Sulfuric Odor Precursor S-Allyl-l-Cysteine Sulfoxide in Garlic Induces Detoxifying Enzymes and Prevents Hepatic Injury

 ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
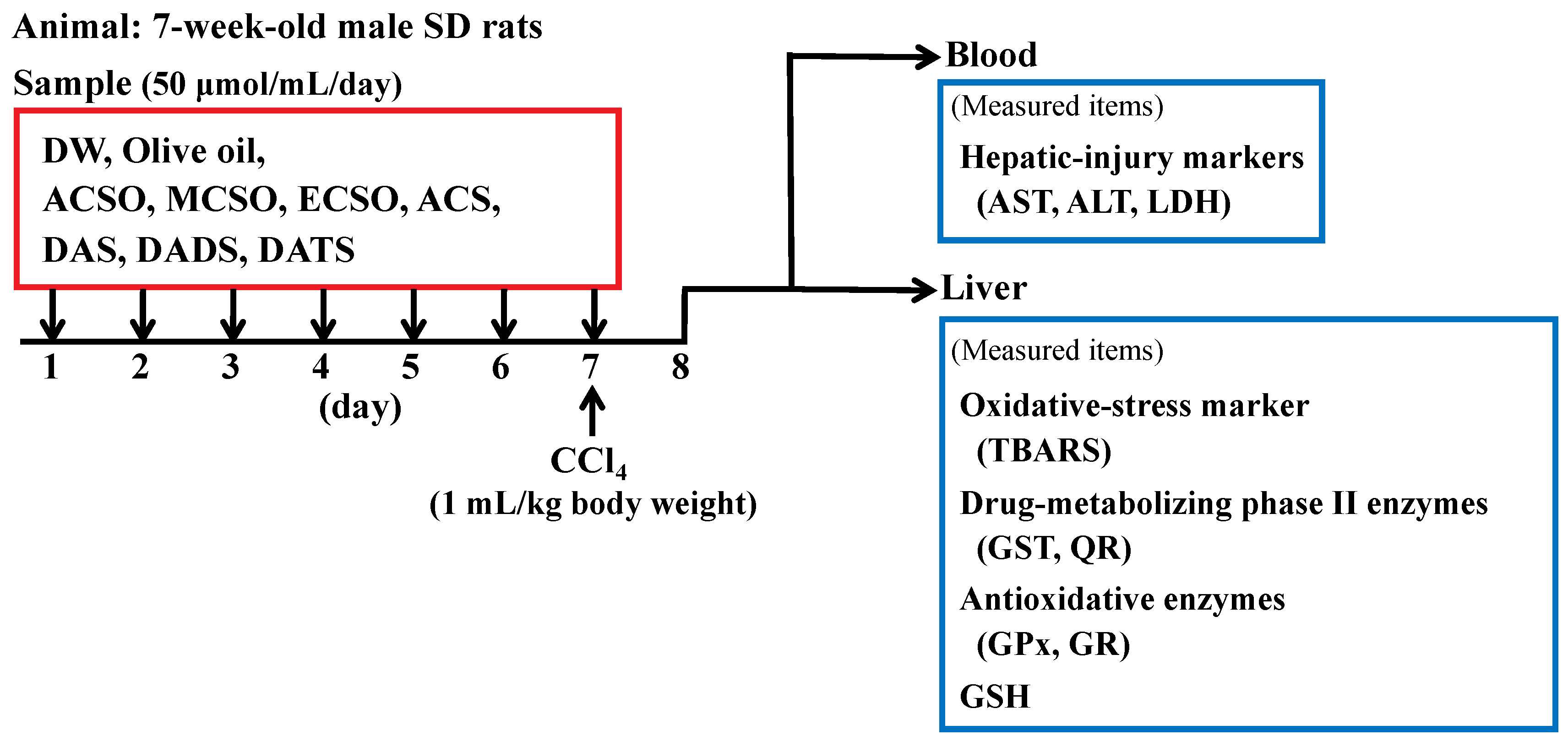
2.2. In Vivo and Ex Vivo Experiments
2.2.1. Animals
2.2.2. Effect of ACSO and Its Related Compounds on Suppression of Hepatic Injury Induced by CCl4
(a) Determination of AST, ALT and LDH Levels
(b) Measurement of Lipid Peroxide
(c) Measurement of GST Activity
(d) Measurement of QR Activity
(e) Measurement of GR Activity
(f) Measurement of GPx Activity
(g) Measurement of total GSH Level
2.2.3. Effect of ACSO and Sulfides on Liver Function of Normal Rats
2.2.4. ACSO Absorption and Metabolism in the Small Intestine
(a) ACSO Absorption from the Small Intestine
(b) Measurement of ACSO and Pyruvic-Acid Content
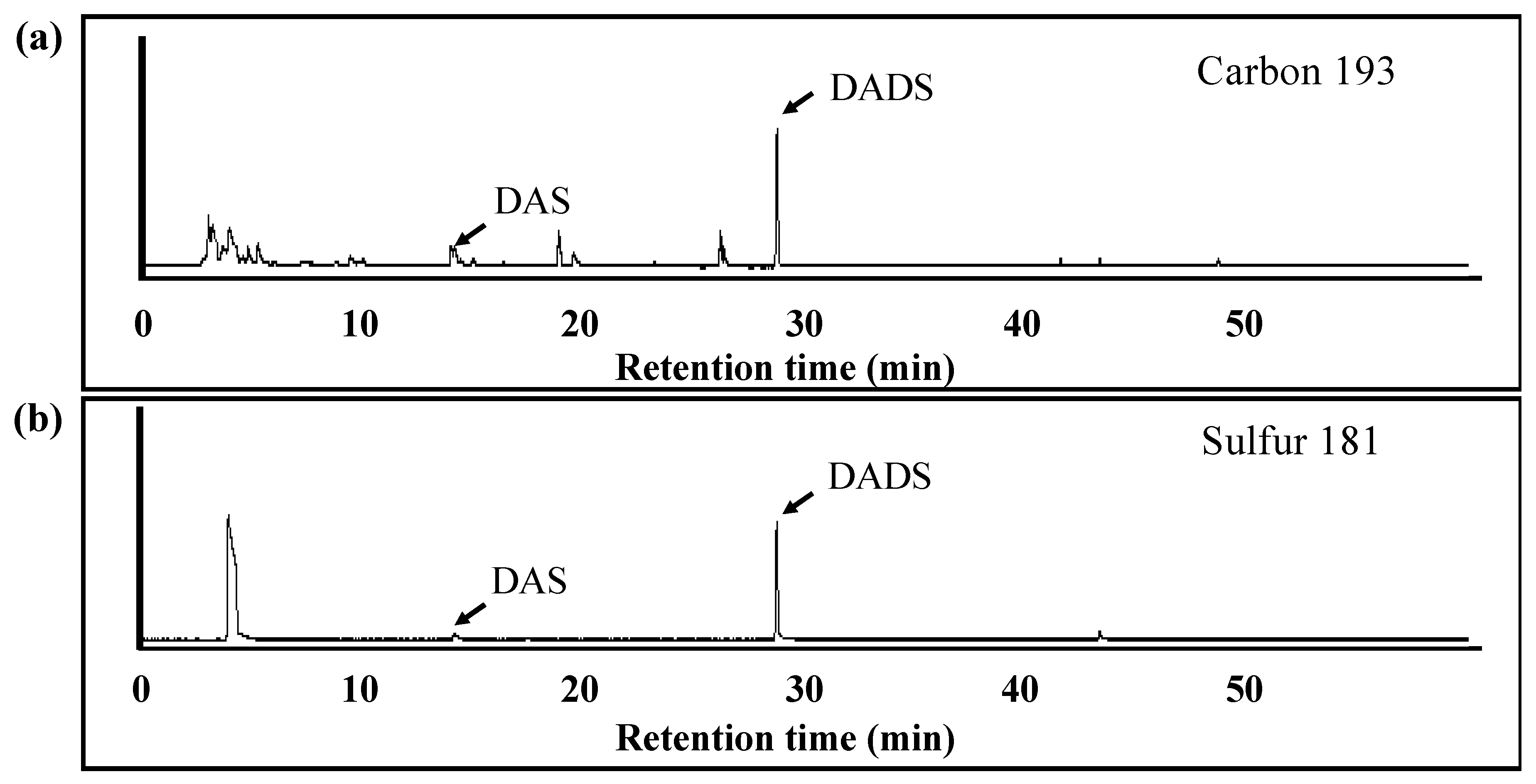
(c) Analysis of Volatile Metabolites from ACSO in the Small Intestine
2.3. In vitro Experiments
2.3.1. Cell Culture
2.3.2. Nrf2 Content in the Nucleus, Cytoplasm, and Whole Cells
2.4. Statistical Analysis
3. Results
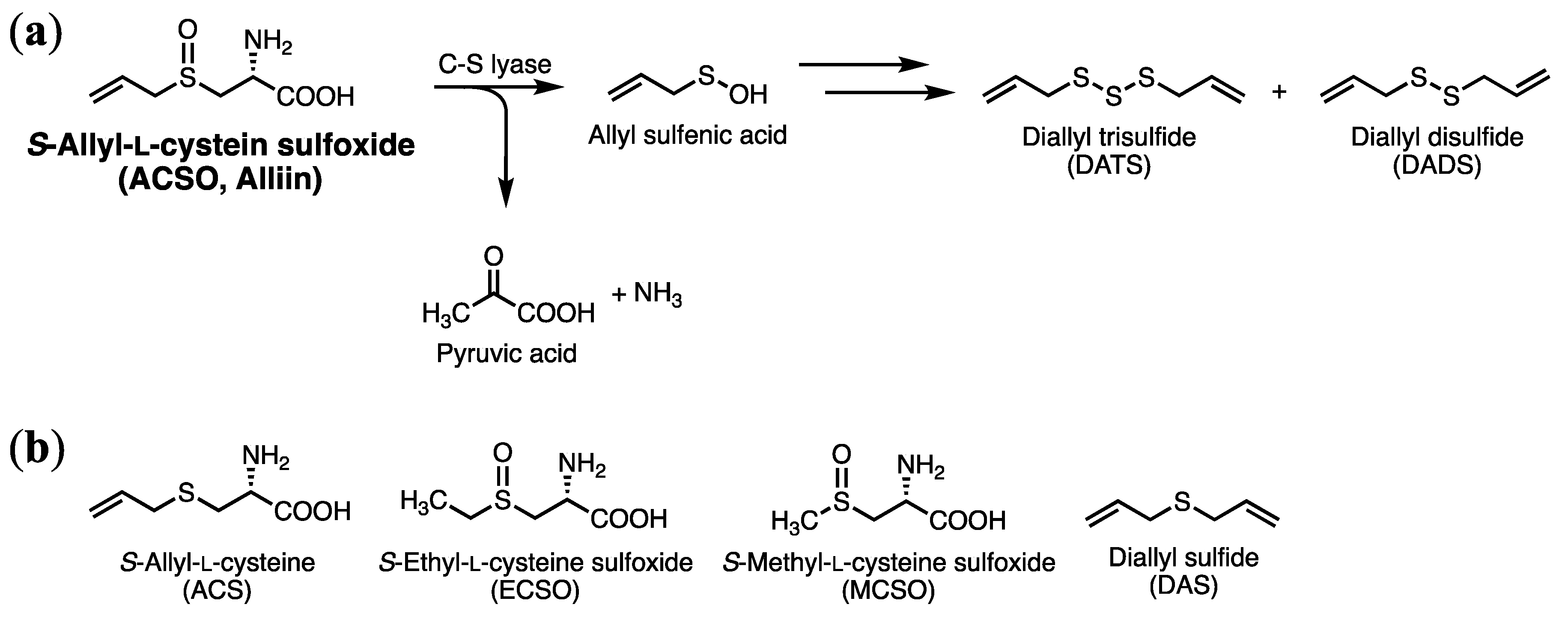
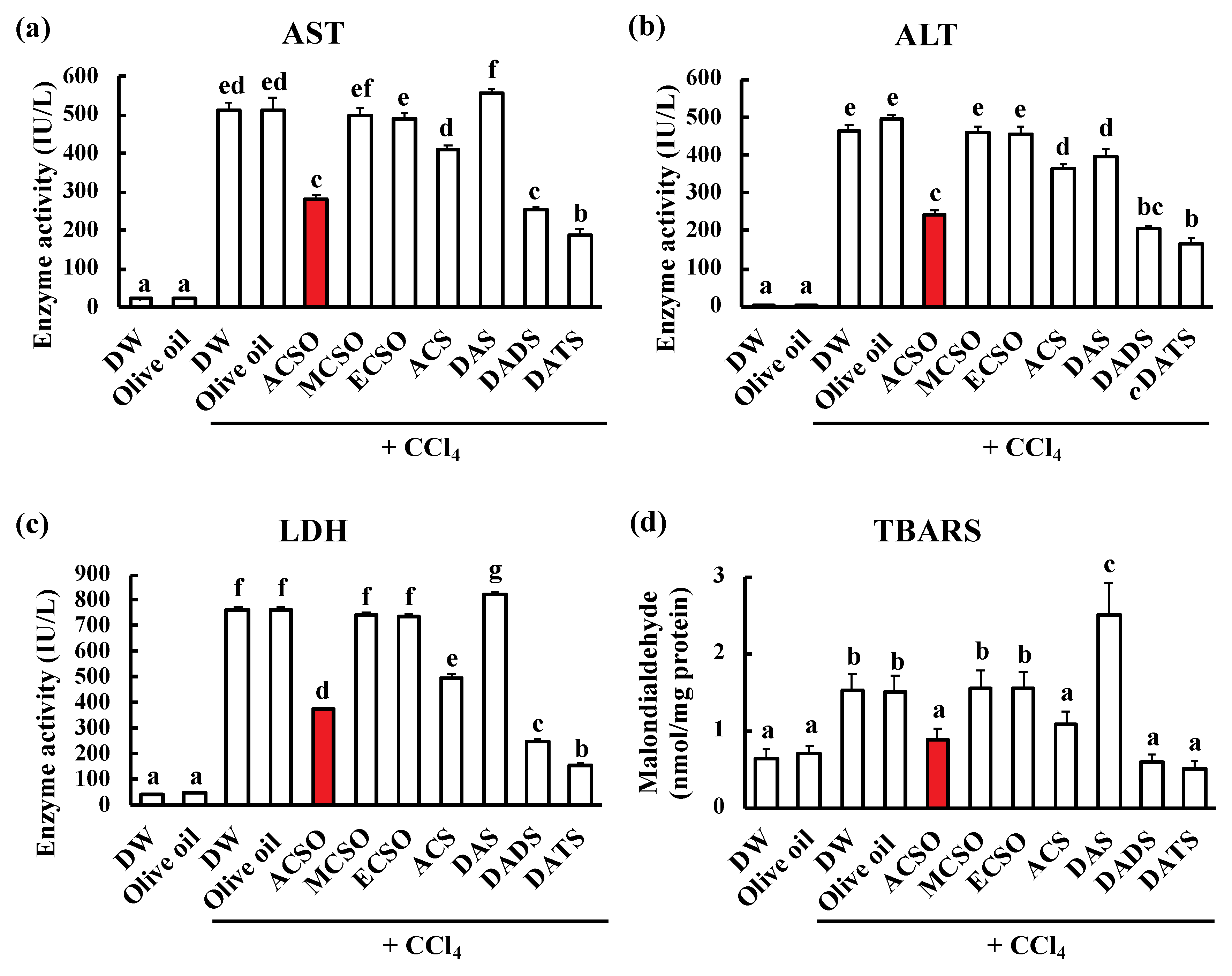
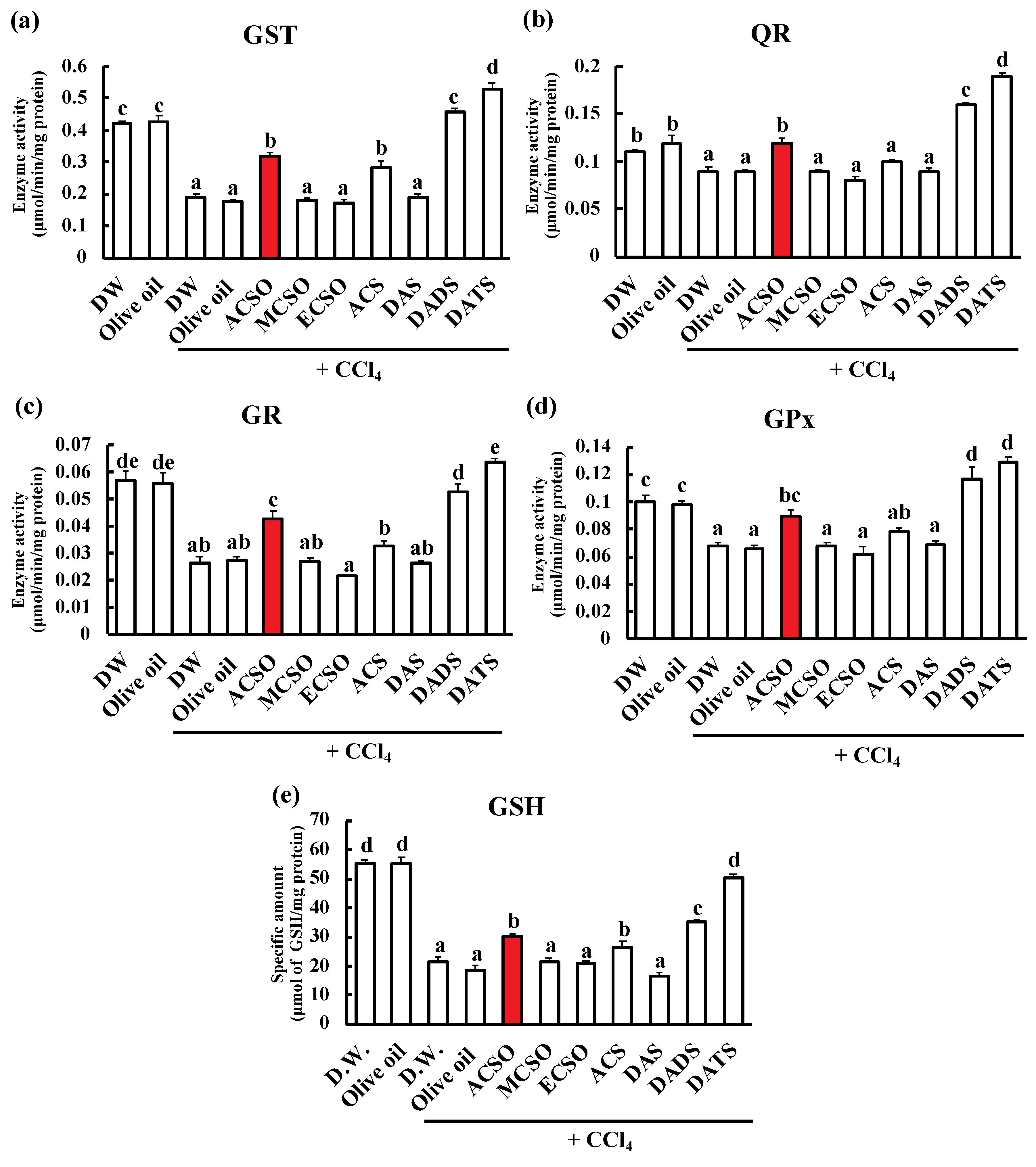
3.1. Effect of ACSO, Its Analogs, and Garlic-Odor Compounds on Suppression of Hepatic Injury Induced by CCl4
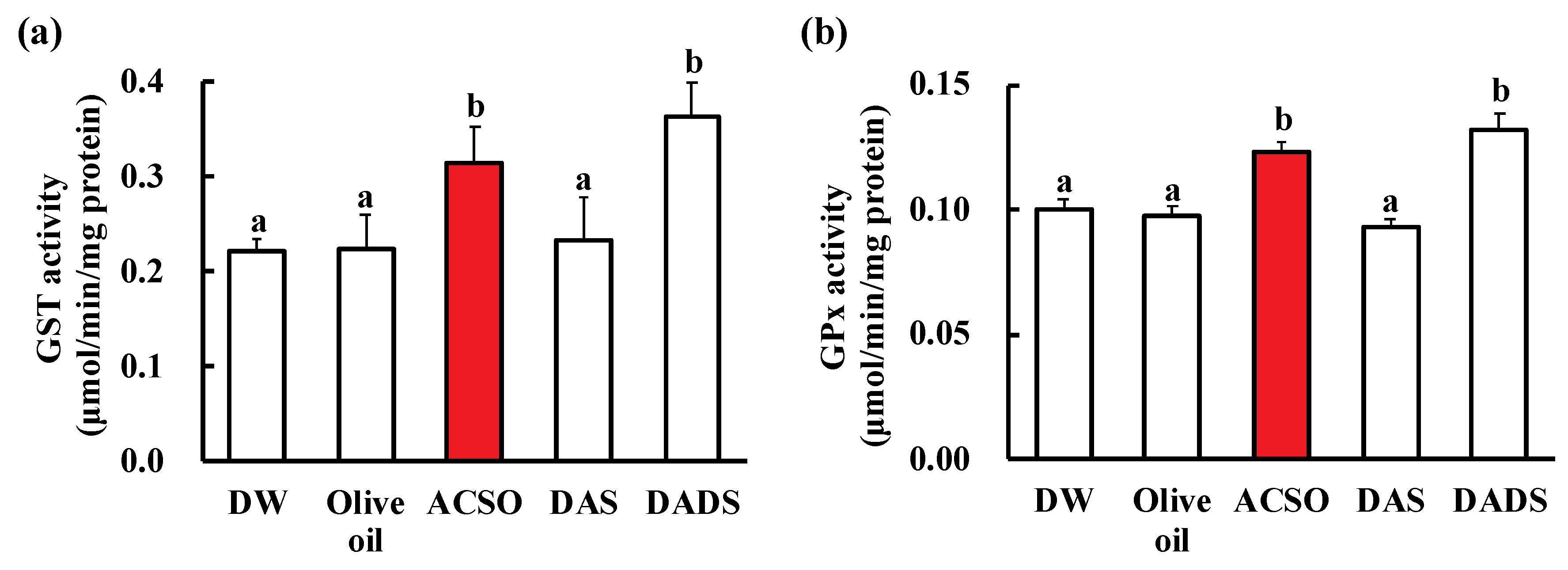
3.2. Effect of ACSO and Sulfides on Liver Function
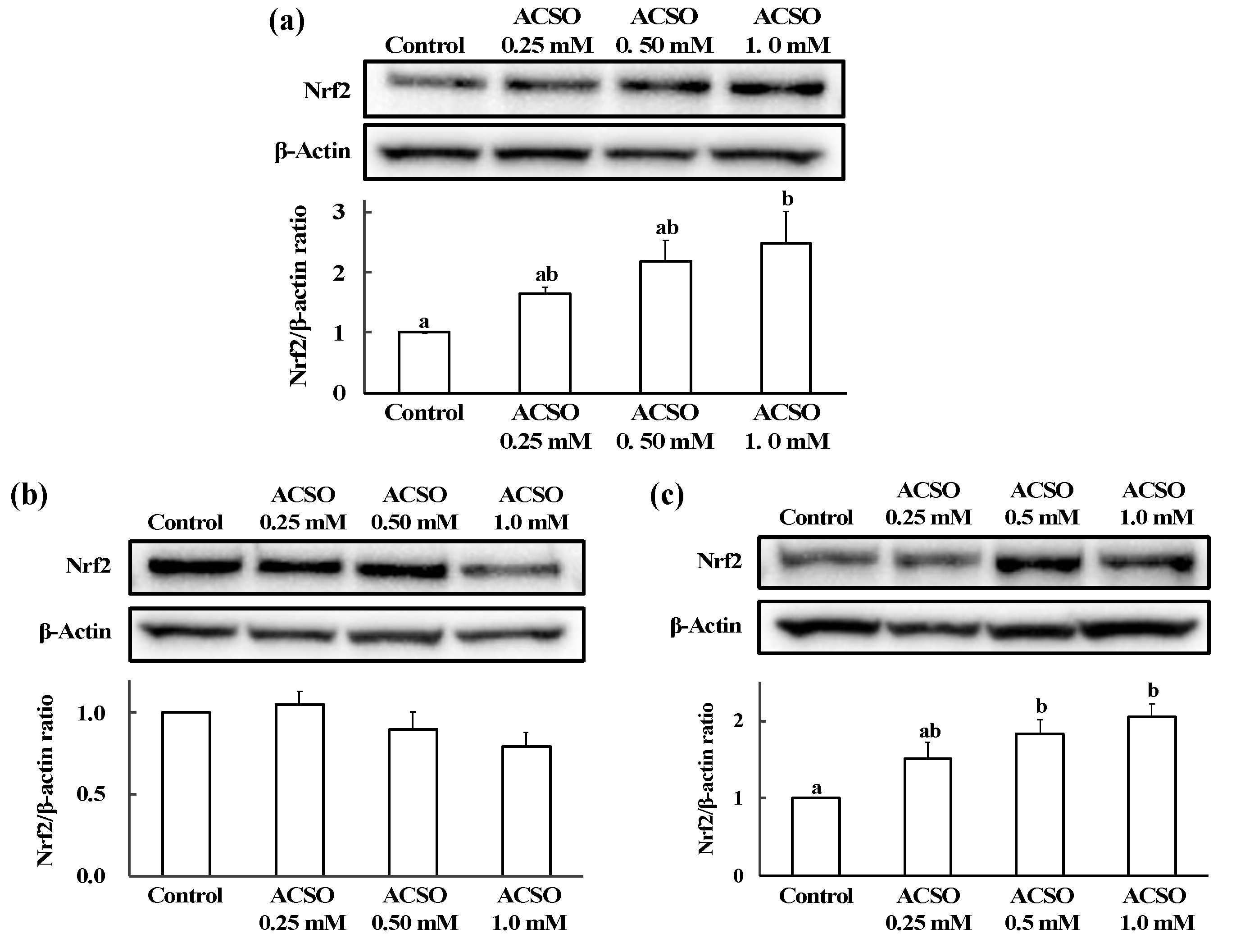
3.3. Effect of ACSO on Nrf2 Nuclear Translocation in HepG2 Cells
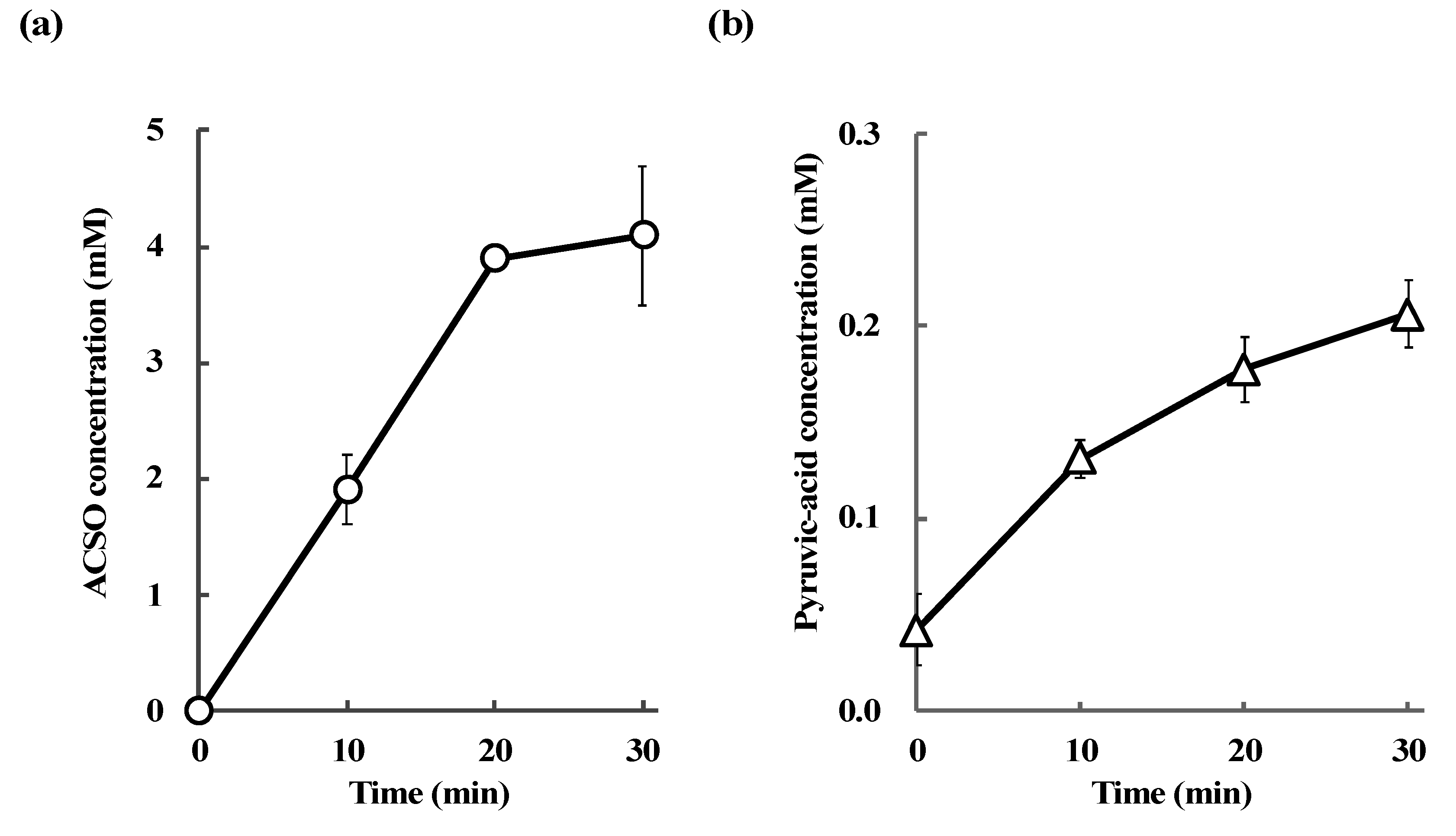
3.4. Absorption and Metabolism of ACSO in the Small Intestine
