Dietary Supplementation of Yerba Mate (Ilex paraguariensis) during the Dry Period Improves Redox Balance in Lactating Dairy Cows


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Location, Husbandry, and Experimental Protocol
2.2. Blood Sampling and Assays
2.3. Statistical Analysis
3. Results
3.1. Redox Balance
3.2. Body Condition Score and Liveweight
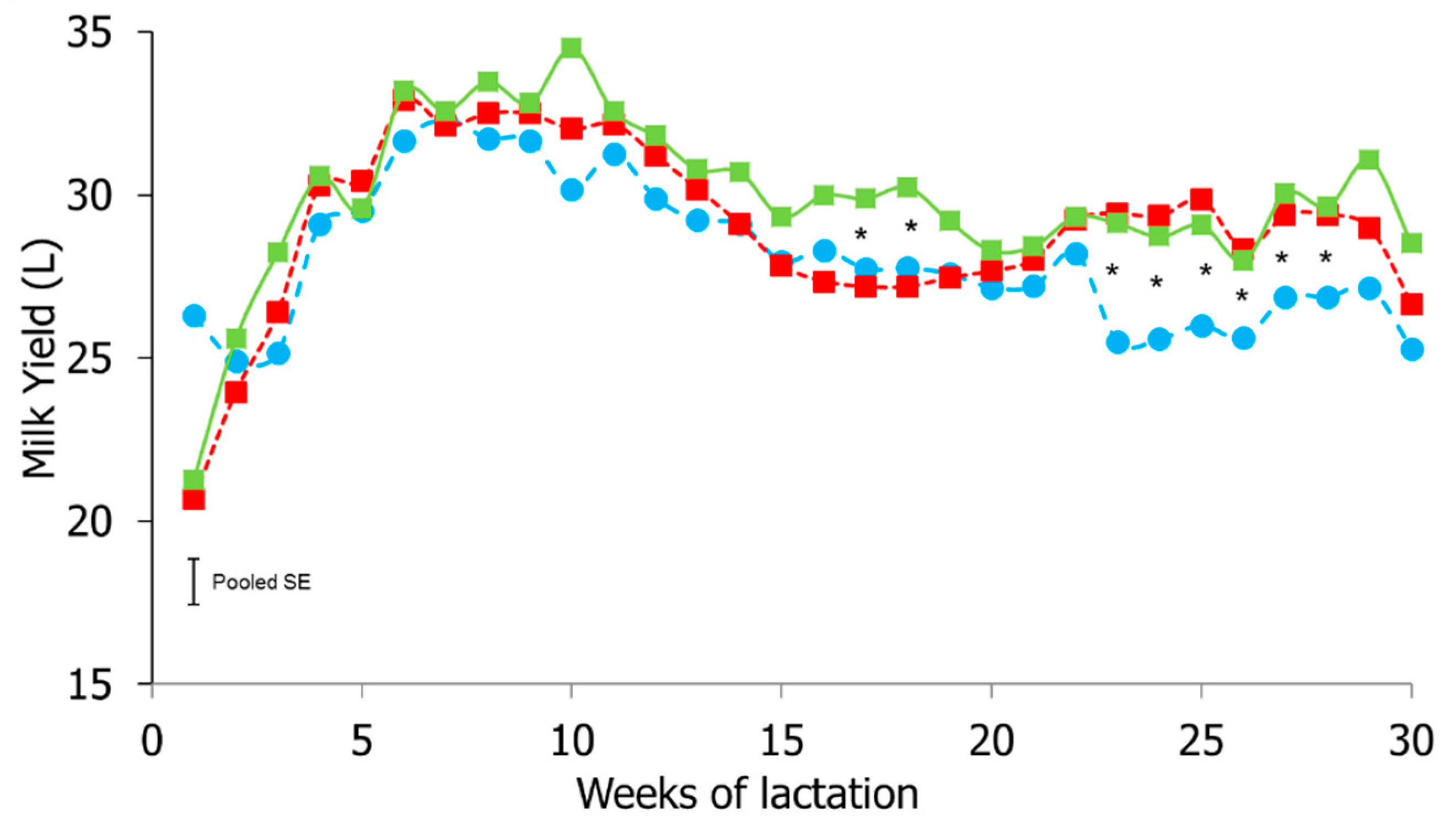
3.3. Milk Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Drackley, J.K. Biology of Dairy Cows during the Transition Period: the Final Frontier. J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Bernabucci, U.; Ronchi, B.; Lacetera, N.; Nardone, A. Influence of body condition score on relationships between metabolic status and oxidative stress in periparturient dairy cows. J. Dairy Sci. 2005, 88, 2017–2026. [Google Scholar] [CrossRef]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. The importance of the oxidative status of dairy cattle in the periparturient period: revisiting antioxidant supplementation. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.; Duval, S.M.; Tamassia, L.F.M.; Kindermann, M.; Stemmler, R.T.; de Gouvea, V.N.; Acedo, T.S.; Immig, I.; Williams, S.N.; Celi, P. Nutritional strategies in ruminants: A lifetime approach. Res. Vet. Sci. 2018, 116, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Celi, P. Oxidative Stress in Ruminants. In Studies on Veterinary Medicine; Mandelker, L., Vajdovich, P., Eds.; Humana Press: New York, NY, USA, 2011; Volume 5, pp. 191–231. [Google Scholar]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.K.; Brzezinska-Slebodzinska, E.; Madsen, F.C. Oxidative stress, antioxidants, and animal function. J. Dairy Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- DeGaris, P.J.; Lean, I.J.; Rabiee, A.R.; Heuer, C. Effects of increasing days of exposure to prepartum transition diets on reproduction and health in dairy cows. Aust. Vet. J. 2010, 88, 84–92. [Google Scholar] [CrossRef]
- Chagas, L.M.; Bass, J.J.; Blache, D.; Burke, C.R.; Kay, J.K.; Lindsay, D.R.; Lucy, M.C.; Martin, G.B.; Meier, S.; Rhodes, F.M.; et al. Invited Review: New Perspectives on the Roles of Nutrition and Metabolic Priorities in the Subfertility of High-Producing Dairy Cows. J. Dairy Sci. 2007, 90, 4022–4032. [Google Scholar] [CrossRef]
- Chandra, S.; De Mejia Gonzalez, E. Polyphenolic compounds, antioxidant capacity, and quinone reductase activity of an aqueous extract of Ardisia compressa in comparison to mate (Ilex paraguariensis) and green (Camellia sinensis) teas. J. Agric. Food Chem. 2004, 52, 3583–3589. [Google Scholar] [CrossRef]
- Filip, R.; Lotito, S.B.; Ferraro, G.; Fraga, C.G. Antioxidant activity of Ilex paraguariensis and related species. Nutr. Res. 2000, 20, 1437–1446. [Google Scholar] [CrossRef]
