
Immunization with GP1 but Not Core-like Particles Displaying Isolated Receptor-Binding Epitopes Elicits Virus-Neutralizing Antibodies against Junín Virus

 , , , , , , and
, , , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of DNA Constructs for Protein Expression
2.2. Expression and Purification of Strep-Tagged JUNV GP1
2.3. Expression and Purification of Histidine-Tagged JUNV GP1
2.4. Expression and Purification of HBV-CLPs
2.5. Characterization of HBV-CLPs by Electron Microscopy
2.6. Sequencing of JUNV GP1
2.7. ELISA Assay
2.8. Immunization and Collection of Serum Samples
2.9. Transcription and Replication-Competent Virus-like Particle (trVLP) Neutralization Assay
2.10. Plaque Reduction Neutralization Assay
2.11. Statistical Analysis
3. Results
3.1. Immunogen Design
3.2. Expression and Purification of Recombinant GP1 and HBV–CLPs Expressing JUNV GP1 Peptide Loops
3.3. Immunization with rGP1 Elicits Antibodies with Neutralizing Activity
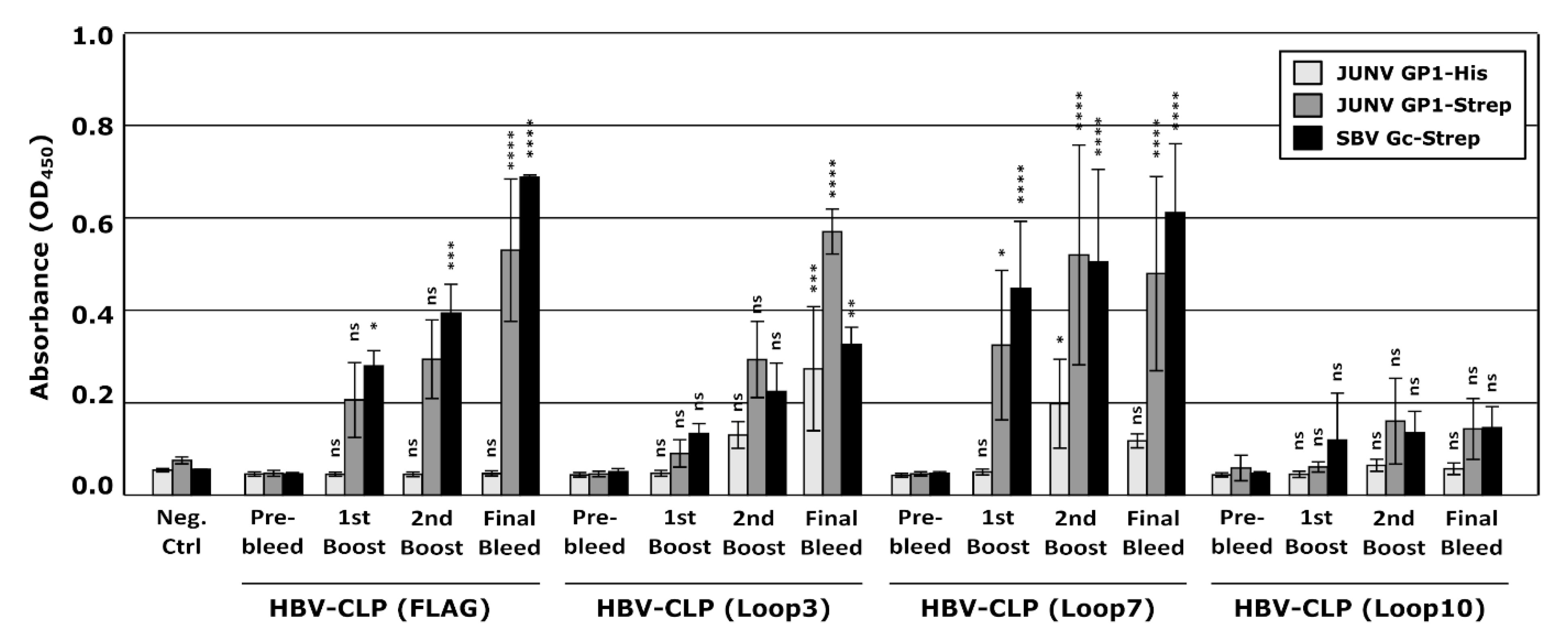
3.4. HBV-CLPs Expressing Receptor-Interacting Loops of JUNV GP1 Elicit Antibodies, but These Do Not Have Neutralizing Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monath, T.P. A short history of Lassa fever: The first 10–15 years after discovery. Curr. Opin. Virol. 2019, 37, 77–83. [Google Scholar] [CrossRef]
- Sarute, N.; Ross, S.R. New World Arenavirus Biology. Annu. Rev. Virol. 2017, 4, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Peters, C.J. Human infection with arenaviruses in the Americas. Curr. Top. Microbiol. Immunol. 2002, 262, 65–74. [Google Scholar] [CrossRef]
- Tesh, R.B. Viral hemorrhagic fevers of South America. Biomedica 2002, 22, 287–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, J.N.; Ellis, B.A.; Childs, J.E.; McKee, K.T.; Maiztegui, J.I.; Peters, C.J.; Ksiazek, T.G.; Jahrling, P.B. Prevalence of infection with Junin virus in rodent populations in the epidemic area of Argentine hemorrhagic fever. Am. J. Trop. Med. Hyg. 1994, 51, 554–562. [Google Scholar] [CrossRef]
- Ambrosio, A.; Saavedra, M.; Mariani, M.; Gamboa, G.; Maiza, A. Argentine hemorrhagic fever vaccines. Hum. Vaccin. 2011, 7, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Rojek, J.M.; Lee, A.M.; Nguyen, N.; Spiropoulou, C.F.; Kunz, S. Site 1 protease is required for proteolytic processing of the glycoproteins of the South American hemorrhagic fever viruses Junin, Machupo, and Guanarito. J. Virol. 2008, 82, 6045–6051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burri, D.J.; Da Palma, J.R.; Kunz, S.; Pasquato, A. Envelope glycoprotein of arenaviruses. Viruses 2012, 4, 2162–2181. [Google Scholar] [CrossRef]
