
In Vivo-Matured Oocyte Resists Post-Ovulatory Aging through the Hub Genes DDX18 and DNAJC7 in Pigs
Abstract
:1. Introduction
2. Methods
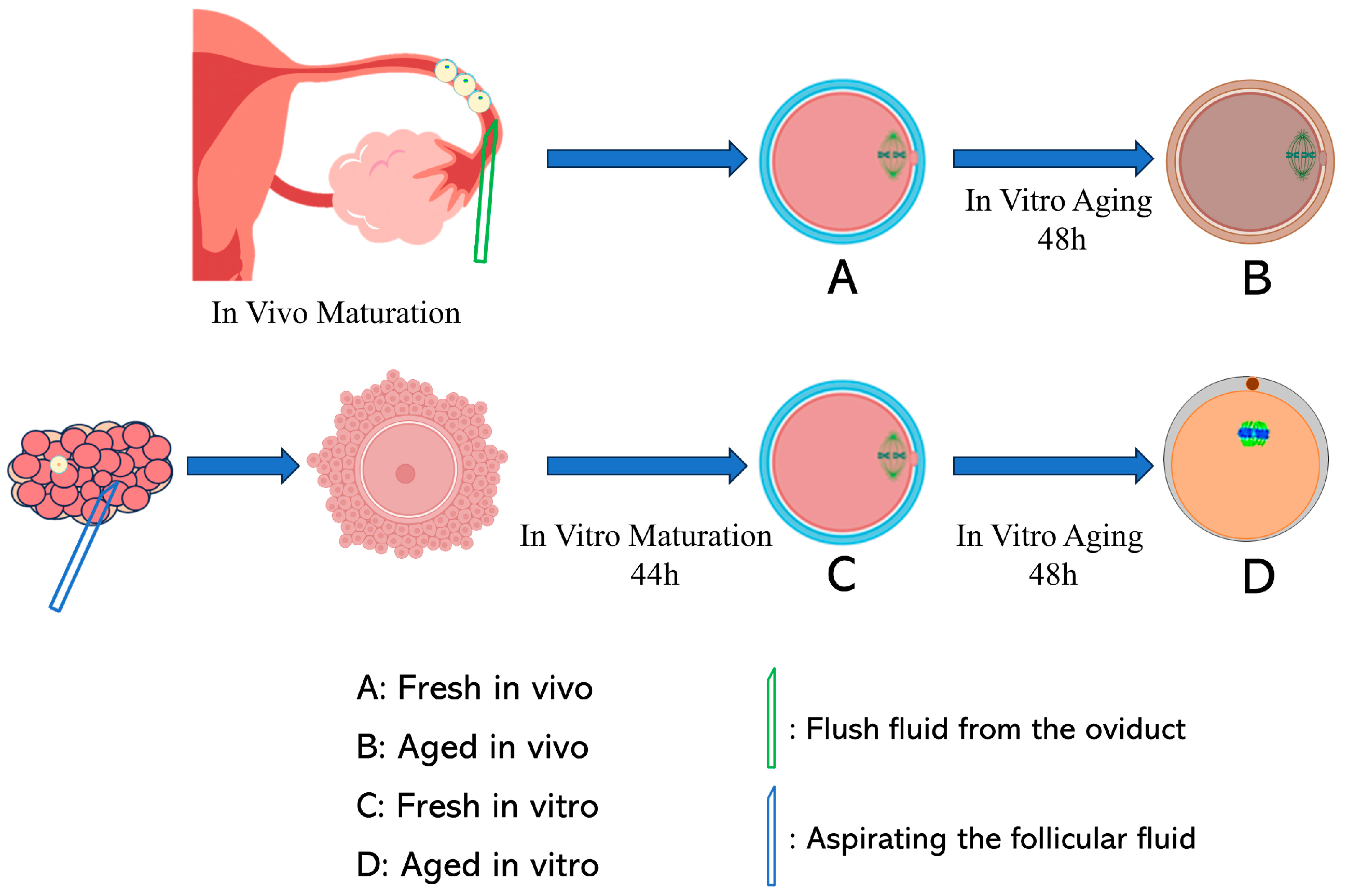
2.1. Collection of Porcine Cumulus–Oocyte Complexes (COCs) and In Vitro Maturation (IVM)
2.2. In Vitro Fresh and Aged Oocyte Collection
2.3. In Vivo—Fresh and Aged—Oocyte Collection
2.4. RNA Extraction
2.5. Quantitative Reverse Transcription PCR (qRT-PCR)
2.6. Construction of cDNA Libraries and RNA Sequencing
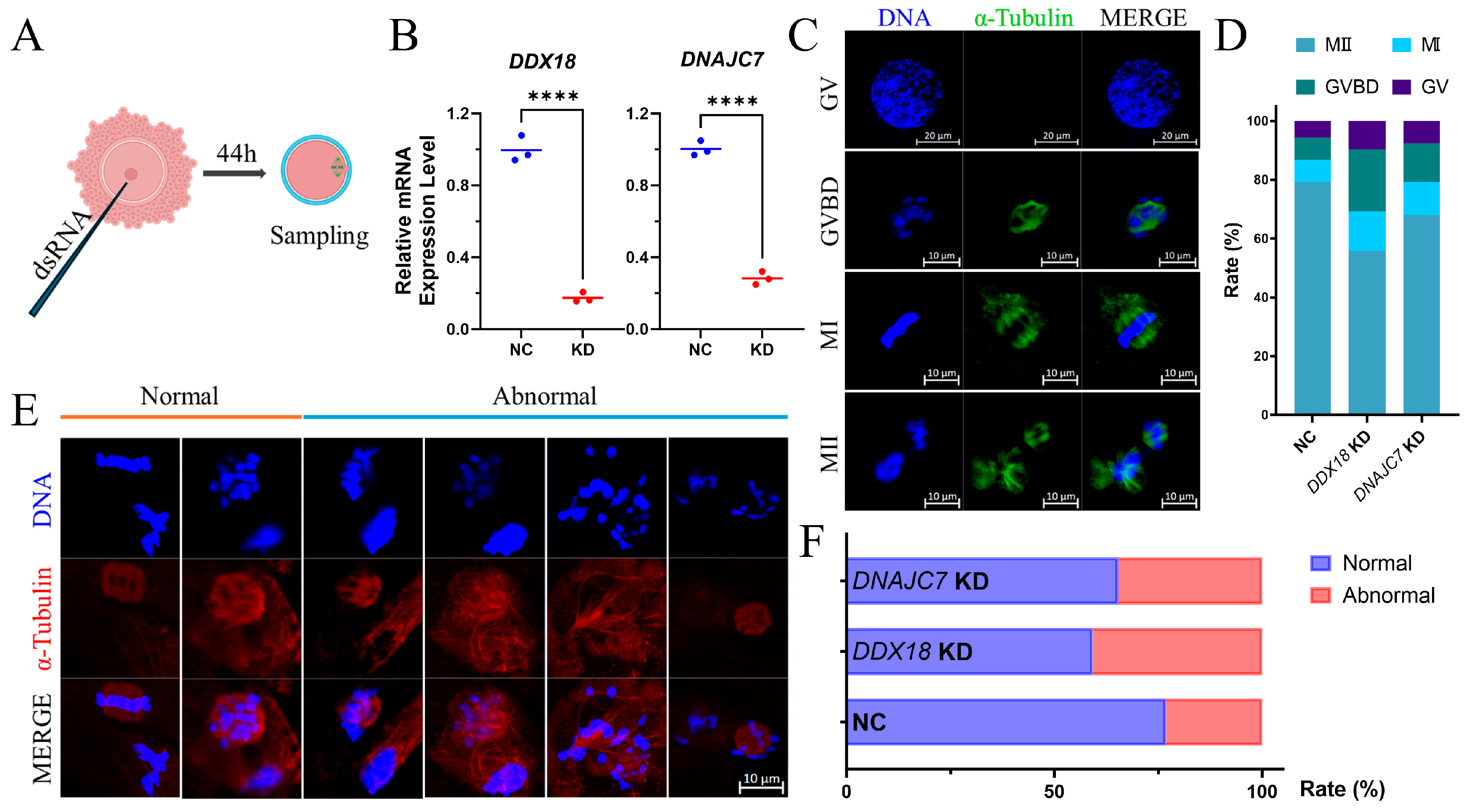
2.7. Preparation of DDX18 and DNAJC7 Double-Stranded RNA (dsRNA)
2.8. dsRNA Injection
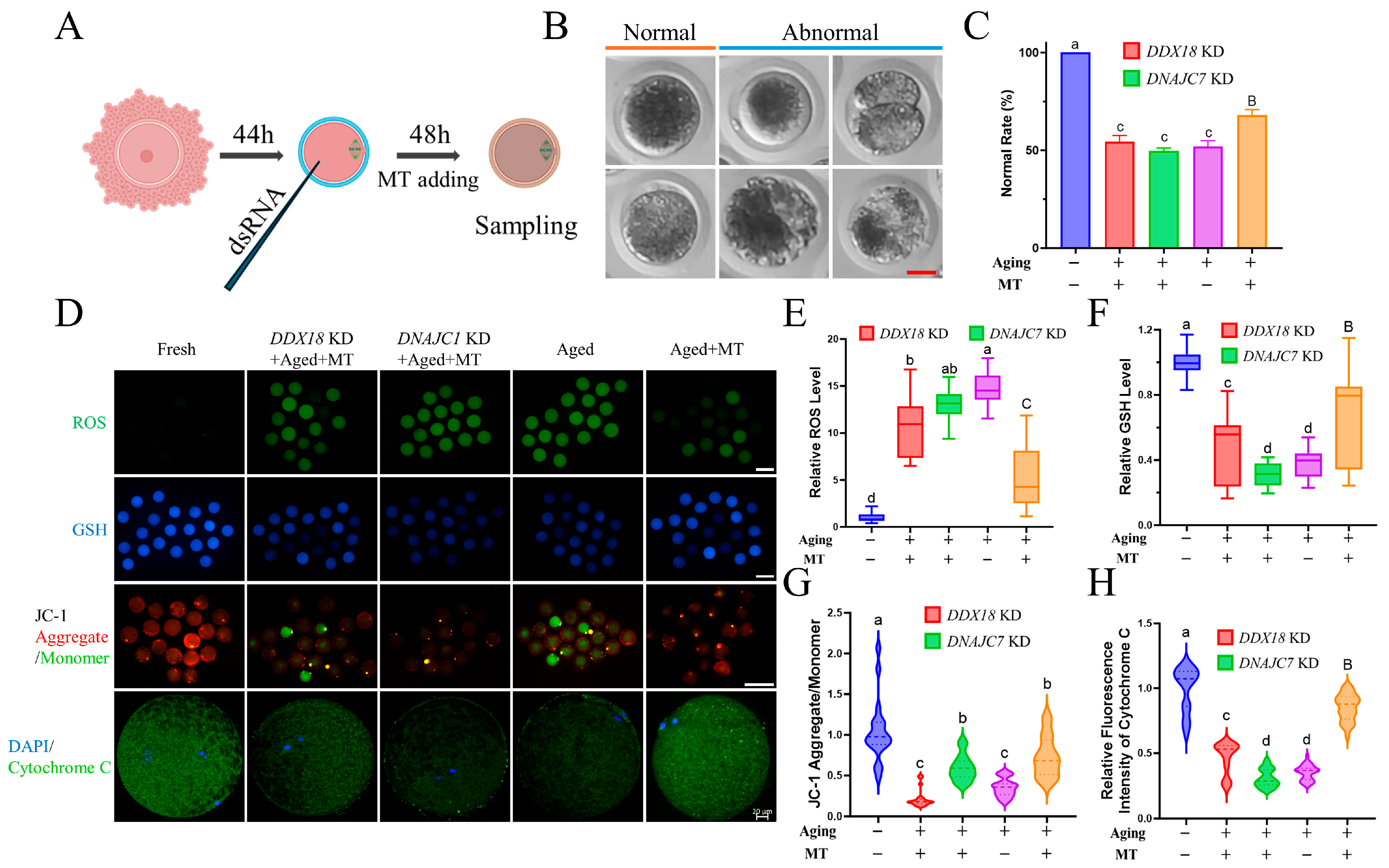
2.9. Measurement of Mitochondrial Membrane Potential (ΔΨm), Reactive Oxygen Species (ROS), and Glutathione (GSH)
2.10. Immunofluorescence Staining
2.11. Statistical Analysis
3. Results
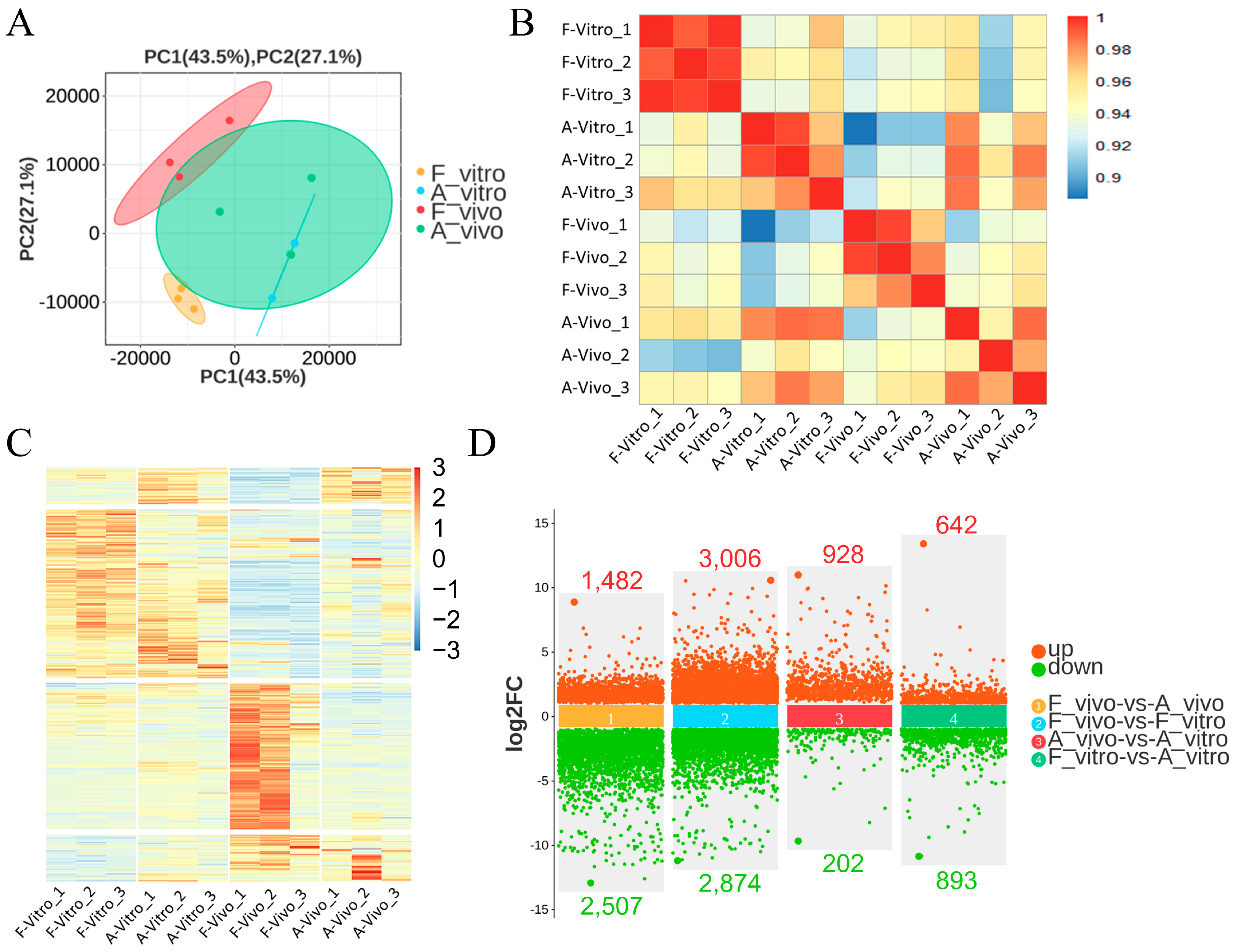
3.1. Global Transcriptome Analysis of Fresh and Aged In Vivo and Vitro-Matured Porcine Oocytes
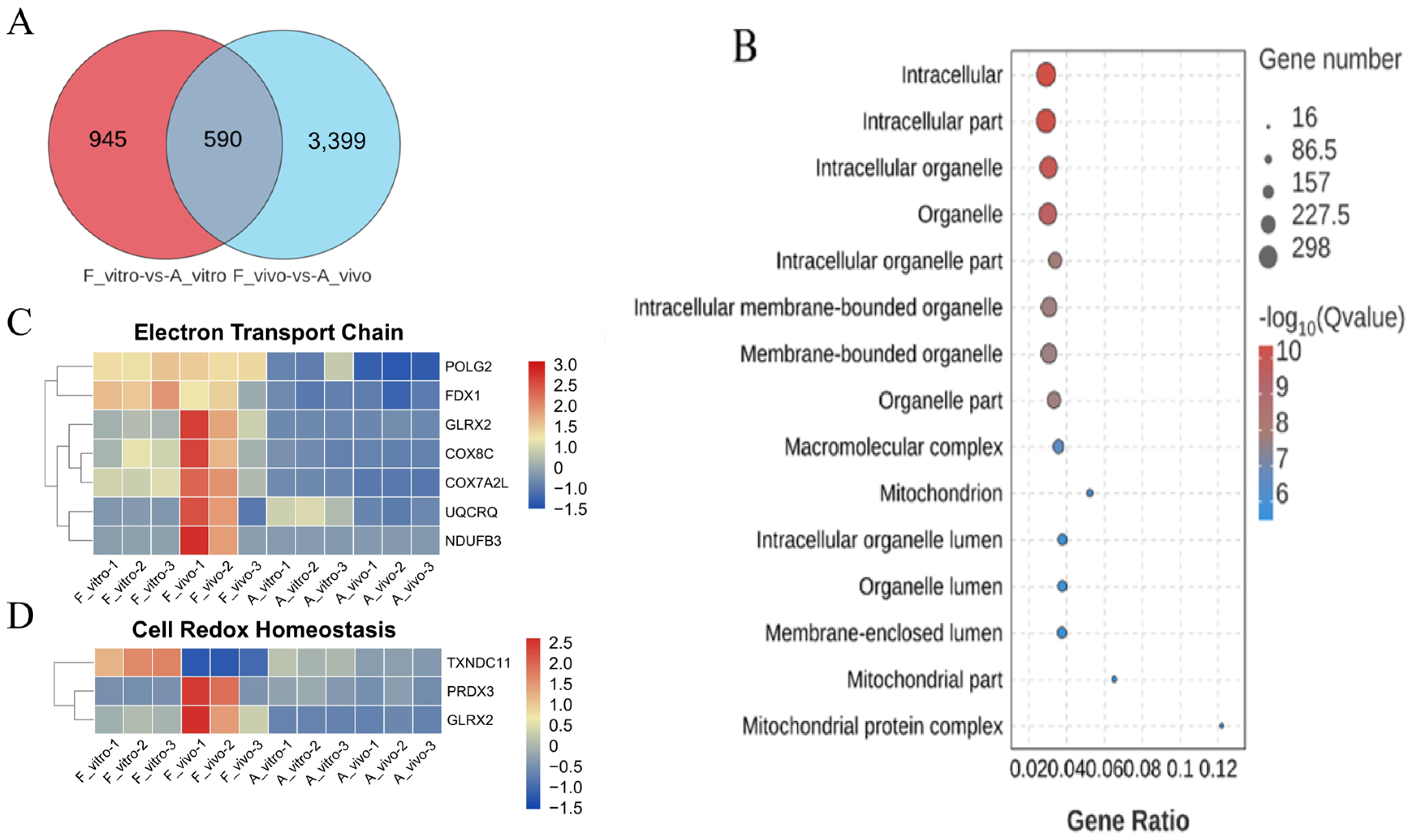
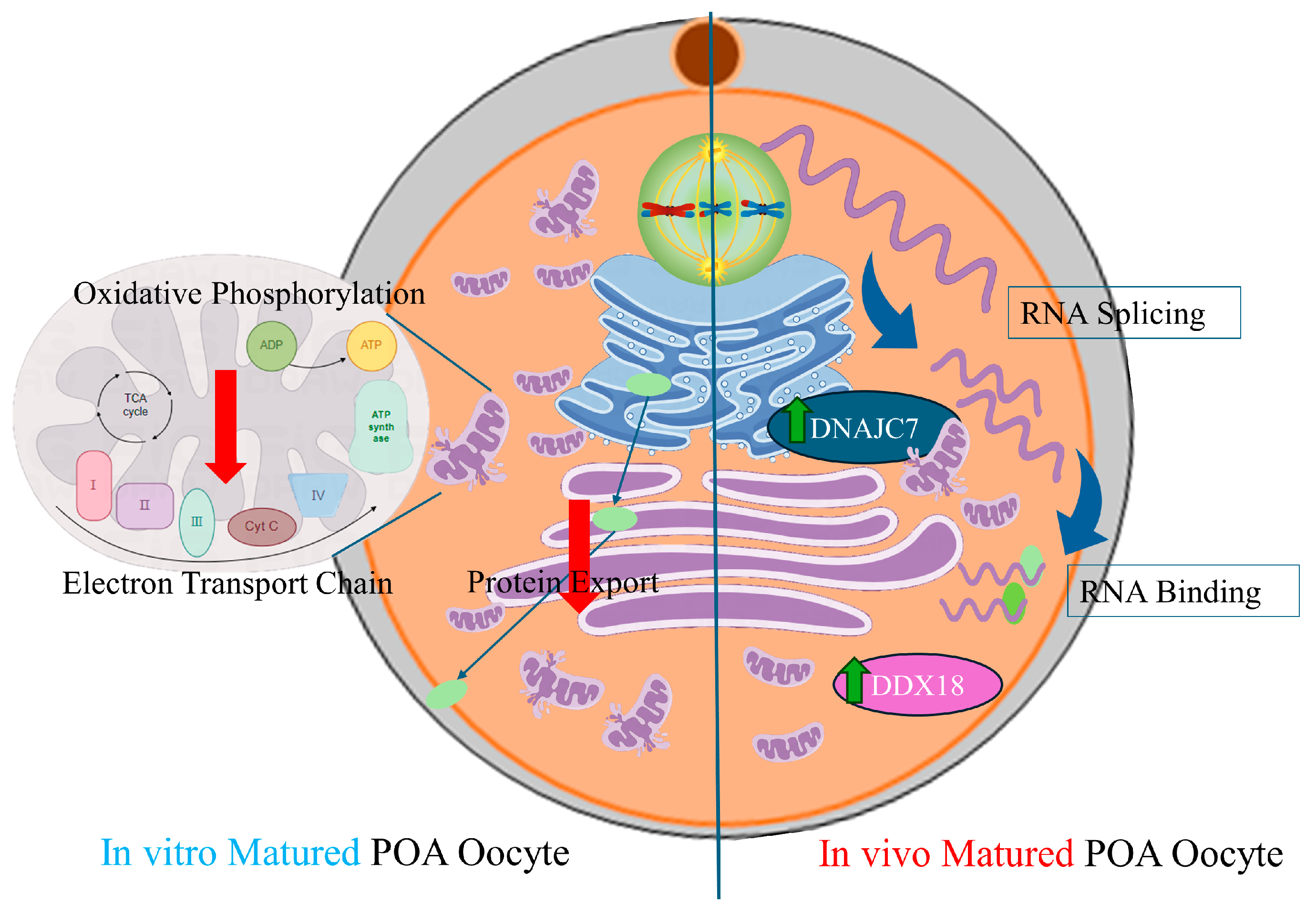
3.2. POA Interferes with the Mitochondrial Function in Both In Vivo- and Vitro-Matured Porcine Oocytes
3.3. POA Affects Protein Export and Oocyte Meiosis in Both In Vivo- and Vitro-Matured Porcine Oocytes
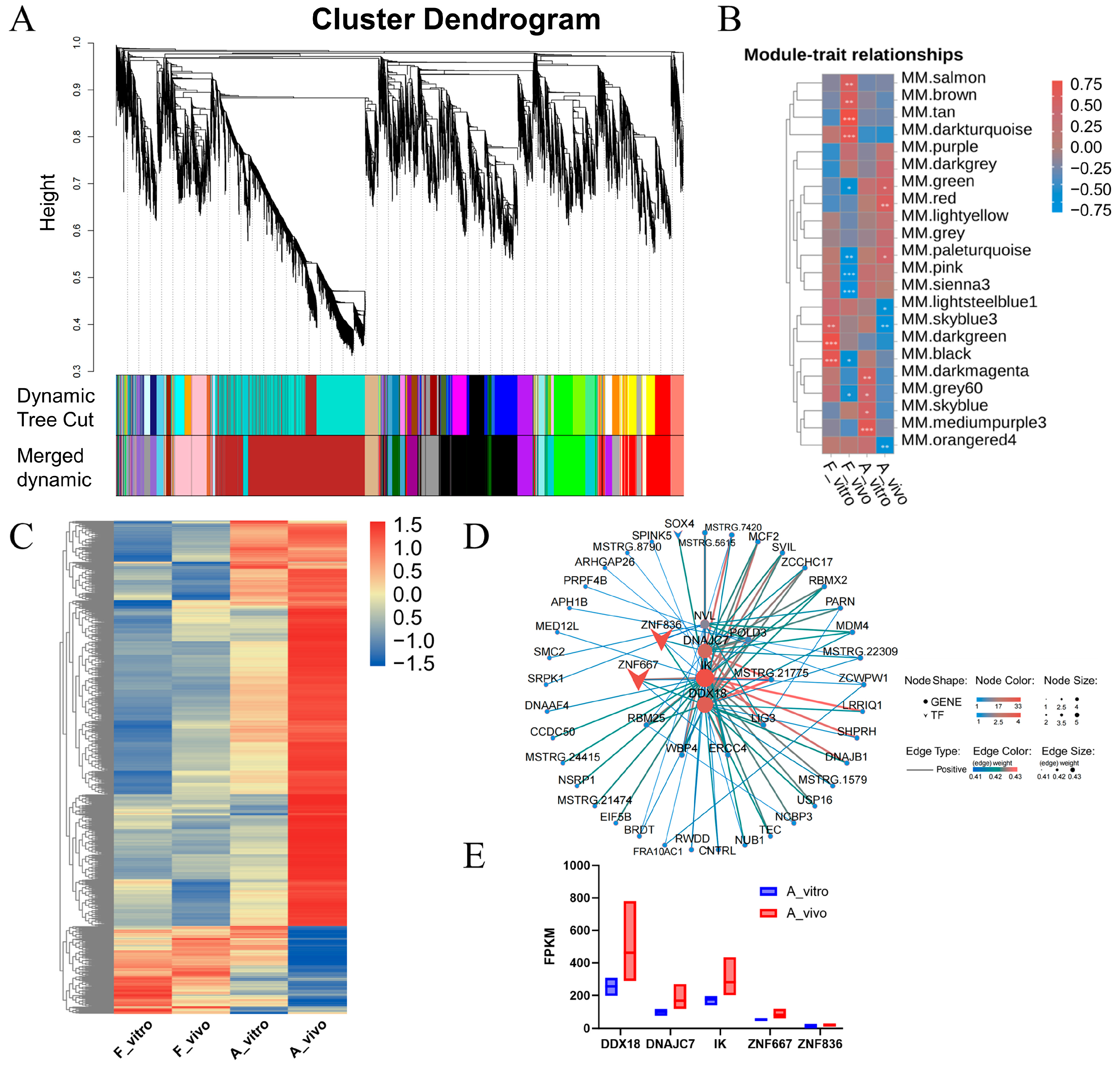
3.4. DNAJC7, DDX18 and IK Were the Hub Genes in the Gene Regulation Network in Aged In Vivo Porcine Oocytes
3.5. Knockdown of DNAJC7 and DDX18 at GV Stage Resulted in Cell Cycle Disorders and Abnormal Spindle Morphology in Porcine Oocytes
3.6. Knockdown of DNAJC7 and DDX18 at MII Stage Resulted in Oxidative Stress and Mitochondrial Dysfunction in Porcine Oocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lord, T.; Aitken, R.J. Oxidative stress and ageing of the post-ovulatory oocyte. Reproduction 2013, 146, R217–R227. [Google Scholar] [CrossRef] [PubMed]