- Schinella, G.R.; Troiani, G.; Dávila, V.; de Buschiazzo, P.M.; Tournier, H.A. Antioxidant Effects of an Aqueous Extract of Ilex paraguariensis. Biochem. Biophys. Res. Commun. 2000, 269, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Heck, C.I.; de Mejia, E.G. Yerba Mate Tea (Ilex paraguariensis): A Comprehensive Review on Chemistry, Health Implications, and Technological Considerations. J. Food Sci. 2007, 72, R138–R151. [Google Scholar] [CrossRef] [PubMed]
- Po, E.; Horsburgh, K.; Raadsma, H.W.; Celi, P. Yerba Mate (Ilex paraguarensis) as a novel feed supplement for growing lambs. Small Ruminant Res. 2012, 106, 131–136. [Google Scholar] [CrossRef]
- Po, E.; Xu, Z.; Celi, P. The Effect of Yerba Mate (Ilex paraguarensis) Supplementation on the Productive Performance of Dorper Ewes and Their Progeny. Asian Australas. J. Anim. Sci. 2012, 25, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Celi, P.; Raadsma, H.W. Effects of Yerba Mate (Ilex paraguariensis) supplementation on the productive performance of dairy cows during mid-lactation. Anim. Prod. Sci. 2010, 50, 339–344. [Google Scholar] [CrossRef]
- Celi, P.; Robinson, A. Effect of Yerba Mate (Ilex paraguariensis) supplementation on the performance of dairy calves. Anim. Prod. Sci. 2010, 50, 376–381. [Google Scholar] [CrossRef]
- Santos, F.S.; Zeoula, L.M.; dos Santos, G.T.; Lima, L.S.; Dias, A.L.G.; Rufino, M.O.A.; Schogor, A.L.B.; De Marchi, F.E.; Petit, H.V. Intake, digestibility and milk production and composition of dairy cows fed different levels of Yerba Mate in the diet. Anim. Feed Sci. Tech. 2017, 230, 70–76. [Google Scholar] [CrossRef]
- Edmonson, A.J.; Lean, I.J.; Weaver, L.D.; Farver, T.; Webster, G. A Body Condition Scoring Chart for Holstein Dairy Cows. J. Dairy Sci. 1989, 72, 68–78. [Google Scholar] [CrossRef]
- Cesarone, M.R.; Belcaro, G.; Carratelli, M.; Cornelli, U.; De Sanctis, M.T.; Incandela, L.; Barsotti, A.; Terranova, R.; Nicolaides, A. A simple test to monitor oxidative stress. Int. Angiol. 1999, 18, 127–130. [Google Scholar] [PubMed]
- Alberti, A.; Bolognini, L.; Macciantelli, D.; Caratelli, M. The Radical Cation of N,N-Diethyl-para-Phenylendiamine: A Possible Indicator of Oxidative Stress in Biological Samples. Res. Chem. Intermediat. 2000, 26, 253–267. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Khoa, T.N.; Capeillere-Blandin, C.; Nguyen, A.T.; Canteloup, S.; Dayer, J.-M.; Jungers, P.; Drueke, T.; Descamps-Latscha, B. Advanced Oxidation Protein Products as Novel Mediators of Inflammation and Monocyte Activation in Chronic Renal Failure. J. Immunol. 1998, 161, 2524–2532. [Google Scholar] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239. [Google Scholar] [CrossRef] [PubMed]
