.4 ± 4.4 a | 29.3 ± 2.0 aA | 32.0 ± 3.6 A | 37.0 ± 6.5 A |
| R-QUICKI | 0.38 ± 0.02 aB | 0.29 ± 0.01 b | 0.28 ± 0.01 bC | 0.31 ± 0.01 A | 0.31 ± 0.01 A |
| ALT/GPT (U/L) | 56 ± 2 bB | 119 ± 11 a | 110 ± 12 aA | 98 ± 7 A | 119 ± 13 A |
| AST/GOT (U/L) | 142 ± 8 bB | 190 ± 9 a | 144 ± 11 bB | 166 ± 11 AB | 199 ± 16 A |
| AST/ALT | 2.6 ± 0.1 aA | 1.8 ± 0.1 b | 1.4 ± 0.1 cC | 1.7 ± 0.1 B | 1.6 ± 0.1 B |
| ALP (U/L) | 278 ± 6 cC | 515 ± 24 a | 388 ± 15 bB | 400 ± 15.1 B | 542 ± 36 A |
| Uric acid (mg/mL) | 1.6 ± 0.1 bB | 5.0 ± 0.8 a | 4.3 ± 0.4 aA | 3.6 ± 0.5 A | 6.4 ± 1.3 A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Arceo, M.; Aguirre, L.; Macarulla, M.T.; Gil-Pitarch, C.; Martínez-Chantar, M.L.; Portillo, M.P.; Gómez-Zorita, S. Effect of Gracilaria vermiculophylla Macroalga on Non-Alcoholic Fatty Liver Disease in Obese Rats. Antioxidants 2024, 13, 369. https://doi.org/10.3390/antiox13030369
González-Arceo M, Aguirre L, Macarulla MT, Gil-Pitarch C, Martínez-Chantar ML, Portillo MP, Gómez-Zorita S. Effect of Gracilaria vermiculophylla Macroalga on Non-Alcoholic Fatty Liver Disease in Obese Rats. Antioxidants. 2024; 13(3):369. https://doi.org/10.3390/antiox13030369
Chicago/Turabian StyleGonzález-Arceo, Maitane, Leixuri Aguirre, María Teresa Macarulla, Clàudia Gil-Pitarch, María Luz Martínez-Chantar, María P. Portillo, and Saioa Gómez-Zorita. 2024. "Effect of Gracilaria vermiculophylla Macroalga on Non-Alcoholic Fatty Liver Disease in Obese Rats" Antioxidants 13, no. 3: 369. https://doi.org/10.3390/antiox13030369
APA StyleGonzález-Arceo, M., Aguirre, L., Macarulla, M. T., Gil-Pitarch, C., Martínez-Chantar, M. L., Portillo, M. P., & Gómez-Zorita, S. (2024). Effect of Gracilaria vermiculophylla Macroalga on Non-Alcoholic Fatty Liver Disease in Obese Rats. Antioxidants, 13(3), 369. https://doi.org/10.3390/antiox13030369