- Helguera, G.; Jemielity, S.; Abraham, J.; Cordo, S.M.; Martinez, M.G.; Rodríguez, J.A.; Bregni, C.; Wang, J.J.; Farzan, M.; Penichet, M.L.; et al. An antibody recognizing the apical domain of human transferrin receptor 1 efficiently inhibits the entry of all new world hemorrhagic Fever arenaviruses. J. Virol. 2012, 86, 4024–4028. [Google Scholar] [CrossRef] [Green Version]
- Radoshitzky, S.R.; Abraham, J.; Spiropoulou, C.F.; Kuhn, J.H.; Nguyen, D.; Li, W.; Nagel, J.; Schmidt, P.J.; Nunberg, J.H.; Andrews, N.C.; et al. Transferrin receptor 1 is a cellular receptor for New World haemorrhagic fever arenaviruses. Nature 2007, 446, 92–96. [Google Scholar] [CrossRef]
- Abraham, J.; Corbett, K.D.; Farzan, M.; Choe, H.; Harrison, S.C. Structural basis for receptor recognition by New World hemorrhagic fever arenaviruses. Nat. Struct. Mol. Biol. 2010, 17, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Mahmutovic, S.; Clark, L.; Levis, S.C.; Briggiler, A.M.; Enria, D.A.; Harrison, S.C.; Abraham, J. Molecular Basis for Antibody-Mediated Neutralization of New World Hemorrhagic Fever Mammarenaviruses. Cell Host Microbe 2015, 18, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, C.O.; Jette, C.A.; Abernathy, M.E.; Dam, K.-M.A.; Esswein, S.R.; Gristick, H.B.; Malyutin, A.G.; Sharaf, N.G.; Huey-Tubman, K.E.; Lee, Y.E.; et al. SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature 2020, 588, 682–687. [Google Scholar] [CrossRef]
- Srivastava, V.; Niu, L.; Phadke, K.S.; Bellaire, B.H.; Cho, M.W. Induction of Potent and Durable Neutralizing Antibodies Against SARS-CoV-2 Using a Receptor Binding Domain-Based Immunogen. Front. Immunol. 2021, 12, 637982. [Google Scholar] [CrossRef] [PubMed]
- Aricescu, A.R.; Lu, W.; Jones, E.Y. A time- and cost-efficient system for high-level protein production in mammalian cells. Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Kratz, P.A.; Böttcher, B.; Nassal, M. Native display of complete foreign protein domains on the surface of hepatitis B virus capsids. Proc. Natl. Acad. Sci. USA 1999, 96, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Roman-Sosa, G.; Brocchi, E.; Schirrmeier, H.; Wernike, K.; Schelp, C.; Beer, M. Analysis of the humoral immune response against the envelope glycoprotein Gc of Schmallenberg virus reveals a domain located at the amino terminus targeted by mAbs with neutralizing activity. J. Gen. Virol. 2016, 97, 571–580. [Google Scholar] [CrossRef]
- Krieger, J.; Riedl, P.; Stifter, K.; Roman-Sosa, G.; Seufferlein, T.; Wagner, M.; Schirmbeck, R. Endogenously Expressed Antigens Bind Mammalian RNA via Cationic Domains that Enhance Priming of Effector CD8 T Cells by DNA Vaccination. Mol. Ther. 2019, 27, 661–672. [Google Scholar] [CrossRef]
- Jangra, R.K.; Herbert, A.S.; Li, R.; Jae, L.T.; Kleinfelter, L.M.; Slough, M.M.; Barker, S.L.; Guardado-Calvo, P.; Román-Sosa, G.; Dieterle, M.E.; et al. Protocadherin-1 is essential for cell entry by New World hantaviruses. Nature 2018, 563, 559–563. [Google Scholar] [CrossRef]