- Celi, P. Biomarkers of oxidative stress in ruminant medicine. Immunopharmacol. Immunotoxicol. 2011, 33, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. Oxidative stress index (OSi) as a new tool to assess redox status in dairy cattle during the transition period. Animal 2013, 7, 1374–1378. [Google Scholar] [CrossRef] [PubMed]
- Bernabucci, U.; Ronchi, B.; Lacetera, N.; Nardone, A. Markers of oxidative status in plasma and erythrocytes of transition dairy cows during hot season. J. Dairy Sci. 2002, 85, 2173–2179. [Google Scholar] [CrossRef]
- Di Trana, A.; Celi, P.; Claps, S.; Fedele, V.; Rubino, R. The effect of hot season and nutrition on the oxidative status and metabolic profile in dairy goats during mid lactation. Anim. Sci. 2006, 82, 717–722. [Google Scholar] [CrossRef]
- Celi, P.; Gabai, G. Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Front Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef]
- Gabai, G.; Testoni, S.; Piccinini, R.; Marinelli, L.; Stradaioli, G. Oxidative stress in primiparous cows in relation to dietary starch and the progress of lactation. Animal Sci. 2004, 79, 99–108. [Google Scholar]
- Pedernera, M.; Celi, P.; García, S.C.; Salvin, H.E.; Barchia, I.; Fulkerson, W.J. Effect of diet, energy balance and milk production on oxidative stress in early-lactating dairy cows grazing pasture. Vet. J. 2010, 186, 352–357. [Google Scholar] [CrossRef]
- Celi, P.; Merlo, M.; Da Dalt, L.; Stefani, A.; Barbato, O.; Gabai, G. Relationship between late embryonic mortality and the increase in plasma advanced oxidised protein products (AOPP) in dairy cows. Reprod Fert Dev 2011, 23, 527–533. [Google Scholar] [CrossRef]
- Celi, P.; Merlo, M.; Barbato, O.; Gabai, G. Relationship between oxidative stress and the success of artificial insemination in dairy cows in a pasture-based system. Vet. J. 2012, 193, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Golder, H.M.; Lean, I.J.; Rabiee, A.R.; King, R.; Celi, P. Effects of grain, fructose, and histidine feeding on endotoxin and oxidative stress measures in dairy heifers. J. Dairy Sci. 2013, 96, 7881–7891. [Google Scholar] [CrossRef] [PubMed]
- Fialovà, L.; Malbohan, I.; Kalousova, M.; Soukupova, J.; Krofta, L.; Štipek, S.; Zima, T. Oxidative stress and inflammation in pregnancy. Scand. J. Clin. Lab. Invest. 2006, 66, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Noyan, T.; Güler, A.; Sekeroglu, M.R.; Kamaci, M. Serum advanced oxidation protein products, myeloperoxidase and ascorbic acid in pre-eclampsia and eclampsia. Aust. N. Z. J. Obstet. Gynaecol. 