- Di Nisio, V.; Antonouli, S.; Damdimopoulou, P.; Salumets, A.; Cecconi, S. In vivo and in vitro postovulatory aging: When time works against oocyte quality? J. Assist. Reprod. Genet. 2022, 39, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; ShiYang, X.; Zhang, Y.; Miao, Y.; Chen, Y.; Cui, Z.; Xiong, B. Coenzyme Q10 ameliorates the quality of postovulatory aged oocytes by suppressing DNA damage and apoptosis. Free Radic. Biol. Med. 2019, 143, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Abbott, A.; Kopf, G.S.; Schultz, R.M.; Ducibella, T. Spontaneous activation of ovulated mouse eggs: Time-dependent effects on M-phase exit, cortical granule exocytosis, maternal messenger ribonucleic acid recruitment, and inositol 1,4,5-trisphosphate sensitivity. Biol. Reprod. 1997, 57, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Ducibella, T.; Duffy, P.; Reindollar, R.; Su, B. Changes in the distribution of mouse oocyte cortical granules and ability to undergo the cortical reaction during gonadotropin-stimulated meiotic maturation and aging in vivo. Biol. Reprod. 1990, 43, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Wakayama, S.; Thuan, N.V.; Kishigami, S.; Ohta, H.; Mizutani, E.; Hikichi, T.; Miyake, M.; Wakayama, T. Production of offspring from one-day-old oocytes stored at room temperature. J. Reprod. Dev. 2004, 50, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Naito, K.; Noguchi, J.; Kaneko, H.; Tojo, H. Maturation/M-phase promoting factor regulates aging of porcine oocytes matured in vitro. Cloning Stem Cells 2002, 4, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Lord, T.; Nixon, B.; Jones, K.T.; Aitken, R.J. Melatonin prevents postovulatory oocyte aging in the mouse and extends the window for optimal fertilization in vitro. Biol. Reprod. 2013, 88, 67. [Google Scholar] [CrossRef]
- Zhang, N.; Wakai, T.; Fissore, R.A. Caffeine alleviates the deterioration of Ca(2+) release mechanisms and fragmentation of in vitro-aged mouse eggs. Mol. Reprod. Dev. 2011, 78, 684–701. [Google Scholar] [CrossRef]
- Tatone, C.; Di Emidio, G.; Barbaro, R.; Vento, M.; Ciriminna, R.; Artini, P.G. Effects of reproductive aging and postovulatory aging on the maintenance of biological competence after oocyte vitrification: Insights from the mouse model. Theriogenology 2011, 76, 864–873. [Google Scholar] [CrossRef]
