
Glutathione S-Transferase Genes Involved in Response to Short-Term Heat Stress in Tetranychus urticae (Koch)



Abstract
1. Introduction
2. Materials and Methods
2.1. Mite Colony
2.2. Selection of GST Genes
2.3. Cloning the CDSs of GST Genes
2.4. Bioinformatic Analysis and Identification of GST Genes
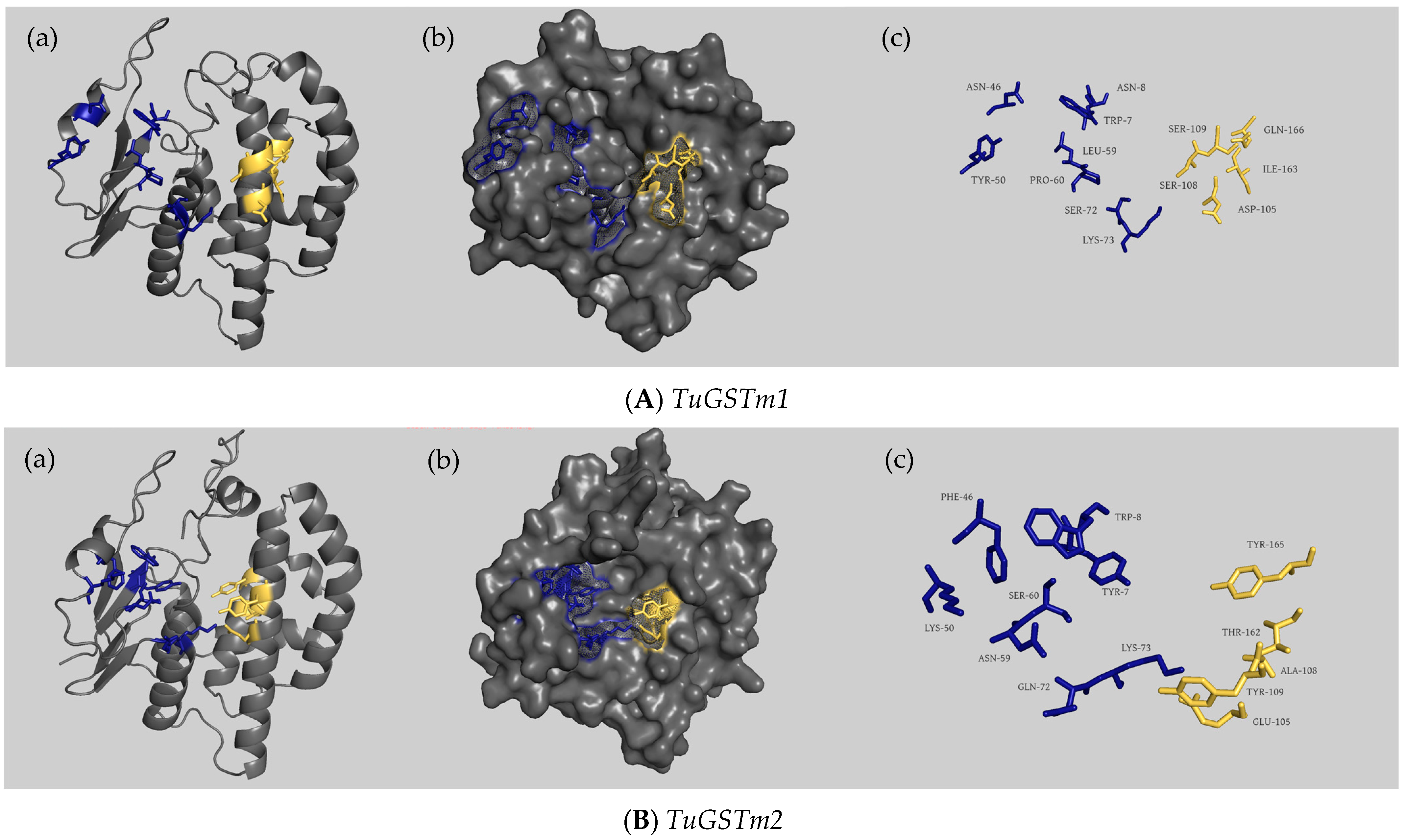
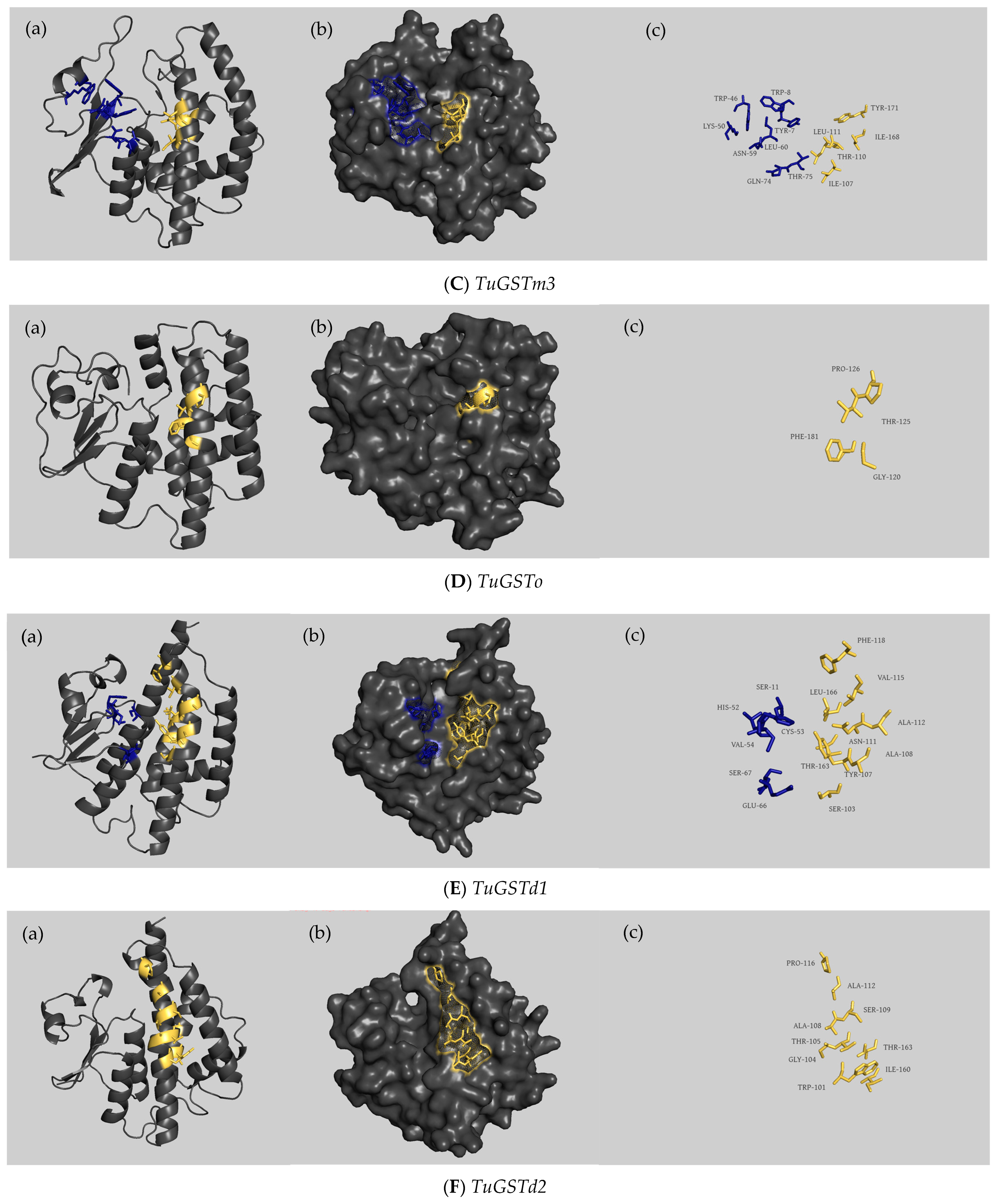
2.5. Structural Characterization of the Coding Region of 6 GST Genes
2.6. Expression of GST Transcripts
2.7. Statistical Analysis
3. Results
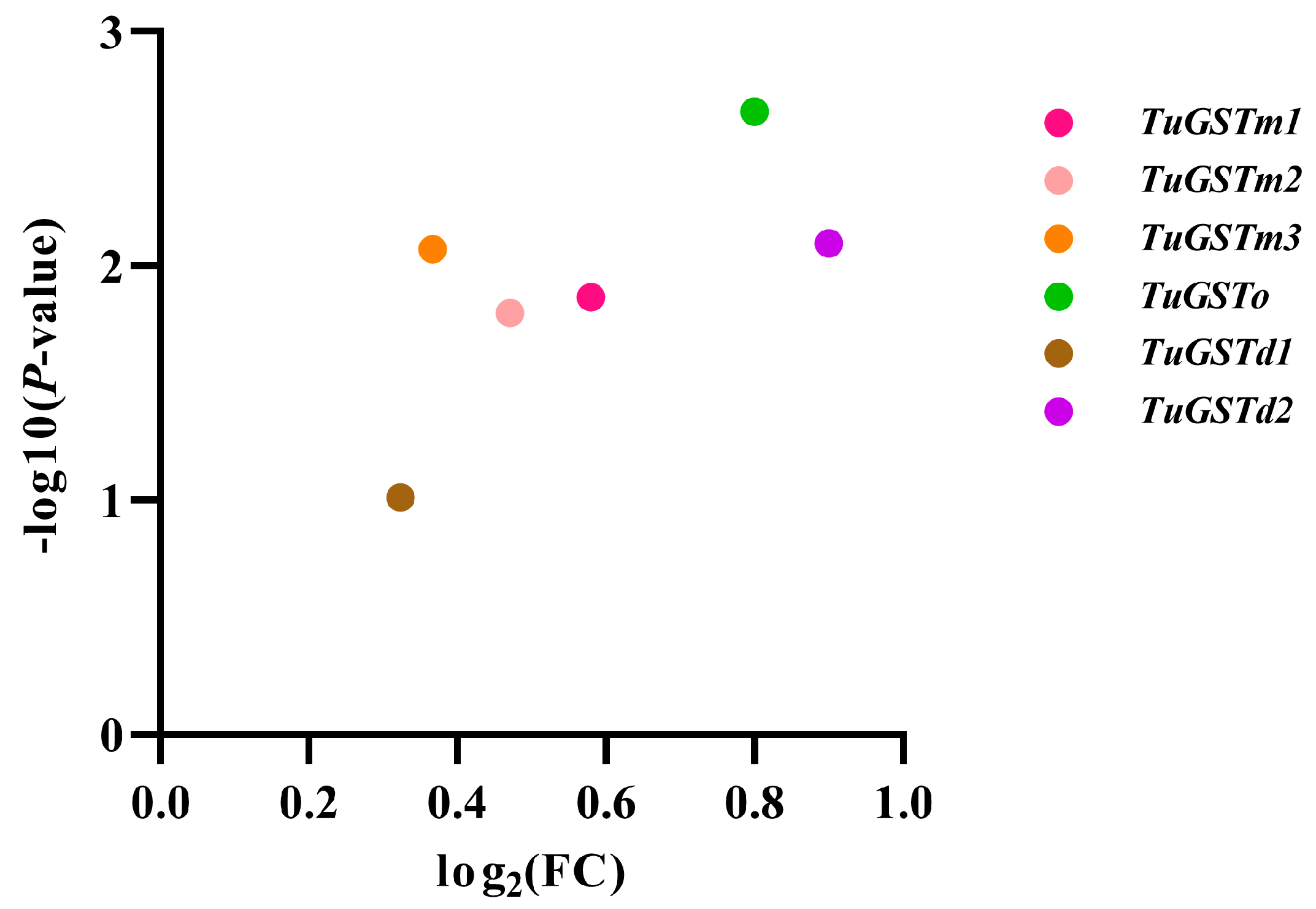
3.1. Selection and Identification of GST Genes in T. urticae
3.2. Sequence Analysis of Six GST Genes
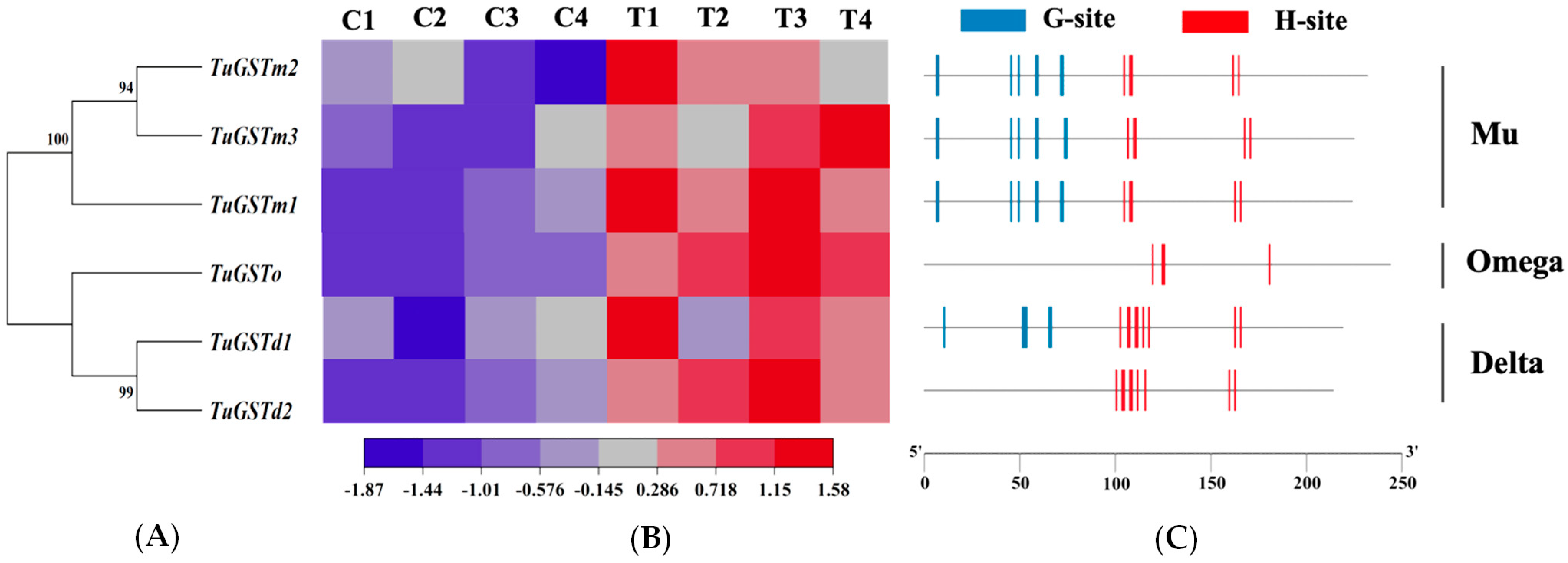
3.3. Structural Characterization of Six GST Genes
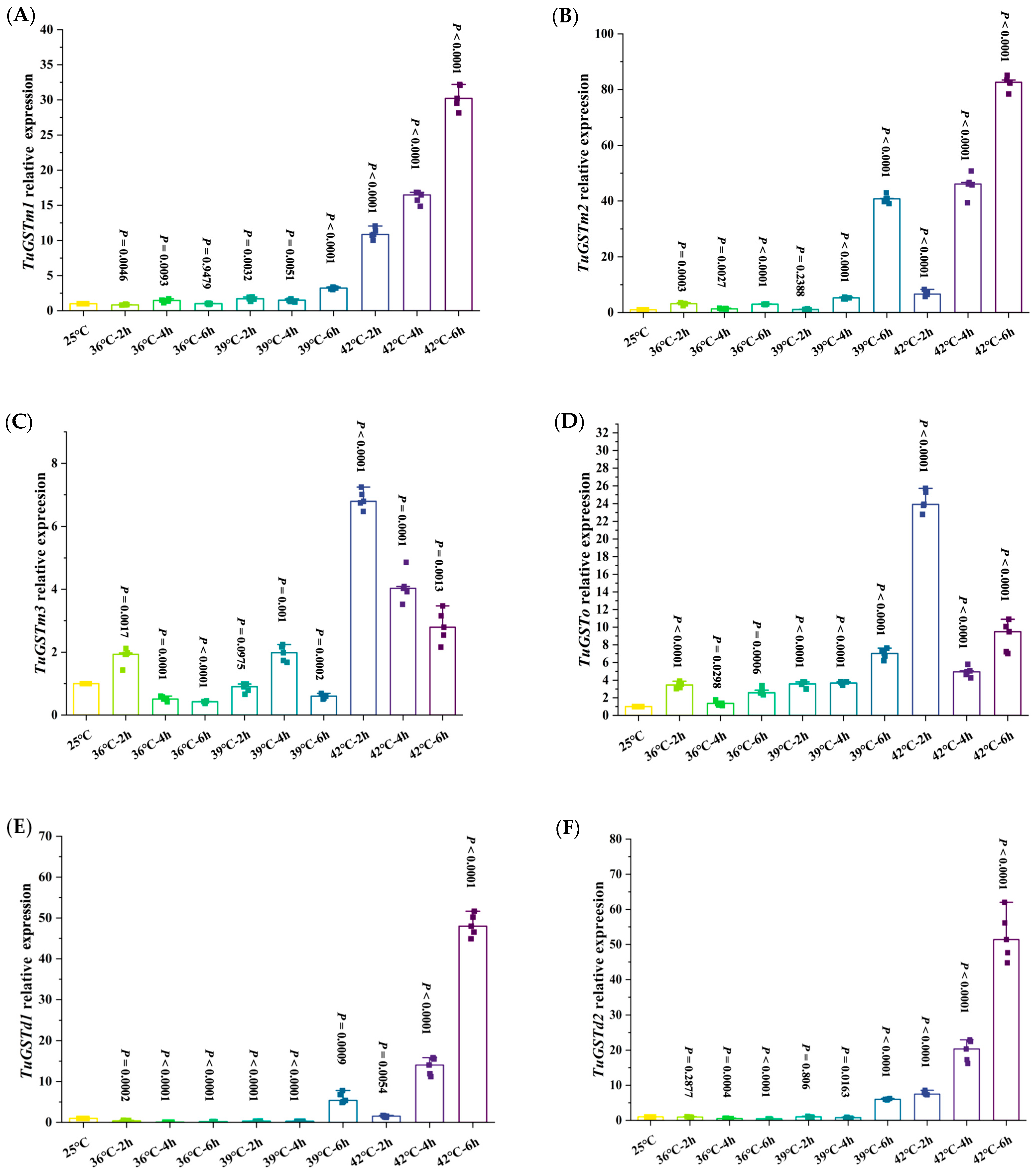
3.4. Transcriptional Expression of Six GST Genes under Different Heat Stress Conditions
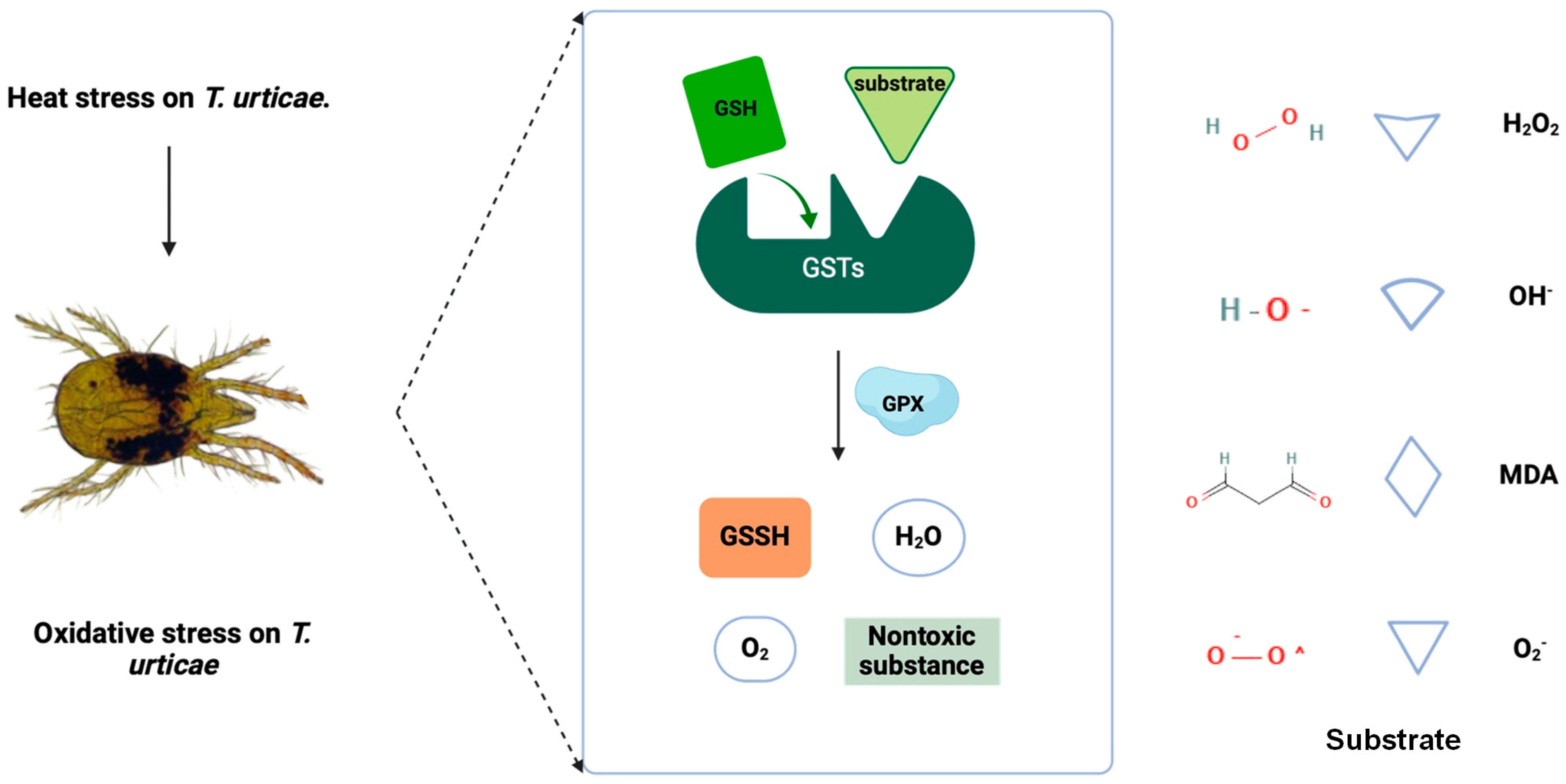
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shang, S.Q.; Chang, Y.; Li, W.Z.; Wang, C.Q.; Nie, P.C. Effects of B-azolemiteacrylic on life-history traits and demographic parameters of two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae). Exp. Appl. Acarol. 2022, 86, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Sousa, V.C.; Zélé, F.; Rodrigues, L.R.; Godinho, D.P.; de la Masselière, M.C.; Magalhães, S. Rapid host-plant adaptation in the herbivorous spider mite Tetranychus urticae occurs at low cost. Curr. Opin. Insect Sci. 2019, 36, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Wermelinger, B.; Oertli, J.J.; Baumgärtner, J. Environmental factors affecting the life-tables of Tetranychus urticae (Acari: Tetranychidae) III. Host-plant nutrition. Exp. Appl. Acarol. 1991, 12, 259–274. [Google Scholar] [CrossRef]
