

Antioxidant Actions of Melatonin: A Systematic Review of Animal Studies

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Esteban-Zubero, E.; García-Gil, F.A.; López-Pingarrón, L.; Alatorre-Jiménez, M.A.; Iñigo-Gil, P.; Tan, D.-X.; García, J.J.; Reiter, R.J. Potential benefits of melatonin in organ transplantation: A review. J. Endocrinol. 2016, 229, R129–R146. [Google Scholar] [CrossRef] [PubMed]
- Yong, W.; Ma, H.; Na, M.; Gao, T.; Zhang, Y.; Hao, L.; Yu, H.; Yang, H.; Deng, X. Roles of melatonin in the field of reproductive medicine. Biomed. Pharmacother. 2021, 144, 112001. [Google Scholar] [CrossRef] [PubMed]
- Cipolla-Neto, J.; Amaral, F.G.D. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Jozaki, M.; Tanabe, M.; Shirafuta, Y.; Mihara, Y.; Shinagawa, M.; Tamura, I.; Maekawa, R.; Sato, S.; Taketani, T.; et al. Importance of Melatonin in Assisted Reproductive Technology and Ovarian Aging. Int. J. Mol. Sci. 2020, 21, 1135. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Chen, L.D.; Poeggeler, B.; Manchester, L.C.; Reiter, R.J. Melatonin: A potent, endogenous hydroxyl radical scavenger. Endocr. J. 1993, 1, 57–60. [Google Scholar]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Ramis, M.R.; Esteban, S.; Miralles, A.; Tan, D.X.; Reiter, R.J. Protective effects of melatonin and mitochondria-targeted antioxidants against oxidative stress: A review. Curr. Med. Chem. 2015, 22, 2690–2711. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Brömme, H.; Mörke, W.; Peschke, E.; Ebelt, H.; Peschke, D. Scavenging effect of melatonin on hydroxyl radicals generated by alloxan. J. Pineal Res. 2000, 29, 201–208. [Google Scholar] [CrossRef]
- Galano, A. On the direct scavenging activity of melatonin towards hydroxyl and a series of peroxyl radicals. Phys. Chem. Chem. Phys. 2011, 13, 7178–7188. [Google Scholar] [CrossRef]