- Guérin, P.; El Mouatassim, S.; Ménézo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.W.; Zhu, J.Q.; Miao, Y.L.; Liu, J.H.; Wei, L.; Lu, S.S.; Hou, Y.; Schatten, H.; Lu, K.H.; Sun, Q.Y. Loss of methylation imprint of Snrpn in postovulatory aging mouse oocyte. Biochem. Biophys. Res. Commun. 2008, 371, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Z.P.; Rienzi, L.F.; Ubaldi, F.M.; Greco, E.; Massey, J.B.; Kort, H.I. Effect of reduced oocyte aging on the outcome of rescue intracytoplasmic sperm injection. Fertil. Steril. 2006, 85, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sekhon, L.; Kim, Y.; Agarwal, A. The role of oxidative stress and antioxidants in assisted reproduction. Curr. Womens Health Rev. 2010, 6, 227–238. [Google Scholar] [CrossRef]
- Reis Soares, S.; Rubio, C.; Rodrigo, L.; Simón, C.; Remohí, J.; Pellicer, A. High frequency of chromosomal abnormalities in embryos obtained from oocyte donation cycles. Fertil. Steril. 2003, 80, 656–657. [Google Scholar] [CrossRef] [PubMed]
- Babayev, E.; Seli, E. Oocyte mitochondrial function and reproduction. Curr. Opin. Obstet. Gynecol. 2015, 27, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A. Usefulness of bovine and porcine IVM/IVF models for reproductive toxicology. Reprod. Biol. Endocrinol. 2014, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, Y.; Yang, J.Y.; Cheng, D.; Liu, X.; Ma, X.; West, F.D.; Wang, H. Comparative gene expression signature of pig, human and mouse induced pluripotent stem cell lines reveals insight into pig pluripotency gene networks. Stem Cell Rev. Rep. 2014, 10, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Jarrell, V.L.; Day, B.N.; Prather, R.S. The transition from maternal to zygotic control of development occurs during the 4-cell stage in the domestic pig, Sus scrofa: Quantitative and qualitative aspects of protein synthesis. Biol. Reprod. 1991, 44, 62–68. [Google Scholar] [CrossRef]
- Haggarty, P.; Wood, M.; Ferguson, E.; Hoad, G.; Srikantharajah, A.; Milne, E.; Hamilton, M.; Bhattacharya, S. Fatty acid metabolism in human preimplantation embryos. Hum. Reprod. 2006, 21, 766–773. [Google Scholar] [CrossRef]