- Savi, P.J.; Gonsaga, R.F.; de Matos, S.T.S.; Braz, L.T.; de Moraes, G.J.; de Andrade, D.J. Performance of Tetranychus urticae (Acari: Tetranychidae) on three hop cultivars (Humulus lupulus). Exp Appl Acarol. 2021, 84, 733–753. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Kim, M.J.; Baek, S. Within-Plant Distribution of Two-Spotted Spider Mites, Tetranychus urticae Koch (Acari: Tetranychidae), on Strawberries: Decision of an Optimal Sampling Unit. Insects 2022, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Gerson, U. Spider Mites: Their Biology. Natural Enemies and Control; Helle, W., Sabelis, M.W., Eds.; Elsevier: Amsterdam, The Netherlands; New York, NY, USA, 1985; Volume 1. [Google Scholar]
- Tetsuo, G.; Daisuke, M.; Gösta, N. Development and reproduction of five Tetranychus species (Acari: Tetranychidae): Do they all have the potential to become major pests? Exp. Appl. Acarol. 2015, 66, 453–479. [Google Scholar]
- Tscholl, T.; Nachman, G.; Spangl, B.; Scalmani, I.; Walzer, A. Parental exposure to heat waves improves offspring reproductive investment in Tetranychus urticae (Acari: Tetranychidae), but not in its predator, Phytoseiulus persimilis (Acari: Phytoseiidae). Ecol. Evol. 2023, 13, e10748. [Google Scholar] [CrossRef] [PubMed]