- Dunham, E.C.; Leske, A.; Shifflett, K.; Watt, A.; Feldmann, H.; Hoenen, T.; Groseth, A. Lifecycle modelling systems support inosine monophosphate dehydrogenase (IMPDH) as a pro-viral factor and antiviral target for New World arenaviruses. Antiviral Res. 2018, 157, 140–150. [Google Scholar] [CrossRef]
- Leske, A.; Waßmann, I.; Schnepel, K.; Shifflett, K.; Holzerland, J.; Bostedt, L.; Bohn, P.; Mettenleiter, T.C.; Briggiler, A.M.; Brignone, J.; et al. Assessing cross-reactivity of Junín virus-directed neutralizing antibodies. Antiviral Res. 2019, 163, 106–116. [Google Scholar] [CrossRef]
- Manning, J.T.; Seregin, A.V.; Yun, N.E.; Koma, T.; Huang, C.; Barral, J.; de La Torre, J.C.; Paessler, S. Absence of an N-Linked Glycosylation Motif in the Glycoprotein of the Live-Attenuated Argentine Hemorrhagic Fever Vaccine, Candid #1, Results in Its Improper Processing, and Reduced Surface Expression. Front. Cell. Infect. Microbiol. 2017, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Del Carmen Saavedra, M.; Sottosanti, J.M.; Riera, L.; Ambrosio, A.M. IgG subclasses in human immune response to wild and attenuated (vaccine) Junin virus infection. J. Med. Virol. 2003, 69, 447–450. [Google Scholar] [CrossRef]
- Enria, D.A.; Briggiler, A.M.; Fernandez, N.J.; Levis, S.C.; Maiztegui, J.I. Importance of dose of neutralising antibodies in treatment of Argentine haemorrhagic fever with immune plasma. Lancet 1984, 2, 255–256. [Google Scholar] [CrossRef]
- Golden, J.W.; Maes, P.; Kwilas, S.A.; Ballantyne, J.; Hooper, J.W. Glycoprotein-Specific Antibodies Produced by DNA Vaccination Protect Guinea Pigs from Lethal Argentine and Venezuelan Hemorrhagic Fever. J. Virol. 2016, 90, 3515–3529. [Google Scholar] [CrossRef] [Green Version]
- Zeitlin, L.; Cross, R.W.; Geisbert, J.B.; Borisevich, V.; Agans, K.N.; Prasad, A.N.; Enterlein, S.; Aman, M.J.; Bornholdt, Z.A.; Brennan, M.B.; et al. Therapy for Argentine hemorrhagic fever in nonhuman primates with a humanized monoclonal antibody. Proc. Natl. Acad. Sci. USA 2021, 118, e2023332118. [Google Scholar] [CrossRef] [PubMed]
- Seregin, A.V.; Yun, N.E.; Poussard, A.L.; Peng, B.-H.; Smith, J.K.; Smith, J.N.; Salazar, M.; Paessler, S. TC83 replicon vectored vaccine provides protection against Junin virus in guinea pigs. Vaccine 2010, 28, 4713–4718. [Google Scholar] [CrossRef] [PubMed]
- Borenstein-Katz, A.; Shulman, A.; Hamawi, H.; Leitner, O.; Diskin, R. Differential Antibody-Based Immune Response against Isolated GP1 Receptor-Binding Domains from Lassa and Junín Viruses. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Wu, Y.; Wang, W.; Zhang, L.; Xiao, G. Development of horse neutralizing immunoglobulin and immunoglobulin fragments against Junín virus. Antiviral Res. 2020, 174, 104666. [Google Scholar] [CrossRef]