- McEvoy, T.G.; Coull, G.D.; Broadbent, P.J.; Hutchinson, J.S.; Speake, B.K. Fatty acid composition of lipids in immature cattle, pig and sheep oocytes with intact zona pellucida. J. Reprod. Fertil. 2000, 118, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Sun, M.H.; Jiang, W.J.; Li, X.H.; Lee, S.H.; Heo, G.; Niu, Y.J.; Ock, S.A.; Cui, X.S. Epigallocatechin-3-gallate protects porcine oocytes against post-ovulatory aging through inhibition of oxidative stress. Aging 2022, 14, 8633–8644. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.J.; Zhou, W.; Nie, Z.W.; Zhou, D.; Xu, Y.N.; Ock, S.A.; Yan, C.G.; Cui, X.S. Ubiquinol-10 delays postovulatory oocyte aging by improving mitochondrial renewal in pigs. Aging 2020, 12, 1256–1271. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.Y.; Sa, S.J.; Jeong, Y.D.; Cho, E.S.; Hong, J.G.; YS, K.I.; Cho, K.H.; Park, S.H.; Kim, K.W.; Lee, H.C.; et al. Altrenogest affects expression of galectin-3 and fibroblast growth factor 9 in the reproductive tract of sows. Anim. Biotechnol. 2021, 32, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Min, C.G.; Zhang, K.Y.; Zhan, C.L.; Wang, Y.C.; Hou, S.K.; Ma, X.; Lu, W.F. Tetrabromobisphenol Exposure Impairs Bovine Oocyte Maturation by Inducing Mitochondrial Dysfunction. Molecules 2022, 27, 8111. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Xu, Y.; Zhang, X.; Shi, X.; Zhang, Y.; Luo, M.; Wu, C.; Ding, Z.; Xiang, H.; Cao, Y. Melatonin mitigates PNMC-induced disruption of spindle assembly and mitochondrial function in mouse Oocytes. Ecotoxicol. Environ. Saf. 2024, 282, 116703. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Cao, X.; Zhang, T.; Guo, J.; Xu, G.; Wang, H.; Yang, W.; Yang, L.; Che, D.; Lu, W.; et al. Melatonin protects porcine oocyte from copper exposure potentially by reducing oxidative stress potentially through the Nrf2 pathway. Theriogenology 2022, 193, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, X.Q.; Liu, R.P.; Li, Y.H.; Yao, X.R.; Kim, N.H.; Xu, Y.N. Melatonin Supplementation during In Vitro Maturation of Porcine Oocytes Alleviates Oxidative Stress and Endoplasmic Reticulum Stress Induced by Imidacloprid Exposure. Animals 2023, 13, 2596. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Li, X.H.; Lu, Q.Y.; Zhan, C.L.; Kim, J.D.; Lee, G.H.; Sim, J.M.; Cui, X.S. Nobiletin enhances mitochondrial function by regulating SIRT1/PGC-1α signaling in porcine oocytes during in vitro maturation. Biochem. Biophys. Res. Commun. 2024, 706, 149747. [Google Scholar] [CrossRef]
- Grøndahl, M.L.; Yding Andersen, C.; Bogstad, J.; Nielsen, F.C.; Meinertz, H.; Borup, R. Gene expression profiles of single human mature oocytes in relation to age. Hum. Reprod. 2010, 25, 957–968. [Google Scholar] [CrossRef]
- Reyes, J.M.; Silva, E.; Chitwood, J.L.; Schoolcraft, W.B.; Krisher, R.L.; Ross, P.J. Differing molecular response of young and advanced maternal age human oocytes to IVM. Hum. Reprod. 2017, 32, 2199–2208. [Google Scholar] [CrossRef] [PubMed]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef] [PubMed]
- Van Blerkom, J.; Davis, P.W.; Lee, J. ATP content of human oocytes and developmental potential and outcome after in-vitro fertilization and embryo transfer. Hum. Reprod. 1995, 10, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hao, J.X.; Liu, B.W.; Ouyang, Y.C.; Guo, J.N.; Dong, M.Z.; Wang, Z.B.; Gao, F.; Yao, Y.Q. Supplementation of mitochondria from endometrial mesenchymal stem cells improves oocyte quality in aged mice. Cell Prolif. 2023, 56, e13372. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Meng, T.G.; Ouyang, Y.C.; Liu, S.L.; Guo, J.N.; Wang, Z.B.; Schatten, H.; Song, C.Y.; Guo, X.P.; Sun, Q.Y. Miro1 regulates mitochondrial homeostasis and meiotic resumption of mouse oocyte. J. Cell Physiol. 2022, 237, 4477–4486. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, N.; Schlotter, F.; Lefebvre, S.; Dodré, M.; Méreau, A.; Soret, J.; Besse, A.; Barkats, M.; Bordonné, R.; Branlant, C.; et al. Implication of the SMN complex in the biogenesis and steady state level of the signal recognition particle. Nucleic Acids Res. 2013, 41, 1255–1272. [Google Scholar] [CrossRef]