- Nie, P.C.; Yang, R.L.; Zhou, J.J.; Dewer, Y.; Shang, S.Q. Elucidating the Effect of Temperature Stress on the Protein Content, Total Antioxidant Capacity, and Antioxidant Enzyme Activities in Tetranychus urticae (Acari: Tetranychidae). Insects 2023, 14, 429. [Google Scholar] [CrossRef]
- Forouzanfar, M.H.; Afshin, A.; Alexander, L.T.; Anderson, H.R.; Bhutta, Z.A.; Biryukov, S.; Brauer, M.; Burnett, R.; Cercy, K.; Charlson, F.J.; et al. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1659–1724. [Google Scholar] [CrossRef] [PubMed]
- Ozougwu, J.C. The role of reactive oxygen species and antioxidants in oxidative stress. Int. J. Res. 2016, 1, 1–8. [Google Scholar]
- Aruoma, O.I. Free radicals, oxidative stress, and antioxidants in human health and disease. J. Am. Oil Chem. Soc. 1998, 75, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Fairley, L.H.; Das, S.; Dharwal, V.; Amorim, N.; Hegarty, K.J.; Wadhwa, R.; Mounika, G.; Hansbro, P.M. Mitochondria-Targeted Antioxidants as a Therapeutic Strategy for Chronic Obstructive Pulmonary Disease. Antioxidants 2023, 12, 973. [Google Scholar] [CrossRef] [PubMed]
- Scandalios, J.G. The rise of ROS. Trends Biochem. Sci. 2002, 27, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Tuzet, A.; Rahantaniaina, M.S.; Noctor, G. Analyzing the function of catalase and the ascorbate–glutathione pathway in H2O2 processing: Insights from an experimentally constrained kinetic model. Antioxid. Redox Signal. 2019, 30, 1238–1268. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Ding, C.; Chen, L.; Yang, B.; Li, M.; Wang, X.; Su, F.; Liu, C.; Huang, Y. Multienzyme-Mimicking Au@Cu2O with Complete Antioxidant Capacity for Reactive Oxygen Species Scavenging. ACS Appl. Mater. Interfaces 2023, 15, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Board, P.G.; Menon, D. Glutathione transferases, regulators of cellular metabolism and physiology. Biochim. Biophys. Acta BBA-Gen. Subj. 2013, 1830, 3267–3288. [Google Scholar] [CrossRef] [PubMed]
- Fragoso, D.B.; Guedes, R.N.C.; Rezende, S.T. Glutathione S-transferase detoxification as a potential pyrethroid resistance mechanism in the maize weevil Sitophilus zeamais. Entomol. Exp. Appl. 2003, 109, 21–29. [Google Scholar] [CrossRef]
- Heidel-Fischer, H.M.; Vogel, H. Molecular mechanisms of insect adaptation to plant secondary compounds. Curr. Opin. Insect Sci. 2015, 8, 8–14. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, H.; Lu, W.; Li, Y.; Guo, X.; Xu, B. A novel Omega-class glutathione S-transferase gene in Apis cerana cerana: Molecular characterisation of GSTO2 and its protective effects in oxidative stress. Cell Stress Chaperones 2013, 18, 503–516. [Google Scholar] [CrossRef]
- BK, S.K.; Moural, T.; Zhu, F. Functional and structural diversity of insect glutathione S-transferases in xenobiotic adaptation. Int. J. Biol. Sci. 2022, 18, 5713. [Google Scholar]
- Simon, J.Y. Insect glutathione S-transferases. Zool. Stud. 1996, 35, 9–19. [Google Scholar]
- Giorgio, M.; Trinei, M.; Migliaccio, E.; Pelicci, P.G. Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals? Nat. Rev. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, F.; Jia, H.; Yan, Y.; Wang, H.; Guo, X.; Xu, B. A glutathione S-transferase gene associated with antioxidant properties isolated from Apis cerana cerana. Sci. Nat. 2016, 103, 43. [Google Scholar] [CrossRef] [PubMed]