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hayes, J.D.; McLellan, L.I. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress. Free Radic. Res. 1999, 31, 273–300. [Google Scholar] [CrossRef] [PubMed]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Ketterer, B.; Coles, B.; Meyer, D.J. The role of glutathione in detoxication. Environ. Health Perspect. 1983, 49, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Coles, B.; Ketterer, B. The role of glutathione and glutathione transferases in chemical carcinogenesis. Crit. Rev. Biochem. Mol. Biol. 1990, 25, 47–70. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.; Bradley, R.D. Effects of oral glutathione supplementation on systemic oxidative stress biomarkers in human volunteers. J. Altern. Complement. Med. 2011, 17, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, B.; Vicenzi, M.; Garrel, C.; Denis, F.M. Effects of N-acetylcysteine, oral glutathione (GSH) and a novel sublingual form of GSH on oxidative stress markers: A comparative crossover study. Redox Biol. 2015, 6, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Favilli, F.; Marraccini, P.; Iantomasi, T.; Vincenzini, M.T. Effect of orally administered glutathione on glutathione levels in some organs of rats: Role of specific transporters. Br. J. Nutr. 1997, 78, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Stark, A.-A.; Zeiger, E.; Pagano, D.A. Glutathione metabolism by γ-glutamyltranspeptidase leads to lipid peroxidation: Characterization of the system and relevance to hepatocarcinogenesis. Carcinogenesis 1993, 14, 183–189. [Google Scholar] [CrossRef]
- Nakamura, Y.; Miyoshi, N. Electrophiles in Foods: The current status of isothiocyanates and their chemical biology. Biosci. Biotechnol. Biochem. 2010, 74, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Parvez, S.; Long, M.J.C.; Poganik, J.R.; Aye, Y. Redox signaling by reactive electrophiles and oxidants. Chem. Rev. 2018, 118, 8798–8888. [Google Scholar] [CrossRef]