- Evans, E.A.; Gilmore, R.; Blobel, G. Purification of microsomal signal peptidase as a complex. Proc. Natl. Acad. Sci. USA 1986, 83, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, N.; Wu, M.J.; Sousa, C.F.; Kalinina, O.V.; Welsch, C.; Yi, M. SPCS1-Dependent E2-p7 processing determines HCV Assembly efficiency. PLoS Pathog. 2022, 18, e1010310. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Mullins, C.; Green, N. In addition to SEC11, a newly identified gene, SPC3, is essential for signal peptidase activity in the yeast endoplasmic reticulum. J. Biol. Chem. 1997, 272, 13152–13158. [Google Scholar] [CrossRef]
- Böhni, P.C.; Deshaies, R.J.; Schekman, R.W. SEC11 is required for signal peptide processing and yeast cell growth. J. Cell Biol. 1988, 106, 1035–1042. [Google Scholar] [CrossRef]
- García-Aguirre, I.; Alamillo-Iniesta, A.; Rodríguez-Pérez, R.; Vélez-Aguilera, G.; Amaro-Encarnación, E.; Jiménez-Gutiérrez, E.; Vásquez-Limeta, A.; Samuel Laredo-Cisneros, M.; Morales-Lázaro, S.L.; Tiburcio-Félix, R.; et al. Enhanced nuclear protein export in premature aging and rescue of the progeria phenotype by modulation of CRM1 activity. Aging Cell 2019, 18, e13002. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Schieweck, R.; Ninkovic, J.; Kiebler, M.A. RNA-binding proteins balance brain function in health and disease. Physiol. Rev. 2021, 101, 1309–1370. [Google Scholar] [CrossRef] [PubMed]
- Lenzken, S.C.; Achsel, T.; Carrì, M.T.; Barabino, S.M. Neuronal RNA-binding proteins in health and disease. Wiley Interdiscip. Rev. RNA 2014, 5, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, F.; Schwarzl, T.; Valcárcel, J.; Hentze, M.W. RNA-binding proteins in human genetic disease. Nat. Rev. Genet. 2021, 22, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Varesi, A.; Campagnoli, L.I.M.; Barbieri, A.; Rossi, L.; Ricevuti, G.; Esposito, C.; Chirumbolo, S.; Marchesi, N.; Pascale, A. RNA binding proteins in senescence: A potential common linker for age-related diseases? Ageing Res. Rev. 2023, 88, 101958. [Google Scholar] [CrossRef] [PubMed]
- Conlon, E.G.; Manley, J.L. RNA-binding proteins in neurodegeneration: Mechanisms in aggregate. Genes. Dev. 2017, 31, 1509–1528. [Google Scholar] [CrossRef] [PubMed]