- Grbić, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Z.; Kang, W.J.; Zhou, J.J.; Shang, S.Q.; Shi, S.L. The antennal transcriptome analysis and characterizations of odorant-binding proteins in Megachile saussurei (Hymenoptera, Megachilidae). BMC Genom. 2023, 24, 781. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Van den Boom, C.E.M.; Van Beek, T.A.; Dicke, M. Differences among plant species in acceptance by the spider mite Tetranychus urticae Koch. J. Appl. Entomol. 2003, 127, 177–183. [Google Scholar] [CrossRef]
- Praslička, J.; Huszár, J. Influence of temperature and host plants on the development and fecundity of the spider mite Tetranychus urticae (Acarina: Tetranychidae). Plant Prot. Sci. 2004, 40, 141. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Tirry, L. Mechanisms of acaricide resistance in the two-spotted spider mite Tetranychus urticae. In Biorational Control of Arthropod Pests: Application and Resistance Management; Springer: Dordrecht, The Netherlands, 2009; pp. 347–393. [Google Scholar]
- Bounfour, M.; Tanigoshi, L.K. Effect of temperature on development and demographic parameters of Tetranychus urticae and Eotetranychus carpini borealis (Acari: Tetranychidae). Ann. Entomol. 2001, 94, 400–404. [Google Scholar] [CrossRef]