- Hardeland, R.; Reiter, R.J.; Poeggeler, B.; Tan, D.X. The significance of the metabolism of the neurohormone melatonin: Antioxidative protection and formation of bioactive substances. Neurosci. Biobehav. Rev. 1993, 17, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Razek, H.A.; Rizk, M.S.; Amer, G.S.; Kora, M.A.; Afifi, A.M.; Donia, S.S. Impact of combined ischemic preconditioning and melatonin on renal ischemia-reperfusion injury in rats. Iran. J. Basic Med. Sci. 2020, 26, 235–240. [Google Scholar] [CrossRef]
- Colares, J.R.; Hartmann, R.M.; Schemitt, E.G.; Fonseca, S.R.B.; Brasil, M.S.; Picada, J.N.; Dias, A.S.; Bueno, A.F.; Marroni, C.A.; Marroni, N.P. Melatonin prevents oxidative stress, inflammatory activity, and DNA damage in cirrhotic rats. World J. Gastroenterol. 2020, 28, 348–364. [Google Scholar] [CrossRef] [PubMed]
- Essawy, A.E.; Mohamed, A.I.; Ali, R.G.; Ali, A.M.; Abdou, H.M. Analysis of Melatonin-Modulating Effects Against Tartrazine-Induced Neurotoxicity in Male Rats: Biochemical, Pathological and Immunohistochemical Markers. Neurochem. Res. 2023, 48, 131–141. [Google Scholar] [CrossRef]
- Aranarochana, A.; Sirichoat, A.; Pannangrong, W.; Wigmore, P.; Welbat, J.U. Melatonin Ameliorates Valproic Acid-Induced Neurogenesis Impairment: The Role of Oxidative Stress in Adult Rats. Oxidative Med. Cell. Longev. 2021, 2021, 9997582. [Google Scholar] [CrossRef] [PubMed]
- Bashandy, S.A.E.; Ebaid, H.; Al-Tamimi, J.; Ahmed-Farid, O.A.-H.; Omara, E.A.; Alhazza, I.M. Melatonin Alleviated Potassium Dichromate-Induced Oxidative Stress and Reprotoxicity in Male Rats. BioMed Res. Int. 2021, 2021, 3565360. [Google Scholar] [CrossRef] [PubMed]
- Olayaki, L.A.; Alagbonsi, I.A.; Abdulrahim, A.H.; Adeyemi, W.J.; Bakare, M.; Omeiza, N. Melatonin prevents and ameliorates lead-induced gonadotoxicity through antioxidative and hormonal mechanisms. Toxicol. Ind. Health 2018, 34, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Nayki, U.; Onk, D.; Balci, G.; Nayki, C.; Onk, A.; Çankaya, M.; Kafa, A.H.T.; Kuzucu, M. The effect of melatonin on oxidative stress and apoptosis in experimental diabetes mellitus-related ovarian injury. Gynecol. Endocrinol. 2016, 32, 421–426. [Google Scholar] [CrossRef]