2006, 46, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Bordignon, M.; Da Dalt, L.; Marinelli, L.; Gabai, G. Advanced oxidation protein products are generated by bovine neutrophils and inhibit free radical production in vitro. Vet. J. 2014, 199, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Ballet, N.; Robert, J.C.; Williams, P.E.V. Vitamins in forages. In Forage Evaluation in Ruminant Nutrition; Givens, D.I., Owen, E., Axford, R.F.E., Omed, H.M., Eds.; CABI Publishing: Oxon, UK, 2000; Vol. Chapter 19, pp. 399–431. [Google Scholar]
- Cattani, M.; Tagliapietra, F.; Bailoni, L.; Schiavon, S. Synthetic and natural polyphenols with antioxidant properties stimulate rumen microbial growth in vitro. Anim. Prod. Sci. 2012, 52, 44–50. [Google Scholar] [CrossRef]
- Lean, I.J.; Lucy, M.C.; McNamara, J.P.; Bradford, B.J.; Block, E.; Thomson, J.M.; Morton, J.M.; Celi, P.; Rabiee, A.R.; Santos, J.E.P.; et al. Invited review: Recommendations for reporting intervention studies on reproductive performance in dairy cattle: Improving design, analysis, and interpretation of research on reproduction. J. Dairy Sci. 2016, 99, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Talukder, S.; Kerrisk, K.L.; Gabai, G.; Celi, P. Role of oxidant–antioxidant balance in reproduction of domestic animals. Anim. Prod. Sci. 2017, 57, 1588–1597. [Google Scholar] [CrossRef]
- Vázquez-Añón, M.; Nocek, J.; Bowman, G.; Hampton, T.; Atwell, C.; Vazquez, P.; Jenkins, T. Effects of Feeding a Dietary Antioxidant in Diets with Oxidized Fat on Lactation Performance and Antioxidant Status of the Cow. J. Dairy Sci. 2008, 91, 3165–3172. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Sheffield, L.G.; Saylor, D. Impact of ethoxyquin on productivity of dairy cattle. J. Dairy Sci. 2002, 85, 358. [Google Scholar]
- Brioukhanov, A.L.; Netrusov, A.I. Catalase and superoxide dismutase: Distribution, properties and physiological role in cells of strict anaerobes. Biochem 2004, 69, 949–962. [Google Scholar] [CrossRef]
- Hartemink, E.; Giorgio, D.; Kaur, R.; Di Trana, A.; Celi, P. The Effect of Yerba Mate (Ilex Paraguariensis) Supplementation on Nutrient Degradability in Dairy Cows: An In sacco and In vitro Study. Asian-Australas J. Anim. Sci. 2015, 28, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Bracesco, N.; Sanchez, A.G.; Contreras, V.; Menini, T.; Gugliucci, A. Recent advances on Ilex paraguariensis research: Minireview. J. Ethnopharmacol. 2011, 136, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Lean, I.J.; DeGaris, P.J.; Celi, P.; McNeill, D.M.; Rodney, R.M.; Fraser, D.R. Influencing the future: Interactions of skeleton, energy, protein and calcium during late gestation and early lactation. Anim. Prod. Sci. 2014, 54, 1177–1189. [Google Scholar] [CrossRef]
