
Biochemical and Molecular Insights into Variation in Sesame Seed Antioxidant Capability as Revealed by Metabolomics and Transcriptomics Analysis

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
2.2. Assessment of Total Phenolic (TPC) and Flavonoid (TFC) Contents and Antioxidant Activity (AOA)
2.3. Polyphenol Extraction and UPLC-MS/MS Analysis
2.4. Identification and Quantification of Phenolic Compounds
2.5. Data Analysis
2.6. RNA Extraction, Library Construction, Sequencing, and Alignment
2.7. Differentially Expressed Genes (DEGs) and Functional Enrichment Analysis
2.8. Quantitative RT–PCR Analysis
3. Results and Discussion
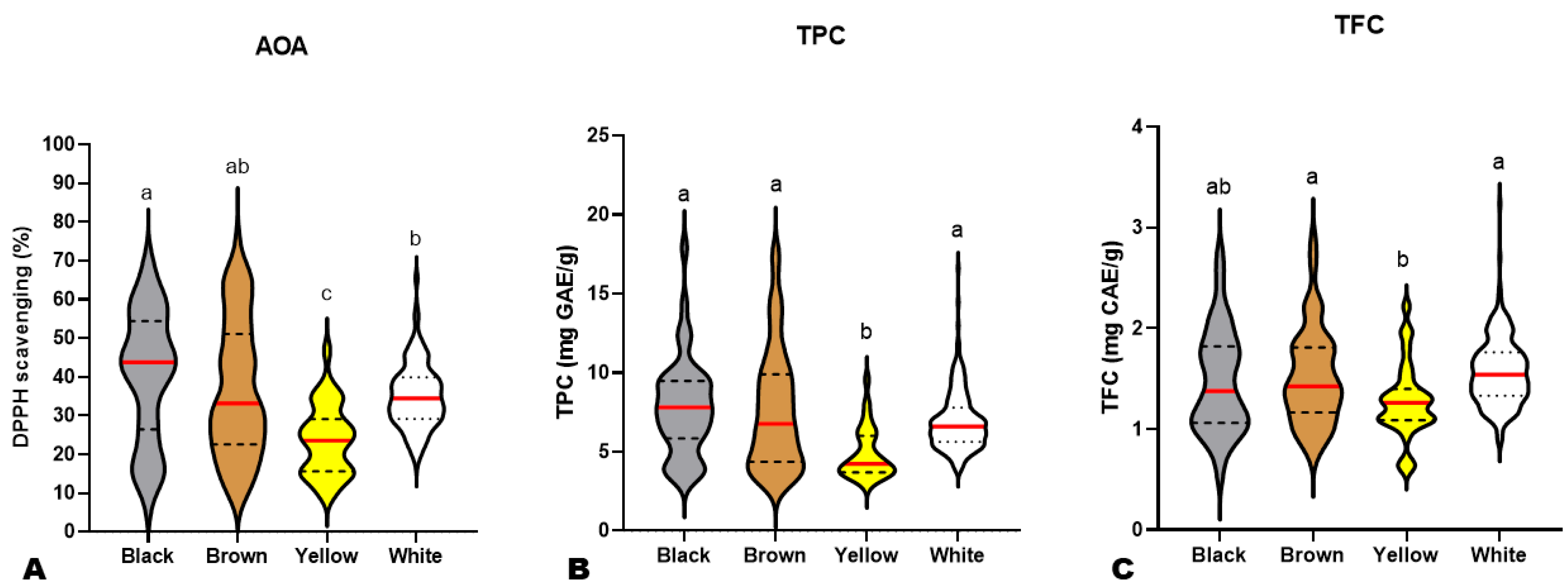
3.1. Variation in Antioxidant Activity (AOA), Total Flavonoid (TFC), and Phenolic (TPC) Contents in the Sesame Population
3.2. Correlation between Seed Antioxidant Activity and Seed Phytochemicals
3.3. Influence of Seed Coat Colors on Sesame Seed Antioxidant Activity
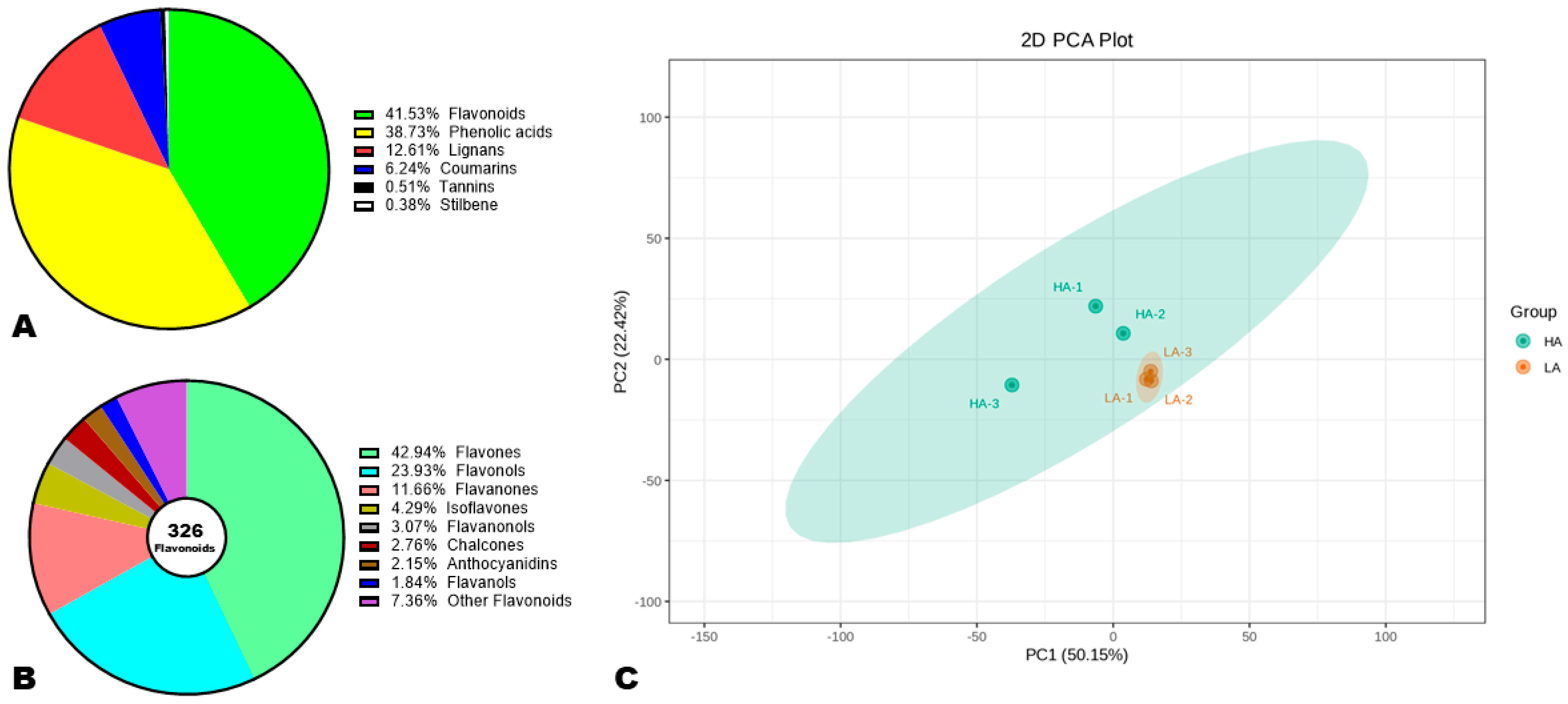
3.4. Polyphenol Profiles of High and Low Antioxidant Sesame Seeds
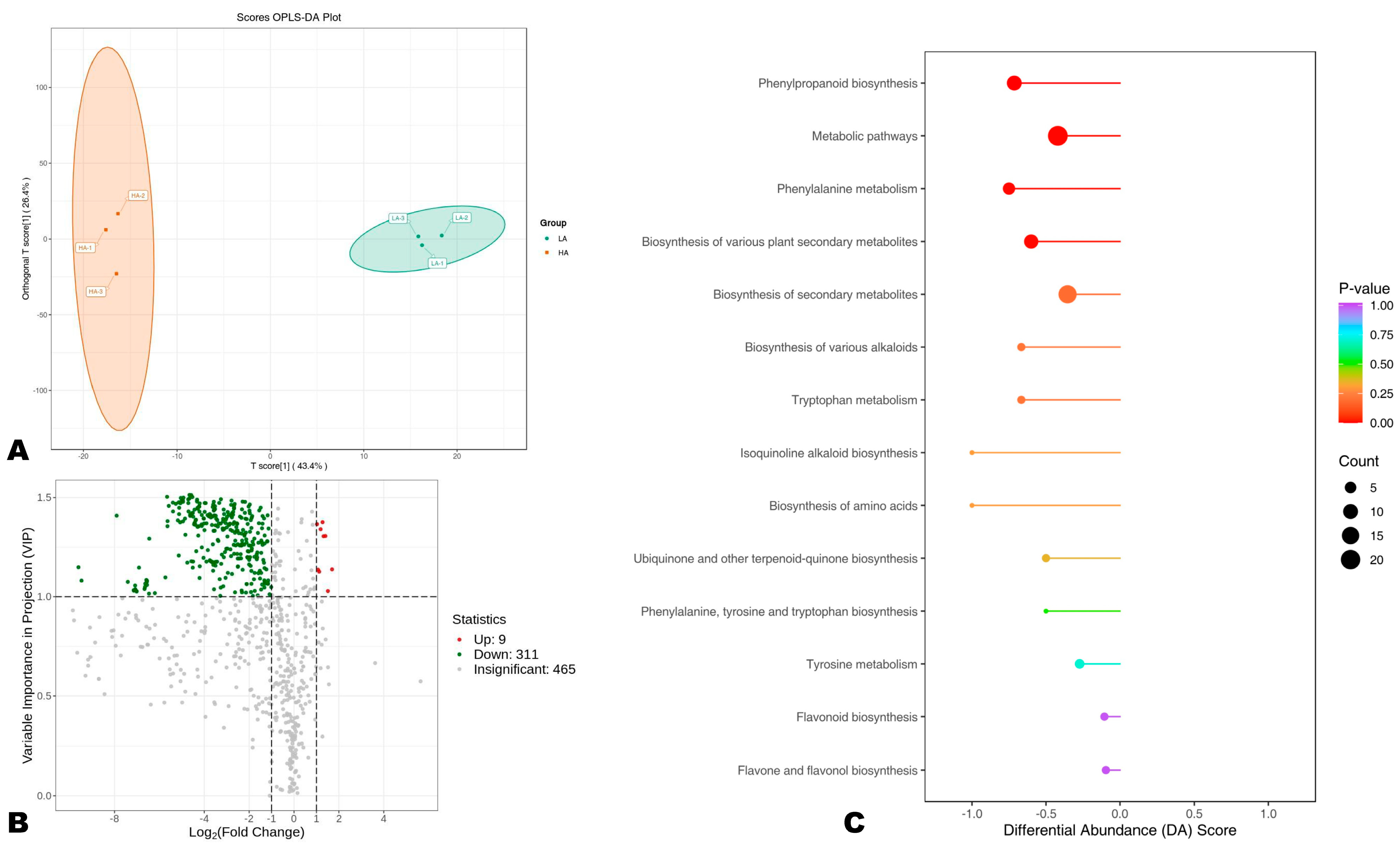
3.5. Differentially Accumulated Phenolic (DAPs) Compounds and KEGG Analysis
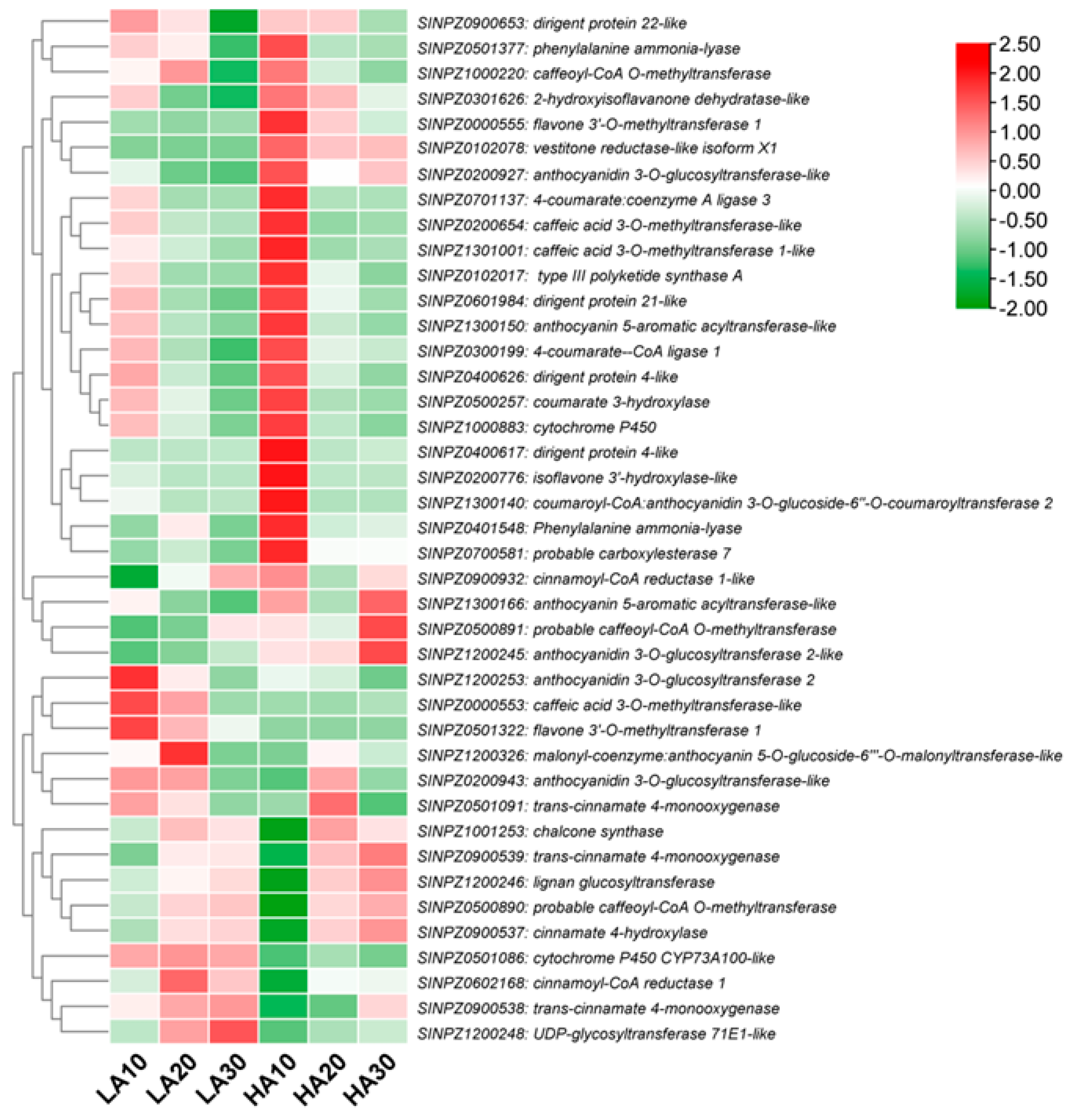
3.6. Major Highly Accumulated Phenolic Compounds in High-Antioxidant Sesame Seeds
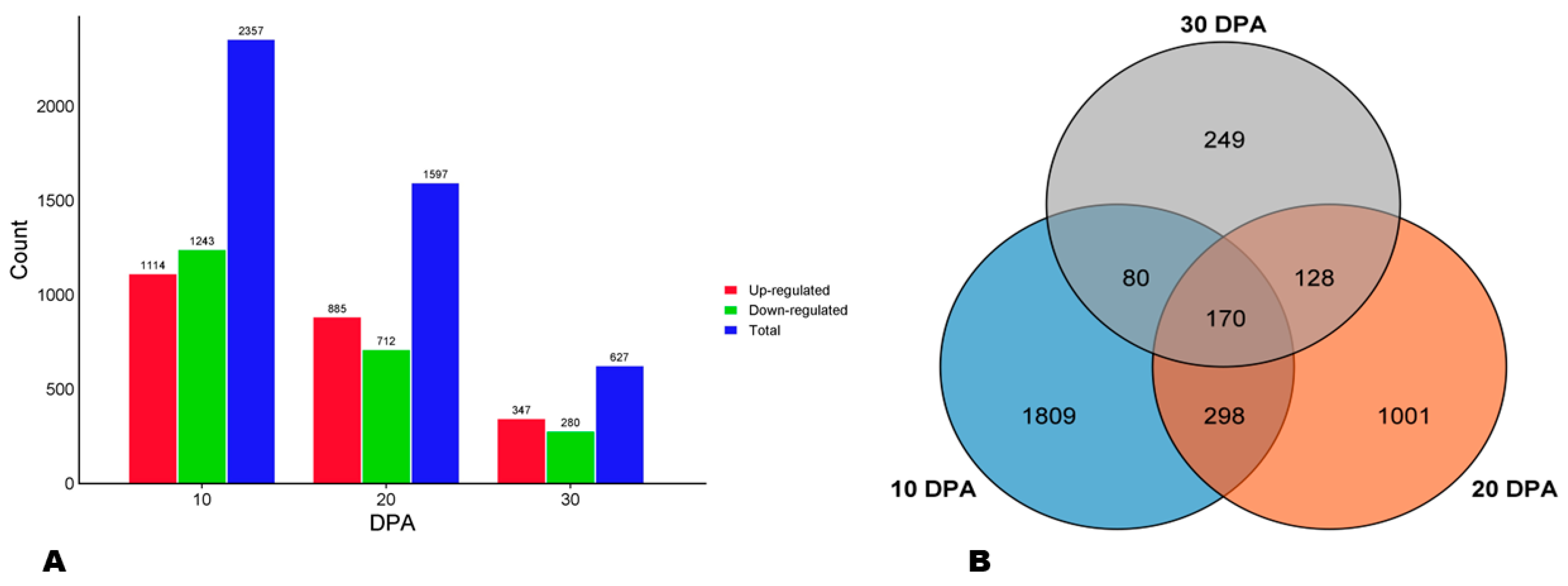
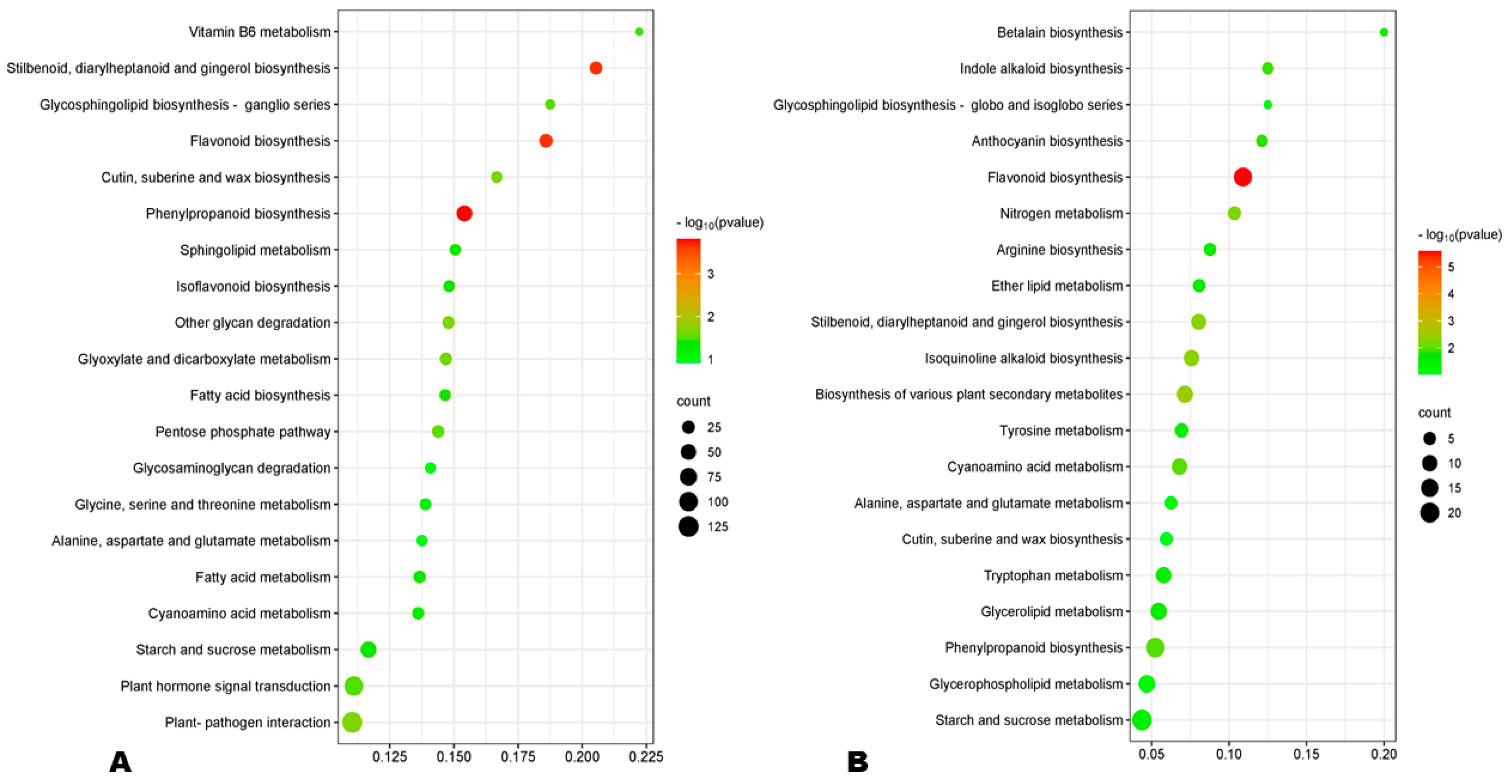
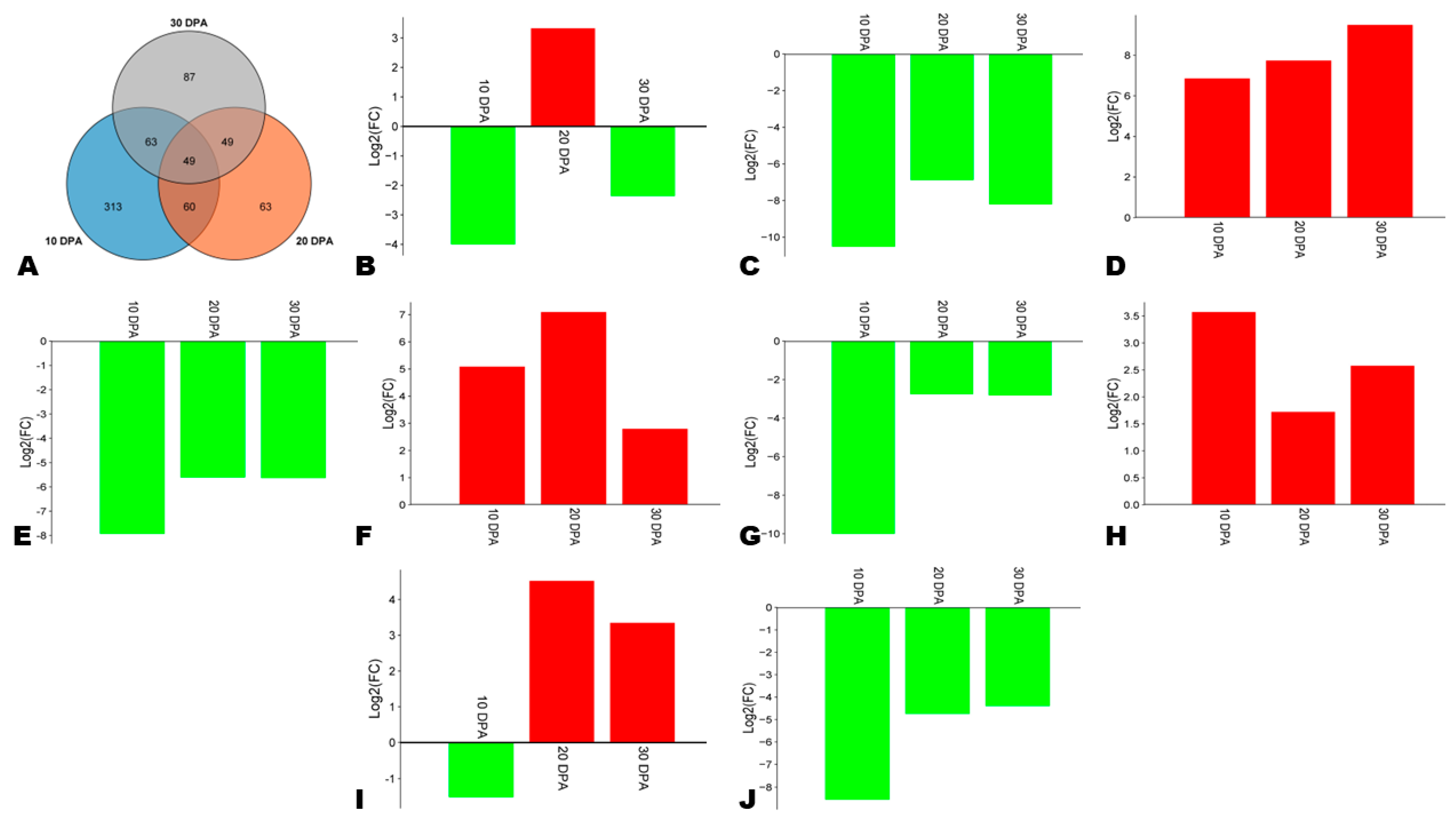
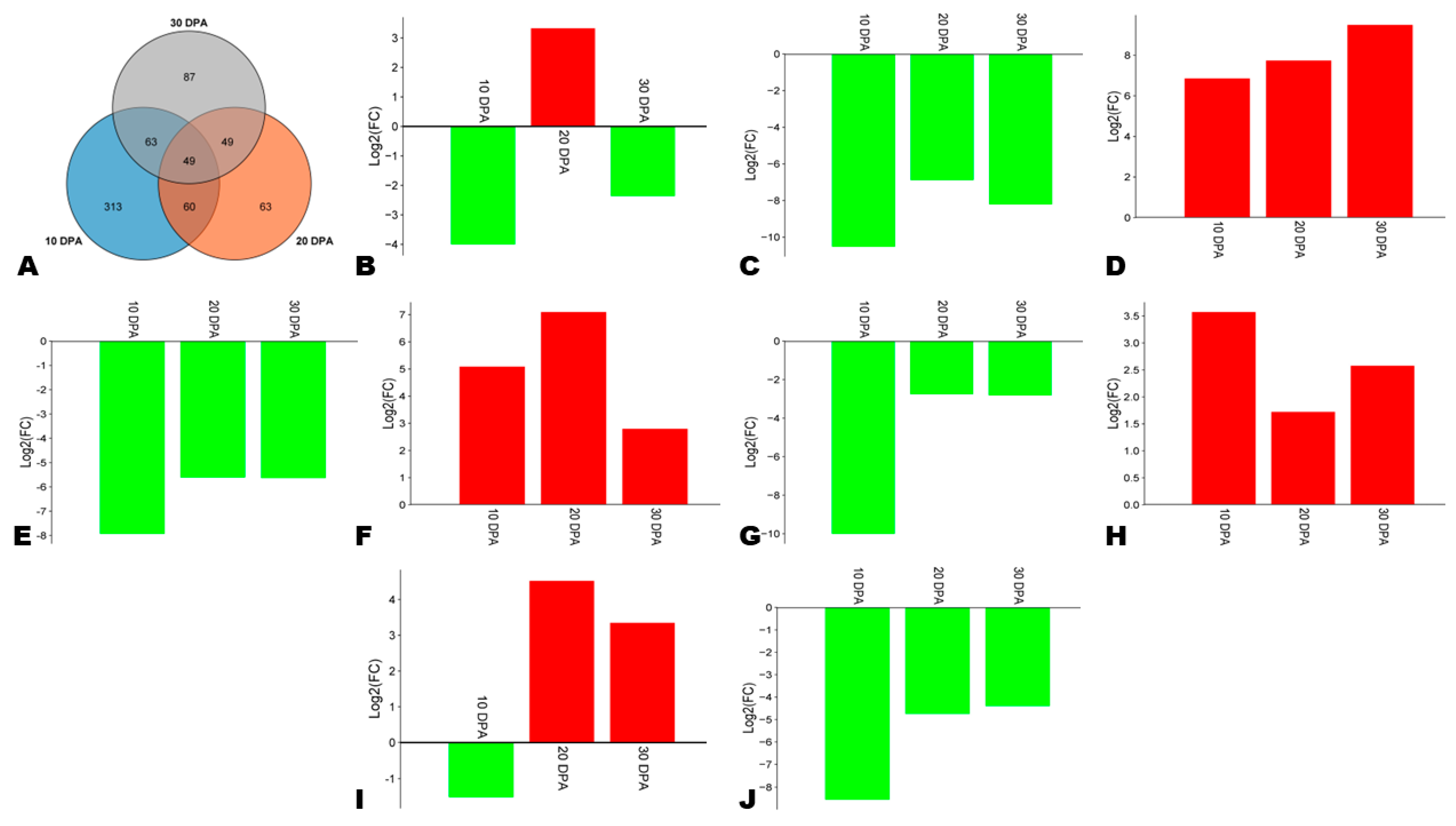
3.7. Differentially Expressed Genes (DEGs) between HA and LA during Seed Development
3.8. Expression Patterns of Phenylpropanoid Biosynthesis-Related DEGs
3.9. Key Differentially Expressed Transcription Factors (TFs)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Langyan, S.; Yadava, P.; Sharma, S.; Gupta, N.C.; Bansal, R.; Yadav, R.; Kalia, S.; Kumar, A. Food and Nutraceutical Functions of Sesame Oil: An Underutilized Crop for Nutritional and Health Benefits. Food Chem. 2022, 389, 132990. [Google Scholar] [CrossRef] [PubMed]
- Anilakumar, K.R.; Pal, A.; Khanum, F.; Bawa, A.S. Nutritional, Medicinal and Industrial Uses of Sesame (Sesamum indicum L.) Seeds—An Overview. Agric. Conspec. Sci. 2010, 75, 159–168. [Google Scholar]
- Wei, P.; Zhao, F.; Wang, Z.; Wang, Q.; Chai, X.; Hou, G.; Meng, Q. Sesame (Sesamum indicum L.): A Comprehensive Review of Nutritional Value, Phytochemical Composition, Health Benefits, Development of Food, and Industrial Applications. Nutrients 2022, 14, 4079. [Google Scholar] [CrossRef] [PubMed]
- Oboulbiga, E.B.; Douamba, Z.; Compaoré-Sérémé, D.; Semporé, J.N.; Dabo, R.; Semde, Z.; Tapsoba, F.W.B.; Hama-Ba, F.; Songré-Ouattara, L.T.; Parkouda, C.; et al. Physicochemical, Potential Nutritional, Antioxidant and Health Properties of Sesame Seed Oil: A Review. Front. Nutr. 2023, 10, 1127926. [Google Scholar] [CrossRef] [PubMed]