- Taghizadieh, M.; Hajipour, B.; Asl, N.A.; Khodadadi, A.; Somi, M.H.; Banei, M. Combination effect of melatonin and dexamethasone on liver ischemia/reperfusion injury. Bratisl. Med. J. 2016, 116, 47–53. [Google Scholar] [CrossRef]
- Cruz, A.; Tasset, I.; Ramírez, L.M.; Arjona, A.; Segura, J.; Túnez, I.; Montilla, P.; Muntané, J.; Padillo, F.J. Effect of melatonin on myocardial oxidative stress induced by experimental obstructive jaundice. Rev. Esp. Enferm. Dig. 2009, 101, 460–463. [Google Scholar] [CrossRef]
- Baydas, G.; Canatan, H.; Turkoglu, A. Comparative analysis of the protective effects of melatonin and vitamin E on streptozocin-induced diabetes mellitus. J. Pineal Res. 2002, 32, 225–230. [Google Scholar] [CrossRef]
- Ferreira, C.d.S.; Maganhin, C.C.; Simões, R.d.S.; Girão, M.J.B.C.; Baracat, E.C.; Soares-Jr, J.M. Melatonin: Cell death Modulator. Rev. Assoc. Med. Bras. 2010, 56, 715–718. [Google Scholar] [CrossRef]
- Ferreira, C.S.; Carvalho, K.C.; Maganhin, C.C.; Paiotti, A.P.R.; Oshima, C.T.F.; Simões, M.J.; Baracat, E.C.; Soares, J.M. Does melatonin influence the apoptosis in rat uterus of animals exposed to continuous light? Apoptosis 2016, 21, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.-M.; Pae, H.-O.; Jang, S.-I.; Kim, Y.-M.; Chung, H.-T. Nitric oxide as a pro-apoptotic as well as anti-apoptotic modulator. J. Biochem. Mol. Biol. 2002, 35, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.A.; Blanco, F.J. Cell death and apoptosis in osteoarthritic cartilage. Curr. Drug Targets 2007, 8, 333–345. [Google Scholar] [CrossRef] [PubMed]
- TaTan, D.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Duan, Q.; Wang, Z.; Lu, T.; Chen, J.; Wang, X. Comparison of 6-hydroxylmelatonin or melatonin in protecting neurons against ischemia/reperfusion-mediated injury. J. Pineal Res. 2006, 41, 351–357. [Google Scholar] [CrossRef]
- Sakano, K.; Oikawa, S.; Hiraku, Y.; Kawanishi, S. Oxidative DNA damage induced by a melatonin metabolite, 6-hydroxymelatonin, via a unique non-o-quinone type of redox cycle. Biochem. Pharmacol. 2004, 68, 1869–1878. [Google Scholar] [CrossRef]