- Li, S.; Hacker, R.R. The effect of caffeine on mammary gland development and milk yield in primiparous sows. J. Anim. Sci. 1995, 73, 534–540. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Item | Diet | Months from Calving | SE | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| −1 | 0 | 1 | 3 | 4 | Diet | Time | DxT | |||
| ROMs | Control | 105.1 | 116.6 | 133.2 | 130.1 | 121.4 | 3.7 | *** | ||
| YM 250 | 104.2 | 119.6 | 122.1 | 121.9 | 126.6 | 2.9 | ||||
| YM 500 | 111.1 | 108.6 | 110.3 | 112.6 | 112.1 | 2.5 | ||||
| BAP | Control | 2560.9 | 2643.3 | 2254.2 | 1954.8 | 2556.4 | 95.6 | *** | ||
| YM 250 | 2671.4 | 3197.4 | 3152.4 | 2543.9 | 2883.4 | 95.9 | ||||
| YM 500 | 2775.1 | 2958.3 | 2600.4 | 2485.9 | 3037.5 | 77.5 | ||||
| OSI | Control | 4.96 | 4.41 | 5.91 a | 6.65 a | 4.75 | 0.4 | *** | * | |
| YM 250 | 3.73 | 3.74 | 3.87 b | 4.79 b | 4.39 | 0.1 | ||||
| YM 500 | 4.01 | 3.67 | 4.23 b | 4.53 b | 3.69 | 0.1 | ||||
| AOPP | Control | 26.4 | 16.3 | 46.4 | 52.9 | 51.9 | 5.5 | *** | ||
| YM 250 | 27.3 | 24.9 | 45.5 | 57.9 | 48.8 | 4.8 | ||||
| YM 500 | 26.4 | 24.4 | 42.8 | 61.9 | 46.6 | 5.2 | ||||
| Item | Diet | Months from Calving | SE | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| −1 | 0 | 1 | 3 | 4 | 5 | 6 | Diet | Time | DxT | |||
| BCS | Control | 3.4 | 3.2 | 2.9 | 2.7 | 2.5 | 2.7 | 3.1 | 0.11 | *** | ||
| YM 250 | 3.4 | 3.3 | 2.8 | 2.8 | 2.7 | 2.7 | 2.9 | 0.10 | ||||
| YM 500 | 3.1 | 3.3 | 3.0 | 2.7 | 2.6 | 2.8 | 3.2 | 0.09 | ||||
| LW | Control | 687 | 693 | 624 | 575 | 600 | 602 | 595 | 16 | *** | ||
| YM 250 | 655 | 677 | 616 | 589 | 581 | 569 | 576 | 14 | ||||
| YM 500 | 712 | 690 | 655 | 602 | 586 | 600 | 617 | 16 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbato, O.; Holmes, B.; Filipescu, I.-E.; Celi, P. Dietary Supplementation of Yerba Mate (Ilex paraguariensis) during the Dry Period Improves Redox Balance in Lactating Dairy Cows. Antioxidants 2019, 8, 38. https://doi.org/10.3390/antiox8020038
Barbato O, Holmes B, Filipescu I-E, Celi P. Dietary Supplementation of Yerba Mate (Ilex paraguariensis) during the Dry Period Improves Redox Balance in Lactating Dairy Cows. Antioxidants. 2019; 8(2):38. https://doi.org/10.3390/antiox8020038
Chicago/Turabian StyleBarbato, Olimpia, Belinda Holmes, Iulia-Elena Filipescu, and Pietro Celi. 2019. "Dietary Supplementation of Yerba Mate (Ilex paraguariensis) during the Dry Period Improves Redox Balance in Lactating Dairy Cows" Antioxidants 8, no. 2: 38. https://doi.org/10.3390/antiox8020038
APA StyleBarbato, O., Holmes, B., Filipescu, I.-E., & Celi, P. (2019). Dietary Supplementation of Yerba Mate (Ilex paraguariensis) during the Dry Period Improves Redox Balance in Lactating Dairy Cows. Antioxidants, 8(2), 38. https://doi.org/10.3390/antiox8020038