- Andargie, M.; Vinas, M.; Rathgeb, A.; Möller, E.; Karlovsky, P. Lignans of Sesame (Sesamum indicum L.): A Comprehensive Review. Molecules 2021, 26, 883. [Google Scholar] [CrossRef] [PubMed]
- Pathak, N.; Rai, A.K.; Kumari, R.; Bhat, K.V. Value Addition in Sesame: A Perspective on Bioactive Components for Enhancing Utility and Profitability. Pharmacogn. Rev. 2014, 8, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Li, H.; Fu, G.; Chen, X.; Chen, F.; Xie, M. The Relationship of Antioxidant Components and Antioxidant Activity of Sesame Seed Oil. J. Sci. Food Agric. 2015, 95, 2571–2578. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.A.; Hameed, A.; Ajmal, I.; Nosheen, S.; Suleria, H.A.R.; Song, Y. Effects of Sesame Seed Extract as a Natural Antioxidant on the Oxidative Stability of Sunflower Oil. J. Food Sci. Technol. 2018, 55, 4099–4110. [Google Scholar] [CrossRef] [PubMed]
- Karshenas, M.; Goli, M.; Zamindar, N. The Effect of Replacing Egg Yolk with Sesame–Peanut Defatted Meal Milk on the Physicochemical, Colorimetry, and Rheological Properties of Low-Cholesterol Mayonnaise. Food Sci. Nutr. 2018, 6, 824–833. [Google Scholar] [CrossRef]
- Kim, J.H.; Seo, W.D.; Lee, S.K.; Lee, Y.B.; Park, C.H.; Ryu, H.W.; Lee, J.H. Comparative Assessment of Compositional Components, Antioxidant Effects, and Lignan Extractions from Korean White and Black Sesame (Sesamum indicum L.) Seeds for Different Crop Years. J. Funct. Foods 2014, 7, 495–505. [Google Scholar] [CrossRef]
- Abib, B.; Afifi, S.M.; El-Din, M.G.S.; Farag, M.A. How Do Cultivar Origin and Stepwise Industrial Processing Impact Sesamum Indicum Seeds’ Metabolome and Its Paste and in Relation to Their Antioxidant Effects? A Case Study from the Sesame Industry. Food Chem. 2023, 420, 136134. [Google Scholar] [CrossRef]
- Arab, R.; Casal, S.; Pinho, T.; Cruz, R.; Freidja, M.L.; Lorenzo, J.M.; Hano, C.; Madani, K.; Boulekbache-Makhlouf, L. Effects of Seed Roasting Temperature on Sesame Oil Fatty Acid Composition, Lignan, Sterol and Tocopherol Contents, Oxidative Stability and Antioxidant Potential for Food Applications. Molecules 2022, 27, 4508. [Google Scholar] [CrossRef] [PubMed]
- Ruslan, K.; Happyniar, S.; Fidrianny, I. Antioxidant Potential of Two Varieties of Sesamum indicum L. Collected from Indonesia. J. Taibah Univ. Med. Sci. 2018, 13, 211–218. [Google Scholar] [CrossRef]
- Dossou, S.S.K.; Xu, F.; You, J.; Zhou, R.; Li, D.; Wang, L. Widely Targeted Metabolome Profiling of Different Colored Sesame (Sesamum indicum L.) Seeds Provides New Insight into Their Antioxidant Activities. Food Res. Int. 2022, 151, 110850. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Massri, M.; Nasrallah, G.K. A Comprehensive Review on the Anti-Cancer Properties and Mechanisms of Action of Sesamin, a Lignan in Sesame Seeds (Sesamum indicum). Eur. J. Pharmacol. 