- Tan, D.-X.; Manchester, L.C.; Burkhardt, S.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Shohami, E.; Huo, Y.-S.; Hardeland, R.; Reiter, R.J. N1-acetyl-N2-formyl-5-methoxykynuramine, a biogenic amine and melatonin metabolite, functions as a potent antioxidant. FASEB J. 2001, 15, 2294–2296. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. A declaração PRISMA 2020: Uma diretriz atualizada para relatar revisões sistemáticas. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: Exceeding Expectations. Physiology 2014, 29, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Macchi, M.M.; Bruce, J.N. Human pineal physiology and functional significance of melatonin. Front. Neuroendocr. 2004, 25, 177–195. [Google Scholar] [CrossRef] [PubMed]
- Shiroma, M.E.; Botelho, N.M.; Damous, L.L.; Baracat, E.C.; Soares-Jr, J.M. Melatonin influence in ovary transplantation: Systematic review. J. Ovarian Res. 2016, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Hoijman, E.; Viegas, L.R.; Sarmiento, M.I.K.; Rosenstein, R.E.; Pecci, A. Involvement of Bax Protein in the Prevention of Glucocorticoid-Induced Thymocytes Apoptosis by Melatonin. Endocrinology 2004, 145, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Pedreañez, A.; Rincón, J.; Romero, M.; Viera, N.; Mosquera, J. Melatonin decreases apoptosis and expression of apoptosis-associated proteins in acute puromycin aminonucleoside nephrosis. Nephrol. Dial. Transpl. 2004, 19, 1098–1105. [Google Scholar] [CrossRef] [PubMed]
- Molpeceres, V.; Mauriz, J.L.; García-Mediavilla, M.V.; González, P.; Barrio, J.P.; González-Gallego, J. Melatonin Is Able to Reduce the Apoptotic Liver Changes Induced by Aging Via Inhibition of the Intrinsic Pathway of Apoptosis. J. Gerontol. Bio. Sci. 2007, 62, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and Reproduction Revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef]