- Thomson, M.; Ali, M. A review of its potential use as an anti-cancer agent. Curr. Cancer Drug Tar. 2003, 3, 67–81. [Google Scholar] [CrossRef]
- Shukla, Y.; Kalra, N. Cancer chemoprevention with garlic and its constituents. Cancer Lett. 2007, 247, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Powolny, A.A.; Singh, S.V. Multitargeted prevention and therapy of cancer by diallyl trisulfide and related allium vegetable-derived organosulfur compounds. Cancer Lett. 2008, 269, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.H.; Efendy, J.L.; Smith, N.J.; Campbell, G.R. Molecular basis by which garlic suppresses atherosclerosis. J. Nutr. 2001, 31, 1006S–1009S. [Google Scholar] [CrossRef] [PubMed]
- Ohaeri, O.C. Effect of garlic oil on the levels of various enzymes in the serum and tissue of streptozotocin diabetic rats. Biosci. Rep. 2001, 21, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.K.; Iqbal, M.; Athar, M. Garlic oil ameliorates ferric nitrilotriacetate (Fe-NTA)-induced damage and tumor promotion: Implications for cancer prevention. Food Chem. Toxicol. 2007, 45, 1634–1640. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, H.; Watanabe, K.; Suma, K.; Origuchi, K.; Matsufuji, H.; Seki, T.; Ariga, T. Antibacterial potential of garlic-derived allicin and its cancellation by sulfhydryl compounds. Biosci. Biotechnol. Biochem. 2009, 73, 1948–1955. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.K.; Maulik, M.; Mancahanda, S.C.; Dinda, A.K.; Gupta, S.K.; Maulik, S.K. Dose-dependent induction of endogenous antioxidants in rat heart by chronic administration of garlic. Life Sci. 2002, 70, 1509–1518. [Google Scholar] [CrossRef]