2017, 815, 512–521. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Mansour, Z.R. Sesamol, a Major Lignan in Sesame Seeds (Sesamum indicum): Anti-Cancer Properties and Mechanisms of Action. Eur. J. Pharmacol. 2019, 855, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Liyana-Pathirana, C.M.; Wall, D.S. Antioxidant Activity of White and Black Sesame Seeds and Their Hull Fractions. Food Chem. 2006, 99, 478–483. [Google Scholar] [CrossRef]
- Zhou, L.; Lin, X.; Abbasi, A.M.; Zheng, B. Phytochemical Contents and Antioxidant and Antiproliferative Activities of Selected Black and White Sesame Seeds. Biomed Res. Int. 2016, 2016, 8495630. [Google Scholar] [CrossRef]
- Lang, Y.; Gao, N.; Zang, Z.; Meng, X.; Lin, Y.; Yang, S.; Yang, Y.; Jin, Z.; Li, B. Classification and Antioxidant Assays of Polyphenols: A Review. J. Future Foods 2024, 4, 193–204. [Google Scholar] [CrossRef]
- Stagos, D. Antioxidant Activity of Polyphenolic Plant Extracts. Antioxidants 2019, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Shahrajabian, M.H. Therapeutic Potential of Phenolic Compounds in Medicinal Plants—Natural Health Products for Human Health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Matsumura, Y.; Kitabatake, M.; Kayano, S.I.; Ito, T. Dietary Phenolic Compounds: Their Health Benefits and Association with the Gut Microbiota. Antioxidants 2023, 12, 880. [Google Scholar] [CrossRef]
- Long, Z.; Xiang, W.; He, Q.; Xiao, W.; Wei, H.; Li, H.; Guo, H.; Chen, Y.; Yuan, M.; Yuan, X.; et al. Efficacy and Safety of Dietary Polyphenols in Rheumatoid Arthritis: A Systematic Review and Meta-Analysis of 47 Randomized Controlled Trials. Front. Immunol. 2023, 14, 1024120. [Google Scholar] [CrossRef]
- Zeb, A. A Comprehensive Review on Different Classes of Polyphenolic Compounds Present in Edible Oils. Food Res. Int. 2021, 143, 110312. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, H.; Lv, X.; Wang, D.; Chen, H.; Wei, F. Comprehensive Review of Composition Distribution and Advances in Profiling of Phenolic Compounds in Oilseeds. Front. Nutr. 2022, 9, 1044871. [Google Scholar] [CrossRef]
- Anwar, M.; Chen, L.; Xiao, Y.; Wu, J.; Zeng, L.; Li, H.; Wu, Q.; Hu, Z. Recent Advanced Metabolic and Genetic Engineering of Phenylpropanoid Biosynthetic Pathways. Int. J. Mol. Sci. 2021, 22, 9544. [Google Scholar] [CrossRef]
- Santos, E.L.; Maia, B.H.L.N.S.; Ferriani, A.P.; Teixeira, S.D. Flavonoids: Classification, Biosynthesis and Chemical Ecology. In Flavonoids—From Biosynthesis to Human Health; InTech: London, UK, 2017; pp. 3–16. [Google Scholar]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The Flavonoid Biosynthetic Pathway in Arabidopsis: Structural and Genetic Diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Yang, G.; Li, L.; Wei, M.; Li, J.; Yang, F. SmMYB113 Is a Key Transcription Factor Responsible for Compositional Variation of Anthocyanin and Color Diversity Among Eggplant Peels. Front. Plant Sci. 2022, 13, 843996. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.K.; Zheng, L.; Liu, R.J.; Chang, M.; Jin, Q.Z.; Wang, X.G. Chemical Characterization, Oxidative Stability, and In Vitro Antioxidant Capacity of Sesame Oils Extracted by Supercritical and Subcritical Techniques and Conventional Methods: A Comparative Study Using Chemometrics. Eur. J. Lipid Sci. Technol. 2018, 120, 1700326. [Google Scholar] [CrossRef]
- Choi, G.-Y.; Han, Y.-S.; Sim, K.-H.; Kim, M.-H. Phenolic Compounds, Antioxidant Capacity, and α-Amylase and α-Glucosidase Inhibitory Activity of Ethanol Extracts of Perilla Seed Meal. Food Sci. Nutr. 2023, 11, 4596–4606. [Google Scholar] [CrossRef] [PubMed]
- Kefale, H.; Segla Koffi Dossou, S.; Li, F.; Jiang, N.; Zhou, R.; Wang, L.; Zhang, Y.; Li, D.; You, J.; Wang, L. Widely Targeted Metabolic Profiling Provides Insights into Variations in Bioactive Compounds and Antioxidant Activity of Sesame, Soybean, Peanut, and Perilla. Food Res. Int. 2023, 174, 113586. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luoa, J. A Novel Integrated Method for Large-Scale Detection, Identification, and Quantification of Widely Targeted Metabolites: Application in the Study of Rice Metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cui, D.; Ma, X.; Han, B.; Han, L. Comparative Analysis of Rice Reveals Insights into the Mechanism of Colored Rice via Widely Targeted Metabolomics. Food Chem. 2022, 399, 133926. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Wu, S.; Chen, D.; Wu, X.; Cai, Q. Polyphenols and Phytohormones Profiling of Pre-Harvest Sprouting Resistant and Susceptible Wheat Genotypes. SN Appl. Sci. 2023, 5, 249. [Google Scholar] [CrossRef]
- Tang, D.; Chen, M.; Huang, X.; Zhang, G.; Zeng, L.; Zhang, G.; Wu, S.; Wang, Y. SRplot: A Free Online Platform for Data Visualization and Graphing. PLoS ONE 2023, 18, e0294236. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Dossou, S.S.K.; Meng, M.; Sheng, C.; Li, H.; Zhou, R.; Li, D.; Xu, P.; You, J.; Wang, L. Five Improved Sesame Reference Genomes and Genome Resequencing Unveil the Contribution of Structural Variants to Genetic Diversity and Yield-Related Traits Variation. Plant Biotechnol. J. 2023, 21, 1722–1724. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Kanani, P.; Shukla, Y.M.; Modi, A.R.; Subhash, N.; Kumar, S. Standardization of an Efficient Protocol for Isolation of RNA from Cuminum Cyminum. J. King Saud Univ. Sci. 2019, 31, 1202–1207. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, Q.; Dossou, S.S.K.; Zhou, R.; Zhao, Y.; Zhou, W.; Zhang, Y.; Li, D.; You, J.; Wang, L. Genome-Wide Association Study Uncovers Loci and Candidate Genes Underlying Phytosterol Variation in Sesame (Sesamum indicum L.). Agriculture 2022, 12, 392. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dossou, S.S.K.; Song, S.; Liu, A.; Li, D.; Zhou, R.; Berhe, M.; Zhang, Y.; Sheng, C.; Wang, Z.; You, J.; et al. Resequencing of 410 Sesame Accessions Identifies SINST1 as the Major Underlying Gene for Lignans Variation. Int. J. Mol. Sci. 2023, 24, 1055. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; You, J.; Liu, A.; Qi, X.; Li, D.; Zhao, Y.; Zhang, Y.; Zhang, L.; Zhang, X.; Li, P. Variation in Melatonin Contents and Genetic Dissection of Melatonin Biosynthesis in Sesame. Plants 2022, 11, 2005. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Song, S.; Segla Koffi Dossou, S.; Zhou, R.; Wei, X.; Wang, Z.; Sheng, C.; Zhang, Y.; You, J.; Wang, L. Genome-Wide Association Analysis and Transcriptome Reveal Novel Loci and a Candidate Regulatory Gene of Fatty Acid Biosynthesis in Sesame (Sesamum indicum L.). Plant Physiol. Biochem. PPB 2022, 186, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A Concise Overview on the Chemistry, Occurrence, and Human Health. Phyther. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [PubMed]
- Panzella, L.; Eidenberger, T.; Napolitano, A.; D’Ischia, M. Black Sesame Pigment: DPPH Assay-Guided Purification, Antioxidant/Antinitrosating Properties, and Identification of a Degradative Structural Marker. J. Agric. Food Chem. 2012, 60, 8895–8901. [Google Scholar] [CrossRef]
- Dossou, S.S.K.; Xu, F.; Dossa, K.; Zhou, R.; Zhao, Y.; Wang, L. Antioxidant Lignans Sesamin and Sesamolin in Sesame (Sesamum indicum L.): A Comprehensive Review and Future Prospects. J. Integr. Agric. 2023, 22, 14–30. [Google Scholar] [CrossRef]
- Maeda, H.; Dudareva, N. The Shikimate Pathway and Aromatic Amino Acid Biosynthesis in Plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nunes Nesi, A.; Araújo, W.L.; Braun, H.P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef]
- Qian, Y.; Lynch, J.H.; Guo, L.; Rhodes, D.; Morgan, J.A.; Dudareva, N. Completion of the Cytosolic Post-Chorismate Phenylalanine Biosynthetic Pathway in Plants. Nat. Commun. 2019, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.; Hatan, E.; Kumar, V.; Galsurker, O.; Nisim-Levi, A.; Ovadia, R.; Galili, G.; Lewinsohn, E.; Elad, Y.; Alkan, N.; et al. Increased Phenylalanine Levels in Plant Leaves Reduces Susceptibility to Botrytis Cinerea. Plant Sci. 2020, 290, 110289. [Google Scholar] [CrossRef]
- Poulev, A.; Heckman, J.R.; Raskin, I.; Belanger, F.C. Tricin Levels and Expression of Flavonoid Biosynthetic Genes in Developing Grains of Purple and Brown Pericarp Rice. PeerJ 2019, 7, e6477. [Google Scholar] [CrossRef]