- Singh, M.; Sori, H.; Ahuja, R.; Meena, J.; Sehgal, D.; Panda, A.K. Effect of N-terminal poly histidine-tag on immunogenicity of Streptococcus pneumoniae surface protein SP0845. Int. J. Biol. Macromol. 2020, 163, 1240–1248. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Fehling, S.K.; Dorna, J.; Urbanowicz, R.A.; Oestereich, L.; Krebs, Y.; Kolesnikova, L.; Schauflinger, M.; Krähling, V.; Magassouba, N.; et al. Adjuvant formulated virus-like particles expressing native-like forms of the Lassa virus envelope surface glycoprotein are immunogenic and induce antibodies with broadly neutralizing activity. NPJ Vaccines 2020, 5, 71. [Google Scholar] [CrossRef]
- Pan, X.; Wu, Y.; Wang, W.; Zhang, L.; Xiao, G. Novel neutralizing monoclonal antibodies against Junin virus. Antivir. Res. 2018, 156, 21–28. [Google Scholar] [CrossRef]
- Zeltina, A.; Krumm, S.A.; Sahin, M.; Struwe, W.B.; Harlos, K.; Nunberg, J.H.; Crispin, M.; Pinschewer, D.D.; Doores, K.J.; Bowden, T.A. Convergent immunological solutions to Argentine hemorrhagic fever virus neutralization. Proc. Natl. Acad. Sci. USA 2017, 114, 7031–7036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, A.; Pifat, D.Y.; Kenyon, R.H.; Peters, C.J.; McCormick, J.B.; Kiley, M.P. Junin virus monoclonal antibodies: Characterization and cross-reactivity with other arenaviruses. J. Gen. Virol. 1989, 70 Pt 5, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.E.; Mahmutovic, S.; Raymond, D.D.; Dilanyan, T.; Koma, T.; Manning, J.T.; Shankar, S.; Levis, S.C.; Briggiler, A.M.; Enria, D.A.; et al. Vaccine-elicited receptor-binding site antibodies neutralize two New World hemorrhagic fever arenaviruses. Nat. Commun. 2018, 9, 1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makdasi, E.; Levy, Y.; Alcalay, R.; Noy-Porat, T.; Zahavy, E.; Mechaly, A.; Epstein, E.; Peretz, E.; Cohen, H.; Bar-On, L.; et al. Neutralizing Monoclonal Anti-SARS-CoV-2 Antibodies Isolated from Immunized Rabbits Define Novel Vulnerable Spike-Protein Epitope. Viruses 2021, 13, 566. [Google Scholar] [CrossRef]
- Zhang, Y.-F.; Ho, M. Humanization of rabbit monoclonal antibodies via grafting combined Kabat/IMGT/Paratome complementarity-determining regions: Rationale and examples. MAbs 2017, 9, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoenen, T.; Groseth, A.; Feldmann, H. Therapeutic strategies to target the Ebola virus life cycle. Nat. Rev. Microbiol. 2019, 17, 593–606. [Google Scholar] [CrossRef]
- Wernike, K.; Aebischer, A.; Roman-Sosa, G.; Beer, M. The N-terminal domain of Schmallenberg virus envelope protein Gc is highly immunogenic and can provide protection from infection. Sci. Rep. 2017, 7, 42500. [Google Scholar] [CrossRef] [Green Version]