- Wu, C.-C.; Sheen, L.-Y.; Chen, H.-W.; Kuo, W.-W.; Tsai, S.-J.; Lii, C.-K. Differential effects of garlic oil and its three major organosulfur components on the hepatic detoxification system in rats. J. Agric. Food Chem. 2002, 50, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.G.; Hughes, J.; Tregova, A.; Milne, J.; Tomsett, A.B.; Collin, H.A. Biosynthesis of the flavour precursors of onion and garlic. J. Exp. Bot. 2004, 55, 1903–1918. [Google Scholar] [CrossRef]
- Augusti, K.T.; Sheela, C.G. Antiperoxide Effect of S-allyl cysteine sulfoxide, an insulin secretagogue, in diabetic rats. Experientia 1996, 52, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Sangeetha, T.; Quine, S.D. Preventive Effect of S-allyl cysteine sulfoxide (Alliin) on cardiac marker enzymes and lipids in isoproterenol-induced myocardial injury. J. Pharm. Pharmacol. 2006, 58, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Zhai, B.; Zhang, C.; Sheng, Y.; Zhao, C.; He, X.; Xu, W.; Huang, K.; Luo, Y. Hypoglycemic and hypolipidemic effect of S-allyl-cysteine sulfoxide (alliin) in DIO mice. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Akao, M.; Shibuya, T.; Shimada, S.; Sakurai, H.; Kumagai, H. In vivo production of bioactive compounds from S-allyl-l-cysteine sulfoxide, garlic odor precursor, that inhibit platelet aggregation. J. Clin. Biochem. Nutr. Supple. 2008, 43, 1–3. [Google Scholar]
- Uto-Kondo, H.; Hase, A.; Yamaguchi, Y.; Sakurai, A.; Akao, M.; Saito, T.; Kumagai, H. S-allyl-l-cysteine sulfoxide, a garlic odor precursor, suppresses elevation in blood ethanol concentration by accelerating ethanol metabolism and preventing ethanol absorption from gut. Biosci. Biotech. Biochem. 2018, 82, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Lawson, L.D. Garlic: A review of its medicinal effects and indicated active compounds. In ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1998; pp. 176–209. [Google Scholar] [CrossRef]