- Cai, H.; Sale, S.; Britton, R.G.; Brown, K.; Steward, W.P.; Gescher, A.J. Pharmacokinetics in Mice and Metabolism in Murine and Human Liver Fractions of the Putative Cancer Chemopreventive Agents 3′,4′,5′,5,7- Pentamethoxyflavone and Tricin (4′,5,7-Trihydroxy-3′, 5′-Dimethoxyflavone). Cancer Chemother. Pharmacol. 2011, 67, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Kwon, H.J.; Jung, H.J. Tricin, 4′,5,7-Trihydroxy-3′,5′-Dimethoxyflavone, Exhibits Potent Antiangiogenic Activity in Vitro. Int. J. Oncol. 2016, 49, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Oak, C.; Khalifa, A.O.; Isali, I.; Bhaskaran, N.; Walker, E.; Shukla, S. Diosmetin Suppresses Human Prostate Cancer Cell Proliferation through the Induction of Apoptosis and Cell Cycle Arrest. Int. J. Oncol. 2018, 53, 835–843. [Google Scholar] [CrossRef]
- Choi, J.; Lee, D.H.; Park, S.Y.; Seol, J.W. Diosmetin Inhibits Tumor Development and Block Tumor Angiogenesis in Skin Cancer. Biomed. Pharmacother. 2019, 117, 109091. [Google Scholar] [CrossRef]
- Fang, J. Classification of Fruits Based on Anthocyanin Types and Relevance to Their Health Effects. Nutrition 2015, 31, 1301–1306. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, Y.; Li, Q. Matairesinol Exerts Anti-Inflammatory and Antioxidant Effects in Sepsis-Mediated Brain Injury by Repressing the MAPK and NF-ΚB Pathways through up-Regulating AMPK. Aging 2021, 13, 23780–23795. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, M.; Suryavanshi, S.; Bhowmick, S.; Alasmary, F.A.; Almutairi, T.M.; Islam, M.A.; Kaul-Ghanekar, R. Matairesinol, an Active Constituent of HC9 Polyherbal Formulation, Exhibits HDAC8 Inhibitory and Anticancer Activity. Biophys. Chem. 2021, 273, 106588. [Google Scholar] [CrossRef] [PubMed]
- Pratyusha, D.S.; Sarada, D.V.L. MYB Transcription Factors-Master Regulators of Phenylpropanoid Biosynthesis and Diverse Developmental and Stress Responses. Plant Cell Rep. 2022, 41, 2245–2260. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Min | Max | Mean | SD | SE | CV (%) | Skewness | Kurtosis |
|---|---|---|---|---|---|---|---|---|
| AOA (%) | 2.035 | 78.5 | 33.57 | 15.15 | 0.757 | 45.12 | 0.506 | 0.054 |
| TPC (mg GAE/g) | 2.717 | 21.98 | 6.9 | 3.072 | 0.154 | 44.52 | 1.478 | 3.415 |
| TFC (mg CAE/g) | 0.072 | 3.104 | 1.255 | 0.531 | 0.027 | 42.29 | 0.418 | 0.461 |
| Categories | Sub-Class | Compounds | Formula | VIP | p-Value | FDR | |Log2FC| |
|---|---|---|---|---|---|---|---|
| Flavonoids | Flavones | Tricin (5,7,4′-Trihydroxy-3′,5′-dimethoxyflavone) | C17H14O7 | 1.149 | 0.413 | 0.548 | 9.593 |
| Flavanones | Persicoside | C23H26O11 | 1.081 | 0.419 | 0.548 | 9.457 | |
| Flavones | 5,7,4′,5′-Tetrahydro-3′,6-dimethoxyflavone | C17H14O8 | 1.409 | 0.349 | 0.548 | 7.906 | |
| Flavones | 8-Methoxyapigenin | C16H12O6 | 1.075 | 0.415 | 0.548 | 7.409 | |
| Flavones | 6,7,8-Tetrahydroxy-5-methoxyflavone | C16H12O6 | 1.033 | 0.417 | 0.548 | 7.143 | |
| Flavones | 5,7,3′,5′-tetrahydroxy-6-methylfavanone | C16H12O6 | 1.030 | 0.417 | 0.548 | 7.138 | |