- Friedman, R. Genomic organization of the glutathione S-transferase family in insects. Mol. Phylogenet. Evol. 2011, 61, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Q.; Zhang, M.X.; Yu, J.Y.; Jin, Y.; Ling, B.; Du, J.P.; Li, G.H.; Qin, Q.M.; Cai, Q.N. Glutathione S-transferase of brown planthoppers (Nilaparvata lugens) is essential for their adaptation to gramine-containing host plants. PLoS ONE 2013, 8, e64026. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Strange, R.C. Potential contribution of the glutathione S-transferase supergene family to resistance to oxidative stress. Free Radic. Res. 1995, 22, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.M.; Hooftman, A.; Angiari, S.; Tummala, P.; Zaslona, Z.; Runtsch, M.C.; McGettrick, A.F.; Sutton, C.E.; Diskin, C.; Rooke, M.; et al. Glutathione Transferase Omega-1 Regulates NLRP3 Inflammasome Activation through NEK7 Deglutathionylation. Cell Rep. 2019, 29, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Kim, S.H.; Kim, J.; Kim, H.; Yim, J. Glutathione s-transferase omega 1 activity is sufficient to suppress neurodegeneration in a Drosophila model of Parkinson disease. J. Biol. Chem. 2012, 287, 6628–6641. [Google Scholar] [CrossRef] [PubMed]
- Škerlová, J.; Lindström, H.; Gonis, E.; Sjödin, B.; Neiers, F.; Stenmark, P.; Mannervik, B. Structure and steroid isomerase activity of Drosophila glutathione transferase E14 essential for ecdysteroid biosynthesis. FEBS Lett. 2020, 594, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Saisawang, C.; Wongsantichon, J.; Ketterman, A.J. A preliminary characterization of the cytosolic glutathione transferase proteome from Drosophila melanogaster. Biochem. J. 2012, 442, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Xu, Z.; Zou, H.; Liu, J.; Chen, S.; Feng, Q.; Zheng, S. Glutathione