- Sainz, R.M.; Mayo, J.C.; Rodriguez, C.; Tan, D.X.; Lopez-Burillo, S.; Reiter, R.J. Melatonin and cell death: Differential actions on apoptosis in normal and cancer cells. Cell Mol. Life Sci. 2003, 60, 1407–1426. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Amaral, F.G.; Soares, J.M., Jr.; Gallo, C.C.; Furtado, A.; Cavaco, J.E.; Gonçalves, I.; Santos, C.R.A.; Quintela, T. The Crosstalk between Melatonin and Sex Steroid Hormones. Neuroendocrinology. 2022, 112, 115–129. [Google Scholar] [CrossRef]
- Sapmaz, E.; Ayar, A.; Celik, H.; Sapmaz, T.; Kilic, N.; Yasar, M.A. Effects of melatonin and oxytetracycline in autologous intraperitoneal ovary transplantation in rats. Neuro Endocrinol. Lett. 2003, 24, 350–354. [Google Scholar]
- Shiroma, M.E.; Damous, L.L.; Cotrim, F.P.; Roa, C.L.; Cipolla-Neto, J.; Reiter, R.J.; Baracat, E.C.; Soares, J.M. Pretreatment with melatonin improves ovarian tissue cryopreservation for transplantation. Reprod. Biol. Endocrinol. 2021, 19, 17. [Google Scholar] [CrossRef]
- Ghorbaninejad, P.; Sheikhhossein, F.; Djafari, F.; Tijani, A.J.; Mohammadpour, S.; Shab-Bidar, S. Effects of melatonin supplementation on oxidative stress: A systematic review and meta-analysis of randomized controlled trials. Horm. Mol. Biol. Clin. Investig. 2020, 41, 20200030. [Google Scholar] [CrossRef]




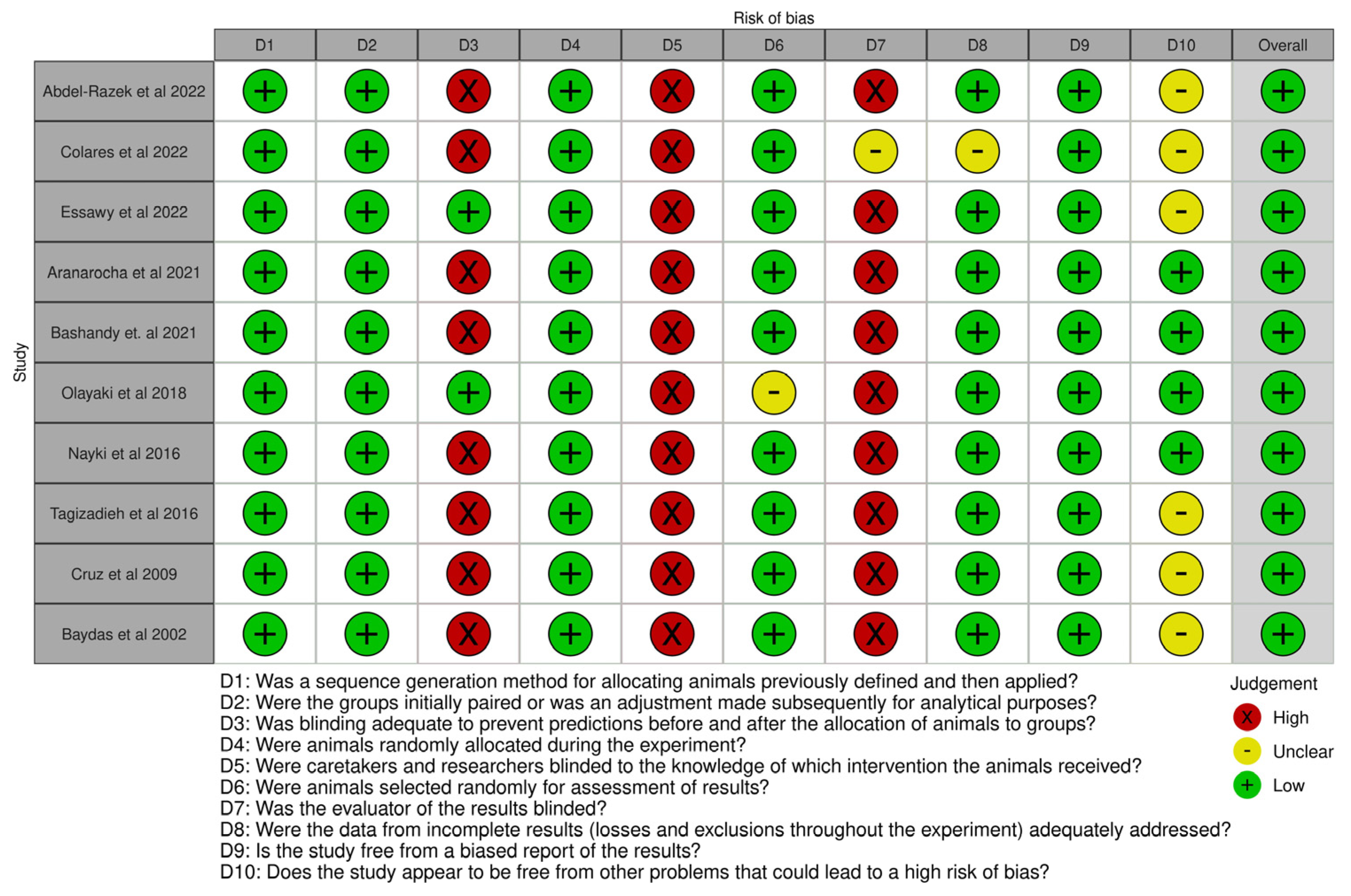{kind=link}
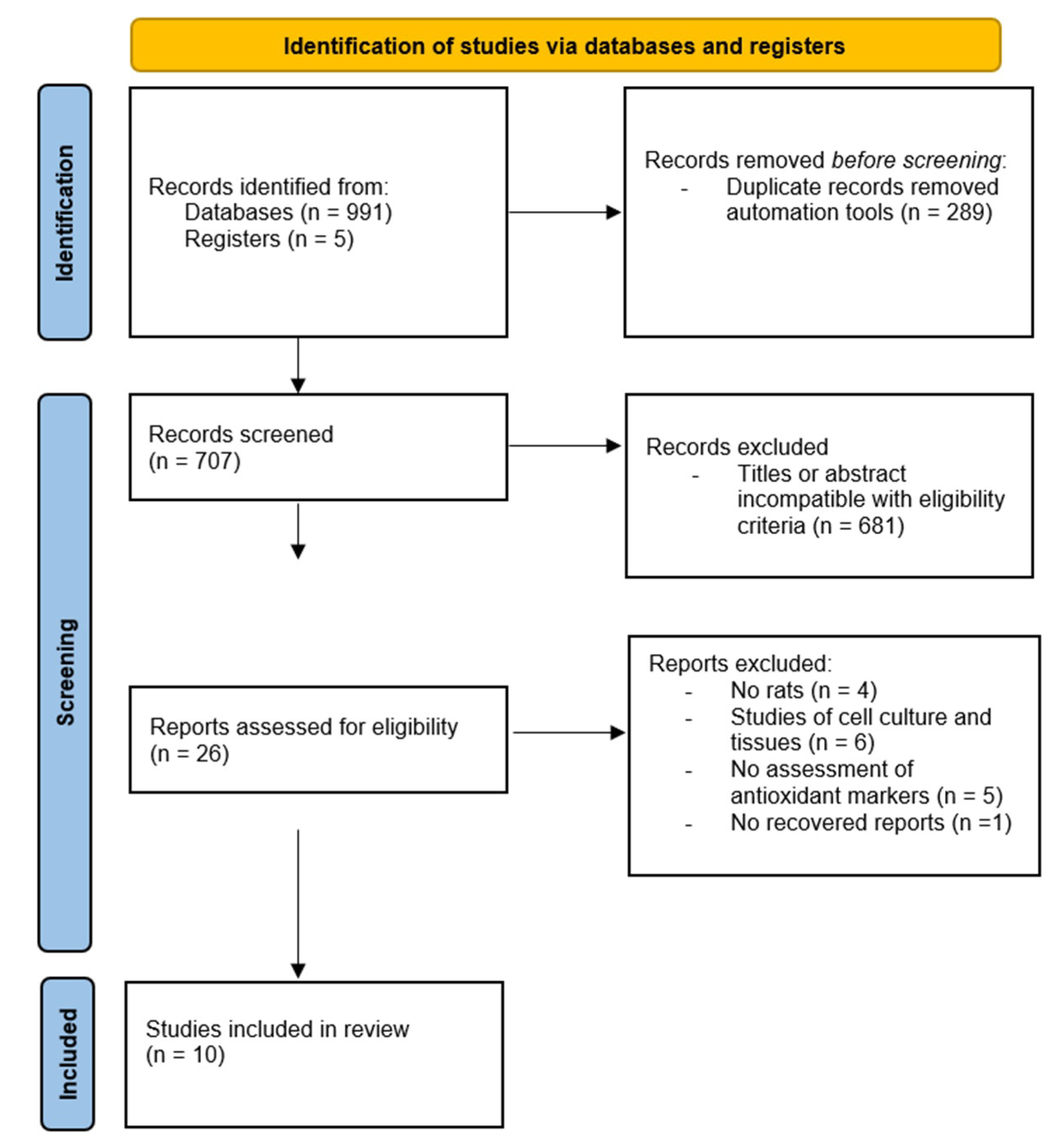{kind=link}
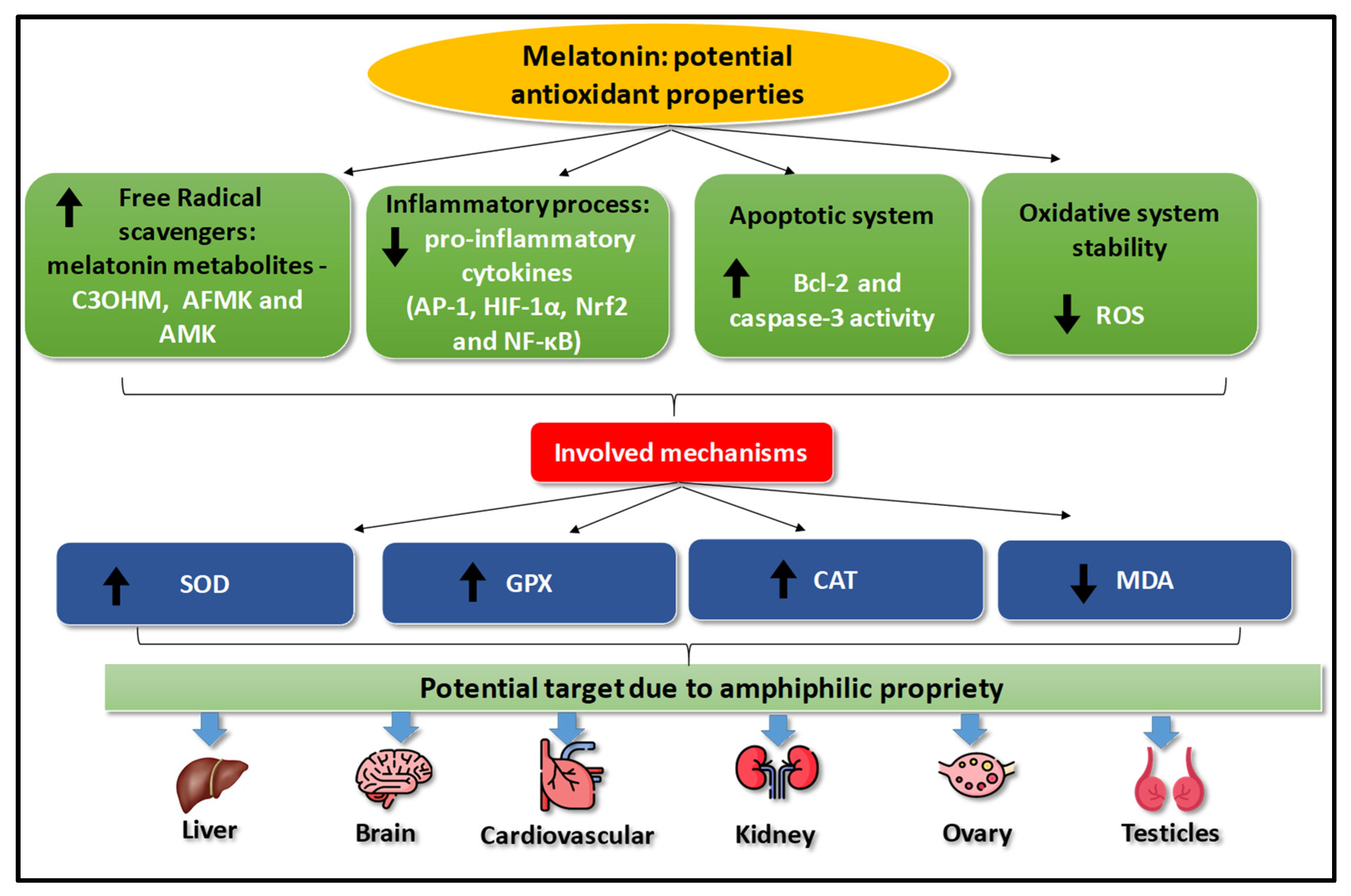{kind=link}
| MeSH Terms |
| Oxidative Stresses Stress OR Oxidative Antioxidative OR Stress Antioxidative Stresses Stress OR Antioxidative OR Antioxidative Stress OR Antioxidative Stress OR Antioxidative Stresses Stress OR Antioxidative Oxidative Damage OR Oxidative Damages OR Oxidative Stress Injury OR Oxidative Stress Injuries OR Stress Injury OR Oxidative Injury OR Oxidative Cleavage OR Oxidative DNA Damage OR DNA Oxidative and Nitrosative Stress OR Nitrative Stress AND (rats) AND (glutathione) AND (GPx) AND (malondialdehyde) AND (SOD) AND (superoxide dismutase) AND (catalase) AND (CAT) |