- Ueda, Y.; Sakaguchi, M.; Hirayama, K.; Miyajima, R.; Kimizuka, A. Characteristic flavor constituents in water extract of garlic. Agric. Biol. Chem. 1990, 54, 163–169. [Google Scholar]
- Jayakumar, T.; Ramesh, E.; Geraldine, P. Antioxidant activity of the oyster mushroom, pleurotus ostreatus, on CCl4-induced liver injury in rats. Food. Chem. Toxicol. 2006, 44, 1989–1996. [Google Scholar] [CrossRef] [PubMed]
- Huo, H.Z.; Wang, B.; Liang, Y.K.; Bao, Y.Y.; Gu, Y. Hepatoprotective and antioxidant effects of licorice extract against CCl4-induced oxidative damage in rats. Int. J. Mol. Sci. 2011, 12, 6529–6543. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Ren, N.; Gao, H.; Lei, X.; Zheng, J.; Cao, W. Antioxidant and hepatoprotective effects of schisandra chinensis pollen extract on CCl4-induced acute liver damage in mice. Food. Chem. Toxicol. 2013, 55, 234–240. [Google Scholar] [CrossRef]
- Pan, Y.; Long, X.; Yi, R.; Zhao, X. Polyphenols in liubao tea can prevent CCl4-induced hepatic damage in mice through its antioxidant capacities. Nutrients 2018, 10, 1280. [Google Scholar] [CrossRef]
- Raucy, J.L.; Kraner, J.C.; Lasker, J.M. Bioactivation of halogenated hydrocarbons by cytochrome P4502E1. Crit. Rev. Toxicol. 1993, 23, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.W.D.; Boll, M.; Stampfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef] [PubMed]
- Recknagel, R.O. A New direction in the study of carbon tetrachloride hepatotoxicity. Life Sci. 1983, 33, 401–408. [Google Scholar] [CrossRef]
- Slater, T.F. Free-radical mechanisms in tissue injury. Biochem. J. 1984, 222, 1–15. [Google Scholar] [CrossRef]