| Flavones | Scutevulin | C16H12O6 | 1.057 | 0.416 | 0.548 | 7.083 | |
| Isoflavones | Tectorigenin | C16H12O6 | 1.036 | 0.416 | 0.548 | 7.081 | |
| Flavones | 5,6,7-Tetrahydroxy-8-methoxyflavone | C16H12O6 | 1.037 | 0.416 | 0.548 | 7.076 | |
| Isoflavones | 5,7,4′-Trihydroxy-3′-methoxyisoflavone; 3′-O-methylorobol | C16H12O6 | 1.028 | 0.417 | 0.548 | 7.022 | |
| Flavones | Rhamnocitrin (7-methylkaempferol) | C16H12O6 | 1.026 | 0.417 | 0.548 | 7.015 | |
| Flavones | Chrysoeriol-7-O-gentiobioside | C28H32O16 | 1.147 | 0.405 | 0.548 | 6.910 | |
| Anthocyanidins | Peonidin-3-O-(6′-O-caffeoyl)glucoside | C31H29O14+ | 1.039 | 0.411 | 0.548 | 6.664 | |
| Flavones | Chrysoeriol-7,4′-di-O-glucoside | C28H32O16 | 1.049 | 0.412 | 0.548 | 6.643 | |
| Other Flavonoids | 9,11-dimethoxy-2h-[1,3]dioxolo [4,5-b]xanthen-10-one | C16H12O6 | 1.066 | 0.413 | 0.548 | 6.631 | |
| Flavones | Gnetifolin B | C16H12O6 | 1.084 | 0.412 | 0.548 | 6.583 | |
| Isoflavones | Aracarpene 2 | C16H12O6 | 1.075 | 0.412 | 0.548 | 6.560 | |
| Flavones | 5,7,2′-Trihydroxy-8-methoxyflavone | C16H12O6 | 1.080 | 0.412 | 0.548 | 6.553 | |
| Flavones | Diosmetin (5,7,3′-Trihydroxy-4′-methoxyflavone) | C16H12O6 | 1.060 | 0.413 | 0.548 | 6.553 | |
| Other Flavonoids | 3-(3,4-dihydroxybenzyl)-5,7-dihydroxy-6-methoxychroman-4-one diglucoside | C29H36O17 | 1.015 | 0.410 | 0.548 | 6.465 | |
| Flavones | 5,7,3′,4′-Tetrahydroxy-6-methoxyflavone-8-C-[glucosyl-(1-2)]-glucoside | C28H32O17 | 1.018 | 0.410 | 0.548 | 6.204 | |
| Flavonols | Patuletin-3-O-gentiobioside | C28H32O18 | 1.097 | 0.397 | 0.548 | 5.734 | |
| Flavones | Gardenin B 5-(6′-Malonyl)glucoside | C28H30O15 | 1.430 | 0.100 | 0.542 | 5.393 | |
| Flavonols | Herbacetin | C15H10O7 | 1.385 | 0.244 | 0.548 | 5.178 | |
| Flavonols | Azaleatin (5-O-methylquercetin) | C16H12O7 | 1.476 | 0.113 | 0.542 | 5.142 | |
| Flavones | Tricetin-5-O-(6′-malonyl)glucoside | C24H22O15 | 1.417 | 0.126 | 0.542 | 5.123 | |
| Other Flavonoids | 2′,7-Dihydroxy-3′,4′-dimethoxyisoflavan | C17H18O5 | 1.375 | 0.170 | 0.542 | 5.076 | |
| Flavones | Viscumneoside III (Homoeriodictyol-7-O-apiosyl-(1-2)-glucopyranoside) | C27H32O15 | 1.472 | 0.038 | 0.542 | 5.061 | |
| Flavones | 6-Hydroxyluteolin | C15H10O7 | 1.380 | 0.231 | 0.548 | 4.950 | |
| Lignans and coumarins | Coumarins | Peucedanol | C14H16O5 | 1.381 | 0.318 | 0.548 | 5.637 |
| Lignans | Sanshodiol | C20H22O6 | 1.475 | 0.090 | 0.542 | 5.400 | |
| Lignans | Matairesinol | C20H22O6 | 1.469 | 0.092 | 0.542 | 5.268 | |
| Lignans | Erythro-guaiacylglycerol-β-coniferyl ether | C20H24O7 | 1.449 | 0.150 | 0.542 | 5.119 | |
| Lignans | Lappaol C | C30H34O10 | 1.479 | 0.086 | 0.542 | 4.945 | |
| Coumarins | Esculin glucoside | C21H26O14 | 1.172 | 0.369 | 0.548 | 4.757 | |