- Zakeri, B.; Fierer, J.O.; Celik, E.; Chittock, E.C.; Schwarz-Linek, U.; Moy, V.T.; Howarth, M. Peptide tag forming a rapid covalent bond to a protein, through engineering a bacterial adhesin. Proc. Natl. Acad. Sci. USA 2012, 109, E690–E697. [Google Scholar] [CrossRef] [Green Version]
- Okba, N.M.A.; Widjaja, I.; van Dieren, B.; Aebischer, A.; van Amerongen, G.; de Waal, L.; Stittelaar, K.J.; Schipper, D.; Martina, B.; van den Brand, J.M.A.; et al. Particulate multivalent presentation of the receptor binding domain induces protective immune responses against MERS-CoV. Emerg. Microbes Infect. 2020, 9, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, S.; Flores, M.D.; Short, C.; Vazquez, C.A.; Clark, L.E.; Ziegenbein, J.; Zink, S.; Fuentes, D.; Payes, C.; Batto, M.V.; et al. Antibody-Based Inhibition of Pathogenic New World Hemorrhagic Fever Mammarenaviruses by Steric Occlusion of the Human Transferrin Receptor 1 Apical Domain. J. Virol. 2021, 95, e0186820. [Google Scholar] [CrossRef]
- Cohen-Dvashi, H.; Amon, R.; Agans, K.N.; Cross, R.W.; Borenstein-Katz, A.; Mateo, M.; Baize, S.; Padler-Karavani, V.; Geisbert, T.W.; Diskin, R. Rational design of universal immunotherapy for TfR1-tropic arenaviruses. Nat. Commun. 2020, 11, 67. [Google Scholar] [CrossRef]
- van Regenmortel, M.H. Antigenicity and immunogenicity of synthetic peptides. Biologicals 2001, 29, 209–213. [Google Scholar] [CrossRef]
- Gedvilaite, A.; Kucinskaite-Kodze, I.; Lasickiene, R.; Timinskas, A.; Vaitiekaite, A.; Ziogiene, D.; Zvirbliene, A. Evaluation of Trichodysplasia Spinulosa-Associated Polyomavirus Capsid Protein as a New Carrier for Construction of Chimeric Virus-Like Particles Harboring Foreign Epitopes. Viruses 2015, 7, 4204–4229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zvirbliene, A.; Kucinskaite-Kodze, I.; Razanskiene, A.; Petraityte-Burneikiene, R.; Klempa, B.; Ulrich, R.G.; Gedvilaite, A. The use of chimeric virus-like particles harbouring a segment of hantavirus Gc glycoprotein to generate a broadly-reactive hantavirus-specific monoclonal antibody. Viruses 2014, 6, 640–660. [Google Scholar] [CrossRef] [Green Version]
- Snapper, C.M. Distinct Immunologic Properties of Soluble Versus Particulate Antigens. Front. Immunol. 2018, 9, 598. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Rohrer, U.H.; Kündig, T.M.; Bürki, K.; Hengartner, H.; Zinkernagel, R.M. The influence of antigen organization on B cell responsiveness. Science 1993, 262, 1448–1451. [Google Scholar] [CrossRef]