S-transferase SlGSTE1 in Spodoptera litura may be associated with feeding adaptation of host plants. Insect Biochem. Mol. Biol. 2016, 70, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Fan, D.; Liu, L.; Feng, J. Identification and Characterization of Glutathione S-Transferase Genes in the Antennae of Codling Moth (Lepidoptera: Tortricidae). Ann. Entomol. 2017, 110, 409–416. [Google Scholar] [CrossRef]
- Ma, M.; Zhang, Y.X.; Chen, D.; Smagghe, G.; Wang, J.J.; Wei, D. Functional characterization of a glutathione S-transferase gene GSTe10 that contributes to ovarian development in Bactrocera dorsalis (Hendel). Entomol. Gen. 2022, 42, 539–547. [Google Scholar] [CrossRef]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef]
- Vaish, S.; Gupta, D.; Mehrotra, R.; Mehrotra, S.; Basantani, M.K. Glutathione S-transferase: A versatile protein family. 3 Biotech. 2020, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, M.K.; Shobbar, Z.S.; Shahbazi, M.; Abedini, R.; Zare, S. Glutathione S-transferase (GST) family in barley: Identification of members, enzyme activity, and gene expression pattern. J. Plant Physiol. 2013, 170, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Raza, H. Dual localization of glutathione S-transferase in the cytosol and mitochondria: Implications in oxidative stress, toxicity and disease. FEBS J. 2011, 278, 4243–4251. [Google Scholar] [CrossRef]