- Cookson, M.R. RNA-binding proteins implicated in neurodegenerative diseases. Wiley Interdiscip. Rev. RNA 2017, 8, e1397. [Google Scholar] [CrossRef] [PubMed]
- Salvato, I.; Ricciardi, L.; Dal Col, J.; Nigro, A.; Giurato, G.; Memoli, D.; Sellitto, A.; Lamparelli, E.P.; Crescenzi, M.A.; Vitale, M.; et al. Expression of targets of the RNA-binding protein AUF-1 in human airway epithelium indicates its role in cellular senescence and inflammation. Front. Immunol. 2023, 14, 1192028. [Google Scholar] [CrossRef]
- Yoon, D.S.; Lee, K.M.; Choi, Y.; Ko, E.A.; Lee, N.H.; Cho, S.; Park, K.H.; Lee, J.H.; Kim, H.W.; Lee, J.W. TLR4 downregulation by the RNA-binding protein PUM1 alleviates cellular aging and osteoarthritis. Cell Death Differ. 2022, 29, 1364–1378. [Google Scholar] [CrossRef]
- Kwon, S.M.; Min, S.; Jeoun, U.W.; Sim, M.S.; Jung, G.H.; Hong, S.M.; Jee, B.A.; Woo, H.G.; Lee, C.; Yoon, G. Global spliceosome activity regulates entry into cellular senescence. FASEB J. 2021, 35, e21204. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, M.; Zhang, P.; Hou, E.; Li, T.; Liu, X.; Xu, X.; Wang, Z.; Fan, Y.; Zhen, X.; et al. Aberrant spliceosome expression and altered alternative splicing events correlate with maturation deficiency in human oocytes. Cell Cycle 2020, 19, 2182–2194. [Google Scholar] [CrossRef] [PubMed]
- Kubo, N.; Wu, D.; Yoshihara, Y.; Sang, M.; Nakagawara, A.; Ozaki, T. Co-chaperon DnaJC7/TPR2 enhances p53 stability and activity through blocking the complex formation between p53 and MDM2. Biochem. Biophys. Res. Commun. 2013, 430, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.L.; Chen, J.K.; Wen, X.; He, W.; Zarceno, G.A.; Chen, Y.; Chen, S.; Paull, T.T.; Liu, H.W. DDX18 prevents R-loop-induced DNA damage and genome instability via PARP-1. Cell Rep. 2022, 40, 111089. [Google Scholar] [CrossRef] [PubMed]
- Ka, H.I.; Seo, H.; Choi, Y.; Kim, J.; Cho, M.; Choi, S.Y.; Park, S.; Han, S.; An, J.; Chung, H.S.; et al. Loss of splicing factor IK impairs normal skeletal muscle development. BMC Biol. 2021, 19, 44. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Lu, Y.; Zhang, M.; Miao, Y.; Zhou, C.; Cui, Z.; Xiong, B. Melatonin improves the fertilization ability of post-ovulatory aged mouse oocytes by stabilizing ovastacin and Juno to promote sperm binding and fusion. Hum. Reprod. 2017, 32, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Full Name | Primer Sequence (5′~3′) | Gene ID |