- Ramaiah, S.K. A Toxicologist guide to the diagnostic interpretation of hepatic biochemical parameters. Food Chem. Toxicol. 2007, 45, 1551–1557. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef]
- Bataille, A.M.; Manautou, J.E. Nrf2: A potential target for new therapeutics in liver disease. Clin. Pharmacol. Ther. 2012, 92, 340–348. [Google Scholar] [CrossRef]
- Hakamata, W.; Koyama, R.; Tanida, M.; Haga, T.; Hirano, T.; Akao, M.; Kumagai, H.; Nishio, T. A simple synthesis of alliin and allo-alliin: X-ray diffraction analysis and determination of their absolute configurations. J. Agric. Food Chem. 2015, 63, 10778–10784. [Google Scholar] [CrossRef]
- Haber, D.; Siess, M.-H.; De Waziers, I.; Beaune, P.; Suschetet, M. Modification of hepatic drug-metabolizing enzymes in rat fed naturally occurring allyl sulphides. Xenobiotica 1994, 24, 169–182. [Google Scholar] [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [PubMed]
- Ernster, L.; Danielson, L.; Ljunggren, M. DT diaphorase. I. Purification from the soluble fraction of rat-liver cytoplasm, and properties. Biochim. Biophys. Acta 1962, 58, 171–188. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Method Enzymol. 1985, 113, 484–490. [Google Scholar]
- Yamamoto, Y.; Takahashi, K. Glutathione peroxidase isolated from plasma reduces phospholipid hydroperoxides. Arch. Biochem. Biophys. 1993, 305, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Kodera, Y.; Suzuki, A.; Imada, O.; Kasuga, S.; Sumioka, I.; Kanezawa, A.; Taru, N.; Fujikawa, M.; Nagae, S.; Masamoto, K.; et al. Physical, chemical, and biological properties of S-allylcysteine, an amino acid derived from garlic. J. Agric. Food Chem. 2002, 50, 622–632. [Google Scholar] [CrossRef]