- Freivalds, J.; Dislers, A.; Ose, V.; Pumpens, P.; Tars, K.; Kazaks, A. Highly efficient production of phosphorylated hepatitis B core particles in yeast Pichia pastoris. Protein Expr. Purif. 2011, 75, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Wynne, S.A.; Crowther, R.A.; Leslie, A.G. The crystal structure of the human hepatitis B virus capsid. Mol. Cell 1999, 3, 771–780. [Google Scholar] [CrossRef]
- Akhras, S.; Toda, M.; Boller, K.; Himmelsbach, K.; Elgner, F.; Biehl, M.; Scheurer, S.; Gratz, M.; Vieths, S.; Hildt, E. Cell-permeable capsids as universal antigen carrier for the induction of an antigen-specific CD8+ T-cell response. Sci. Rep. 2017, 7, 9630. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, R.; Nassal, M.; Meisel, H.; Krüger, D.H. Core particles of hepatitis B virus as carrier for foreign epitopes. Adv. Virus Res. 1998, 50, 141–182. [Google Scholar] [CrossRef] [PubMed]
- Schödel, F.; Moriarty, A.M.; Peterson, D.L.; Zheng, J.A.; Hughes, J.L.; Will, H.; Leturcq, D.J.; McGee, J.S.; Milich, D.R. The position of heterologous epitopes inserted in hepatitis B virus core particles determines their immunogenicity. J. Virol. 1992, 66, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitacre, D.C.; Lee, B.O.; Milich, D.R. Use of hepadnavirus core proteins as vaccine platforms. Expert Rev. Vaccines 2009, 8, 1565–1573. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roman-Sosa, G.; Leske, A.; Ficht, X.; Dau, T.H.; Holzerland, J.; Hoenen, T.; Beer, M.; Kammerer, R.; Schirmbeck, R.; Rey, F.A.; et al. Immunization with GP1 but Not Core-like Particles Displaying Isolated Receptor-Binding Epitopes Elicits Virus-Neutralizing Antibodies against Junín Virus. Vaccines 2022, 10, 173. https://doi.org/10.3390/vaccines10020173
Roman-Sosa G, Leske A, Ficht X, Dau TH, Holzerland J, Hoenen T, Beer M, Kammerer R, Schirmbeck R, Rey FA, et al. Immunization with GP1 but Not Core-like Particles Displaying Isolated Receptor-Binding Epitopes Elicits Virus-Neutralizing Antibodies against Junín Virus. Vaccines. 2022; 10(2):173. https://doi.org/10.3390/vaccines10020173
Chicago/Turabian StyleRoman-Sosa, Gleyder, Anne Leske, Xenia Ficht, Tung Huy Dau, Julia Holzerland, Thomas Hoenen, Martin Beer, Robert Kammerer, Reinhold Schirmbeck, Felix A. Rey, and et al. 2022. "Immunization with GP1 but Not Core-like Particles Displaying Isolated Receptor-Binding Epitopes Elicits Virus-Neutralizing Antibodies against Junín Virus" Vaccines 10, no. 2: 173. https://doi.org/10.3390/vaccines10020173
APA StyleRoman-Sosa, G., Leske, A., Ficht, X., Dau, T. H., Holzerland, J., Hoenen, T., Beer, M., Kammerer, R., Schirmbeck, R., Rey, F. A., Cordo, S. M., & Groseth, A. (2022). Immunization with GP1 but Not Core-like Particles Displaying Isolated Receptor-Binding Epitopes Elicits Virus-Neutralizing Antibodies against Junín Virus. Vaccines, 10(2), 173. https://doi.org/10.3390/vaccines10020173