|---|---|---|---|
| BTF3 | Basic transcription factor 3 | F: GTGTGTGCGCCTTATCTCAG R: GTTTGGCGAGTTTCTCCTGG | 100514824 |
| CEP95 | Centrosomal protein 95 | F: AGAGGGCAGGAGAGAGGTTA R: ACATCCTCCTCTTCACAGCC | 100624364 |
| CMC1 | C-X9-C motif containing 1 | F: CGCAGAACAGCATCTCAGAC R: TCCAGAGTCCTTGCAGCATT | 102164646 |
| DBI | Diazepam binding inhibitor | F: ACAGCCACTACAAACAAGCG R: ACGCTTTCATGGCATCTTCC | 397212 |
| NDUFB5 | NADH: ubiquinone oxidoreductase subunit B5 | F: GCTTTGCCCTCAGTCAACAT R: CATGGCTACTATGGGCGAGA | 100523751 |
| DDX18 | DEAD-box helicase 18 | F: GCCAACACGCAGACAGACCAT R: CCTGCTCAAGACCATCCACTGT | 100153560 |
| * F: TGCGACAGTGGATGGTCTTGAG * R: TGCTTCCTGGGCTGACTTATGA | |||
| DNAJC7 | DnaJ heat shock protein family (Hsp40) member C7 | F: CCTGCCTGCCATCGCTTCAA R: AGCATCTGCGTTGGTGGAATCC | 100524895 |
| * F: GGATTCCACCAACGCAGATGCT * R: TCCTTCTGAACTTCCGCACTGG | |||
| 18s | 18S ribosomal RNA | F: CGCGGTTCTATTTTGTTGGT R: AGTCGGCATCGTTTATGGTC | 100861538 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, C.-L.; Zhou, D.; Sun, M.-H.; Jiang, W.-J.; Lee, S.-H.; Li, X.-H.; Lu, Q.-Y.; Kim, J.-D.; Lee, G.-H.; Sim, J.-M.; et al. In Vivo-Matured Oocyte Resists Post-Ovulatory Aging through the Hub Genes DDX18 and DNAJC7 in Pigs. Antioxidants 2024, 13, 867. https://doi.org/10.3390/antiox13070867
Zhan C-L, Zhou D, Sun M-H, Jiang W-J, Lee S-H, Li X-H, Lu Q-Y, Kim J-D, Lee G-H, Sim J-M, et al. In Vivo-Matured Oocyte Resists Post-Ovulatory Aging through the Hub Genes DDX18 and DNAJC7 in Pigs. Antioxidants. 2024; 13(7):867. https://doi.org/10.3390/antiox13070867
Chicago/Turabian StyleZhan, Cheng-Lin, Dongjie Zhou, Ming-Hong Sun, Wen-Jie Jiang, Song-Hee Lee, Xiao-Han Li, Qin-Yue Lu, Ji-Dam Kim, Gyu-Hyun Lee, Jae-Min Sim, and et al. 2024. "In Vivo-Matured Oocyte Resists Post-Ovulatory Aging through the Hub Genes DDX18 and DNAJC7 in Pigs" Antioxidants 13, no. 7: 867. https://doi.org/10.3390/antiox13070867