| Oxidative Stresses Stress OR Oxidative Antioxidative OR Stress Antioxidative Stresses Stress OR Antioxidative OR Antioxidative Stress OR Antioxidative Stress OR Antioxidative Stresses Stress OR Antioxidative Oxidative Damage OR Oxidative Damages OR Oxidative Stress Injury OR Oxidative Stress Injuries OR Stress Injury OR Oxidative Injury OR Oxidative Cleavage OR Oxidative DNA Damage OR DNA Oxidative and Nitrosative Stress Oxidative Nitrative Stress OR Nitrative Stress OR Oxidative Nitrative Stresses Stress OR Nitro-Oxidative Stress OR Nitro Oxidative Stress OR Nitro-Oxidative Stresses Stress, Nitro-Oxidative Stresses OR Nitro-Oxidative) AND (melatonin OR Indoles OR Tryptamines) AND (rats) AND (mice) |
| Oxidative Stresses Stress OR Oxidative Antioxidative OR Stress Antioxidative Stresses Stress OR Antioxidative OR Antioxidative Stress OR Antioxidative Stress OR Antioxidative Stresses Stress OR Antioxidative Oxidative Damage OR Oxidative Damages OR Oxidative Stress Injury OR Oxidative Stress Injuries OR Stress Injury OR Oxidative Injury OR Oxidative Cleavage OR Oxidative DNA Damage OR DNA Oxidative and Nitrosative Stress Oxidative Nitrative Stress OR Nitrative Stress OR Oxidative Nitrative Stresses Stress OR Nitro-Oxidative Stress OR Nitro Oxidative Stress OR Nitro-Oxidative Stresses Stress, Nitro-Oxidative Stresses OR Nitro-Oxidative) AND (melatonin OR Indoles OR Tryptamines) AND (rats) AND (glutathione) |
| (Oxidative Stress OR Oxidative Antioxidative) AND (melatonin OR Indoles OR Tryptamines) AND (rats) AND (glutathione) AND (GPx) AND (malondialdehyde) AND (SOD) AND (superoxide dismutase) AND (catalase) AND (CAT) |
| (Melatonin OR Indoles OR Tryptamines) AND (glutathione) AND (glutathione peroxidase) AND (malondialdehyde) AND (SOD) AND (superoxide dismutase) AND (catalase) AND (CAT) |
| Authors | No. of Rats | Melatonin Dose | Tests | Results |
|---|---|---|---|---|
| Abdel-Razeket al. 2022 [12] | 48 | 10 mg/kg/d Intraperitoneal route | MDA SOD GPx | decreased ++ increased ++ increased + |
| Colares et al. 2022 [13] | 24 | 20 mg/kg/d | SOD | increased ++ |
| Essawy et al. 2022 [14] | 40 | 10 mg/kg/d Oral route | MDA SOD GPx CAT | decreased ++ increased ++ increased ++ increased ++ |
| Aranarocha et al. 2021 [15] | 48 | 8 mg/kg/d Intraperitoneal route | SOD GPx CAT MDA | increased ++ increased ++ increased ++ no difference |