- Zhu, T.; Li, W.; Xue, H.; Dong, S.; Wang, J.; Shang, S.; Dewer, Y. Selection, Identification, and Transcript Expression Analysis of Antioxidant Enzyme Genes in Neoseiulus barkeri after Short-Term Heat Stress. Antioxidants 2023, 12, 1998. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ORF | aa | Formula | Molecular Weight (kDa) | Theoretical pI |
|---|---|---|---|---|---|
| TuGSTm1 | 672 | 224 | C1207H1829N303O346S6 | 26.313 | 6.00 |
| TuGSTm2 | 696 | 232 | C1233H1856N302O358S3 | 26.734 | 5.01 |
| TuGSTm3 | 675 | 225 | C1193H1846N292O353S7 | 26.152 | 4.71 |
| TuGSTd1 | 657 | 219 | C1123H1752N280O327S4 | 24.536 | 5.82 |
| TuGSTd2 | 642 | 192 | C1015H1575N259O293S11 | 22.447 | 5.49 |
| TuGSTo | 732 | 244 | C1290H1987N307O365S10 | 27.957 | 6.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, T.; Wei, B.; Wang, Y.; Shang, S. Glutathione S-Transferase Genes Involved in Response to Short-Term Heat Stress in Tetranychus urticae (Koch). Antioxidants 2024, 13, 442. https://doi.org/10.3390/antiox13040442
Zhu T, Wei B, Wang Y, Shang S. Glutathione S-Transferase Genes Involved in Response to Short-Term Heat Stress in Tetranychus urticae (Koch). Antioxidants. 2024; 13(4):442. https://doi.org/10.3390/antiox13040442
Chicago/Turabian StyleZhu, Tong, Bin Wei, Yue Wang, and Suqin Shang. 2024. "Glutathione S-Transferase Genes Involved in Response to Short-Term Heat Stress in Tetranychus urticae (Koch)" Antioxidants 13, no. 4: 442. https://doi.org/10.3390/antiox13040442
APA StyleZhu, T., Wei, B., Wang, Y., & Shang, S. (2024). Glutathione S-Transferase Genes Involved in Response to Short-Term Heat Stress in Tetranychus urticae (Koch). Antioxidants, 13(4), 442. https://doi.org/10.3390/antiox13040442