| Lignans | Sesamolinol 4′-O-β-D-glucosyl (1→6)-O-β-D-glucoside | C32H40O17 | 1.430 | 0.196 | 0.548 | 4.703 | |
| Phenolic acids | Phenolic acids | 2′-Acetylacteoside | C31H38O16 | 1.293 | 0.310 | 0.548 | 6.452 |
| Phenolic acids | 10-Hydroxymajoroside | C17H24O11 | 1.504 | 0.017 | 0.542 | 5.658 | |
| Phenolic acids | Echinacoside | C35H46O20 | 1.356 | 0.230 | 0.548 | 5.642 | |
| Phenolic acids | 6-O-Feruloyl-β-D-glucose | C16H20O9 | 1.458 | 0.124 | 0.542 | 5.583 | |
| Phenolic acids | Syringoyl-D-glucose | C15H20O10 | 1.208 | 0.185 | 0.542 | 5.137 | |
| Phenolic acids | 1′-O-(3,4-Dihydroxyphenethyl)-O-caffeoyl-glucoside | C23H26O11 | 1.498 | 0.083 | 0.542 | 5.047 | |
| Phenolic acids | Arillatose B | C22H30O14 | 1.482 | 0.040 | 0.542 | 5.010 | |
| Phenolic acids | 5-O-β-D-Glucopyranosyl-3-hydrobenzo(b)fu-ran-2-one | C14H16O8 | 1.486 | 0.040 | 0.542 | 4.954 | |
| Phenolic acids | Rosmarinic acid methyl ester | C19H18O8 | 1.500 | 0.043 | 0.542 | 4.927 | |
| Phenolic acids | Isoferulic acid | C10H10O4 | 1.394 | 0.178 | 0.542 | 4.860 | |
| Phenolic acids | Purpureaside C | C35H46O20 | 1.312 | 0.255 | 0.548 | 4.842 | |
| Phenolic acids | 1,7-bis(4-hydroxy-3-methoxyphenyl)hept-1-ene-3-ol | C21H26O5 | 1.491 | 0.067 | 0.542 | 4.716 | |
| Phenolic acids | Tetrahydrorengyoxide-glucose-caffeoyl | C23H26O11 | 1.472 | 0.085 | 0.542 | 4.712 | |
| Phenolic acids | 3,4,5-Trihydroxy-6-[4-[[(2R)-5-oxooxolan-2-yl]methyl]phenoxy]oxane-2-carboxylic acid | C17H20O9 | 1.495 | 0.052 | 0.542 | 4.667 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dossou, S.S.K.; Luo, Z.; Deng, Q.; Zhou, R.; Zhang, Y.; Li, D.; Li, H.; Tozo, K.; You, J.; Wang, L. Biochemical and Molecular Insights into Variation in Sesame Seed Antioxidant Capability as Revealed by Metabolomics and Transcriptomics Analysis. Antioxidants 2024, 13, 514. https://doi.org/10.3390/antiox13050514
Dossou SSK, Luo Z, Deng Q, Zhou R, Zhang Y, Li D, Li H, Tozo K, You J, Wang L. Biochemical and Molecular Insights into Variation in Sesame Seed Antioxidant Capability as Revealed by Metabolomics and Transcriptomics Analysis. Antioxidants. 2024; 13(5):514. https://doi.org/10.3390/antiox13050514
Chicago/Turabian StyleDossou, Senouwa Segla Koffi, Zishu Luo, Qianchun Deng, Rong Zhou, Yanxin Zhang, Donghua Li, Huan Li, Koffi Tozo, Jun You, and Linhai Wang. 2024. "Biochemical and Molecular Insights into Variation in Sesame Seed Antioxidant Capability as Revealed by Metabolomics and Transcriptomics Analysis" Antioxidants 13, no. 5: 514. https://doi.org/10.3390/antiox13050514
APA StyleDossou, S. S. K., Luo, Z., Deng, Q., Zhou, R., Zhang, Y., Li, D., Li, H., Tozo, K., You, J., & Wang, L. (2024). Biochemical and Molecular Insights into Variation in Sesame Seed Antioxidant Capability as Revealed by Metabolomics and Transcriptomics Analysis. Antioxidants, 13(5), 514. https://doi.org/10.3390/antiox13050514