| Bashandy et al. 2021 [16] | 40 | 2.5 or 5 mg/kg/d Intraperitoneal route | MDA SOD CAT GSH | decreased ++ increased ++ increased ++ increased ++ |
| Olayaki et al. 2018 [17] | 50 | 4 or 10 mg/kg/d Oral route | MDA SOD GPx GSH | decreased ++ increased ++ increased ++ increased ++ |
| Nayki et al. 2016 [18] | 24 | 20 mg/kg/d Intraperitoneal route | MDA SOD CAT TOS TAS | decreased ++ increased ++ increased + decreased ++ increased ++ |
| Tagizadieh et al. 2016 [19] | 60 | 20 mg/kg/d Intraperitoneal route | GPx SOD MDA | increased ++ increased ++ decreased ++ |
| Cruz et al. 2009 [20] | 21 | 0.5 mg /kg/d Intraperitoneal route | MDA GSH CAT SOD GPx | decreased ++ increased ++ increased ++ increased ++ increased ++ |
| Baydas et al. 2002 [21] | 80 | 0.1 mg/kg/d Intraperitoneal route | GPx | increased ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, K.K.A.C.; Shiroma, M.E.; Damous, L.L.; Simões, M.d.J.; Simões, R.d.S.; Cipolla-Neto, J.; Baracat, E.C.; Soares-Jr., J.M. Antioxidant Actions of Melatonin: A Systematic Review of Animal Studies. Antioxidants 2024, 13, 439. https://doi.org/10.3390/antiox13040439
Monteiro KKAC, Shiroma ME, Damous LL, Simões MdJ, Simões RdS, Cipolla-Neto J, Baracat EC, Soares-Jr. JM. Antioxidant Actions of Melatonin: A Systematic Review of Animal Studies. Antioxidants. 2024; 13(4):439. https://doi.org/10.3390/antiox13040439
Chicago/Turabian StyleMonteiro, Karla Krislane Alves Costa, Marcos Eiji Shiroma, Luciana Lamarão Damous, Manuel de Jesus Simões, Ricardo dos Santos Simões, José Cipolla-Neto, Edmund Chada Baracat, and Jose Maria Soares-Jr. 2024. "Antioxidant Actions of Melatonin: A Systematic Review of Animal Studies" Antioxidants 13, no. 4: 439. https://doi.org/10.3390/antiox13040439
APA StyleMonteiro, K. K. A. C., Shiroma, M. E., Damous, L. L., Simões, M. d. J., Simões, R. d. S., Cipolla-Neto, J., Baracat, E. C., & Soares-Jr., J. M. (2024). Antioxidant Actions of Melatonin: A Systematic Review of Animal Studies. Antioxidants, 13(4), 439. https://doi.org/10.3390/antiox13040439