- Mizuguchi, S.; Takemura, S.; Minamiyama, Y.; Kodai, S.; Tsukioka, T.; Inoue, K.; Okada, S.; Suehiro, S. S-allyl cysteine attenuated CCl4-induced oxidative stress and pulmonary fibrosis in rats. Biofactors 2006, 26, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Kodai, S.; Takemura, S.; Minamiyama, Y.; Hai, S.; Yamamoto, S.; Kubo, S.; Yoshida, Y.; Niki, E.; Okada, S.; Hirohashi, K.; et al. S-allyl cysteine prevents CCl4-induced acute liver injury in rats. Free Radic. Res. 2007, 41, 489–497. [Google Scholar] [CrossRef]
- Fukao, T.; Hosono, T.; Misawa, S.; Seki, T.; Ariga, T. The effects of allyl sulfides on the induction of phase II detoxification enzymes and liver injury by carbon tetrachloride. Food Chem. Toxicol. 2004, 42, 743–749. [Google Scholar] [CrossRef]
- Lee, I.-C.; Kim, S.-H.; Baek, H.-S.; Moon, C.; Kang, S.-S.; Kim, S.-H.; Kim, Y.-B.; Shin, I.-S.; Kim, J.-C. The involvement of Nrf2 in the protective effects of diallyl disulfide on carbon tetrachloride-induced hepatic oxidative damage and inflammatory response in rats. Food. Chem. Toxicol. 2014, 63, 174–185. [Google Scholar] [CrossRef]
- Markesbery, W.R. Oxidative stress hypothesis in alzheimer’s disease. Free Radic. Biol. Med. 1997, 23, 134–147. [Google Scholar] [CrossRef]
- Madamanchi, N.R.; Vendrov, A.; Runge, M.S. Oxidative stress and vascular disease. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Eggler, A.L.; Mesecar, A.D.; van Breemen, R.B. Modification of keap1 cysteine residues by sulforaphane. Chem. Res. Toxicol. 2011, 24, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Hosono, T.; Fukao, T.; Ogihara, J.; Ito, Y.; Shiba, H.; Seki, T.; Ariga, T. Diallyl trisulfide suppresses the proliferation and induces apoptosis of human colon cancer cells through oxidative modification of beta-tubulin. J. Biol. Chem. 2005, 280, 41487–41493. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, H.G.; Park, S.A.; Kundu, J.K.; Keum, Y.S.; Cha, Y.N.; Na, H.K.; Surh, Y.J. Keap1 cysteine 288 as a potential target for diallyl trisulfide-induced Nrf2 activation. PLoS ONE 2014, 9, e85984. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ACSO Concentration (mM) | Pyruvic-Acid Concentration (mM) |
|---|---|---|
| PBS | Not detected | 0.04 ± 0.01 |
| ACSO | 8.02 ± 1.84 | 0.81 ± 0.07 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaguchi, Y.; Honma, R.; Yazaki, T.; Shibuya, T.; Sakaguchi, T.; Uto-Kondo, H.; Kumagai, H. Sulfuric Odor Precursor S-Allyl-l-Cysteine Sulfoxide in Garlic Induces Detoxifying Enzymes and Prevents Hepatic Injury. Antioxidants 2019, 8, 385. https://doi.org/10.3390/antiox8090385
Yamaguchi Y, Honma R, Yazaki T, Shibuya T, Sakaguchi T, Uto-Kondo H, Kumagai H. Sulfuric Odor Precursor S-Allyl-l-Cysteine Sulfoxide in Garlic Induces Detoxifying Enzymes and Prevents Hepatic Injury. Antioxidants. 2019; 8(9):385. https://doi.org/10.3390/antiox8090385
Chicago/Turabian StyleYamaguchi, Yusuke, Ryosuke Honma, Tomoaki Yazaki, Takeshi Shibuya, Tomoya Sakaguchi, Harumi Uto-Kondo, and Hitomi Kumagai. 2019. "Sulfuric Odor Precursor S-Allyl-l-Cysteine Sulfoxide in Garlic Induces Detoxifying Enzymes and Prevents Hepatic Injury" Antioxidants 8, no. 9: 385. https://doi.org/10.3390/antiox8090385
APA StyleYamaguchi, Y., Honma, R., Yazaki, T., Shibuya, T., Sakaguchi, T., Uto-Kondo, H., & Kumagai, H. (2019). Sulfuric Odor Precursor S-Allyl-l-Cysteine Sulfoxide in Garlic Induces Detoxifying Enzymes and Prevents Hepatic Injury. Antioxidants, 8(9), 385. https://doi.org/10.3390/antiox8090385