
Durability and Cross-Reactivity of SARS-CoV-2 mRNA Vaccine in Adolescent Children

 ,
,  ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Sample Collection
2.2. Enzyme-Linked Immunosorbent Assay (ELISA)
2.3. Virus Neutralization
2.4. Antibody-Dependent-Complement-Deposition (ADCD)
2.5. Antibody-Dependent-THP-1 Cellular-Phagocytosis (ADCP)
2.6. Antibody-Dependent-Neutrophil-Phagocytosis (ADNP)
2.7. Statistical Analysis
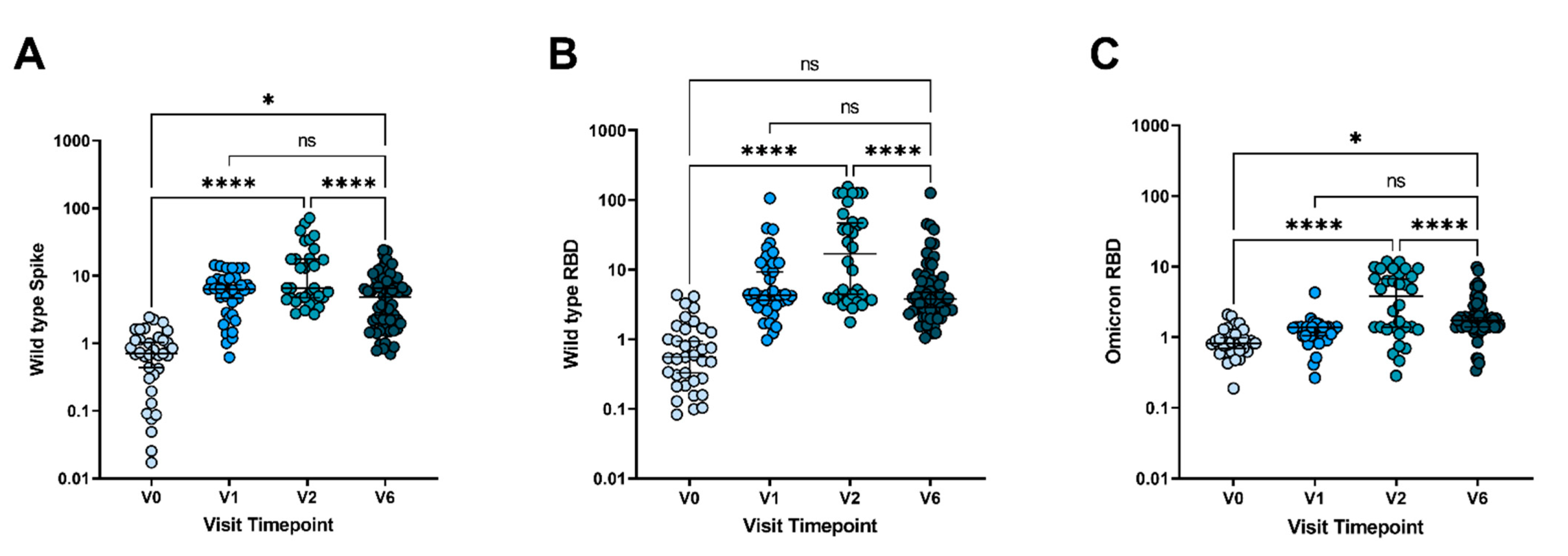
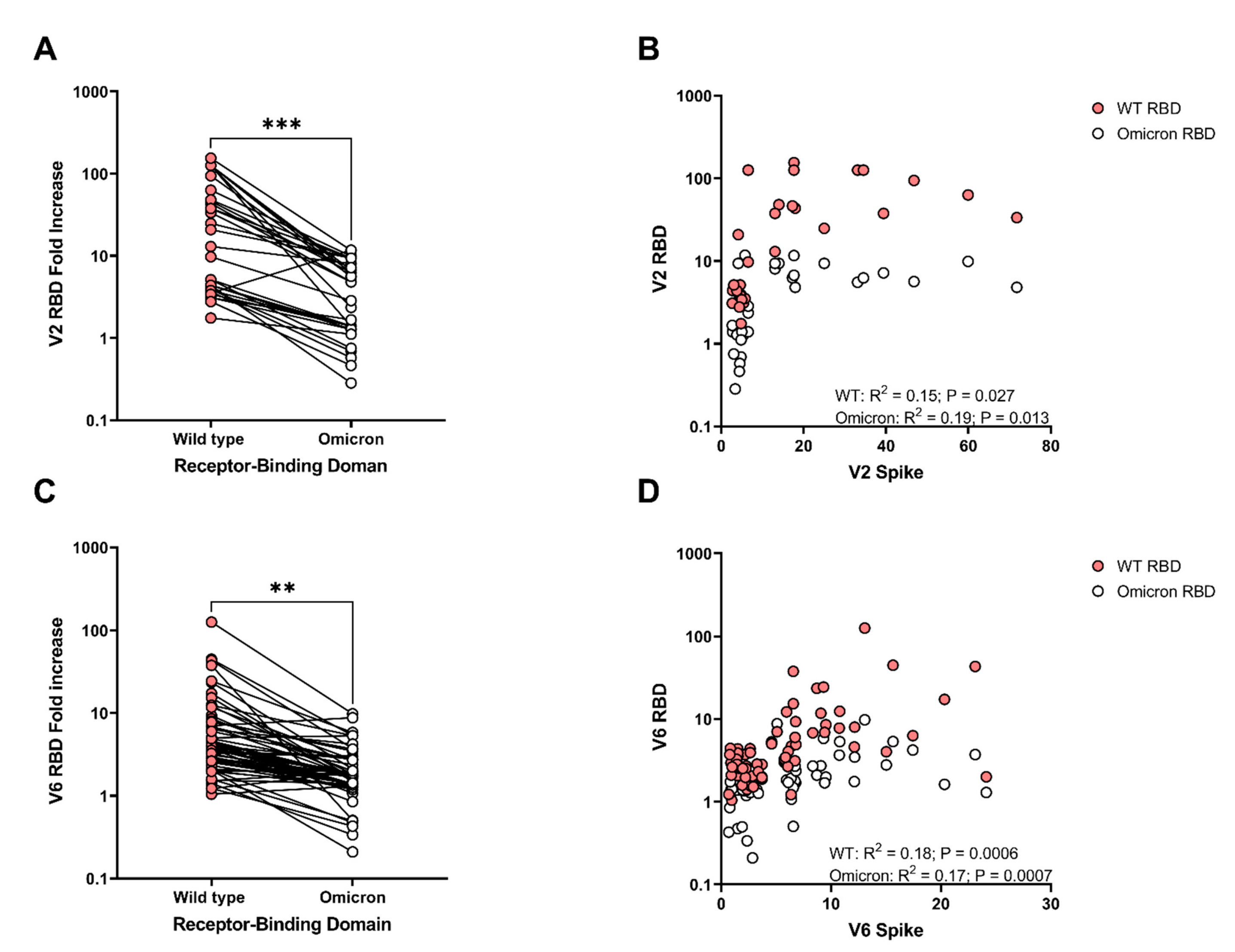
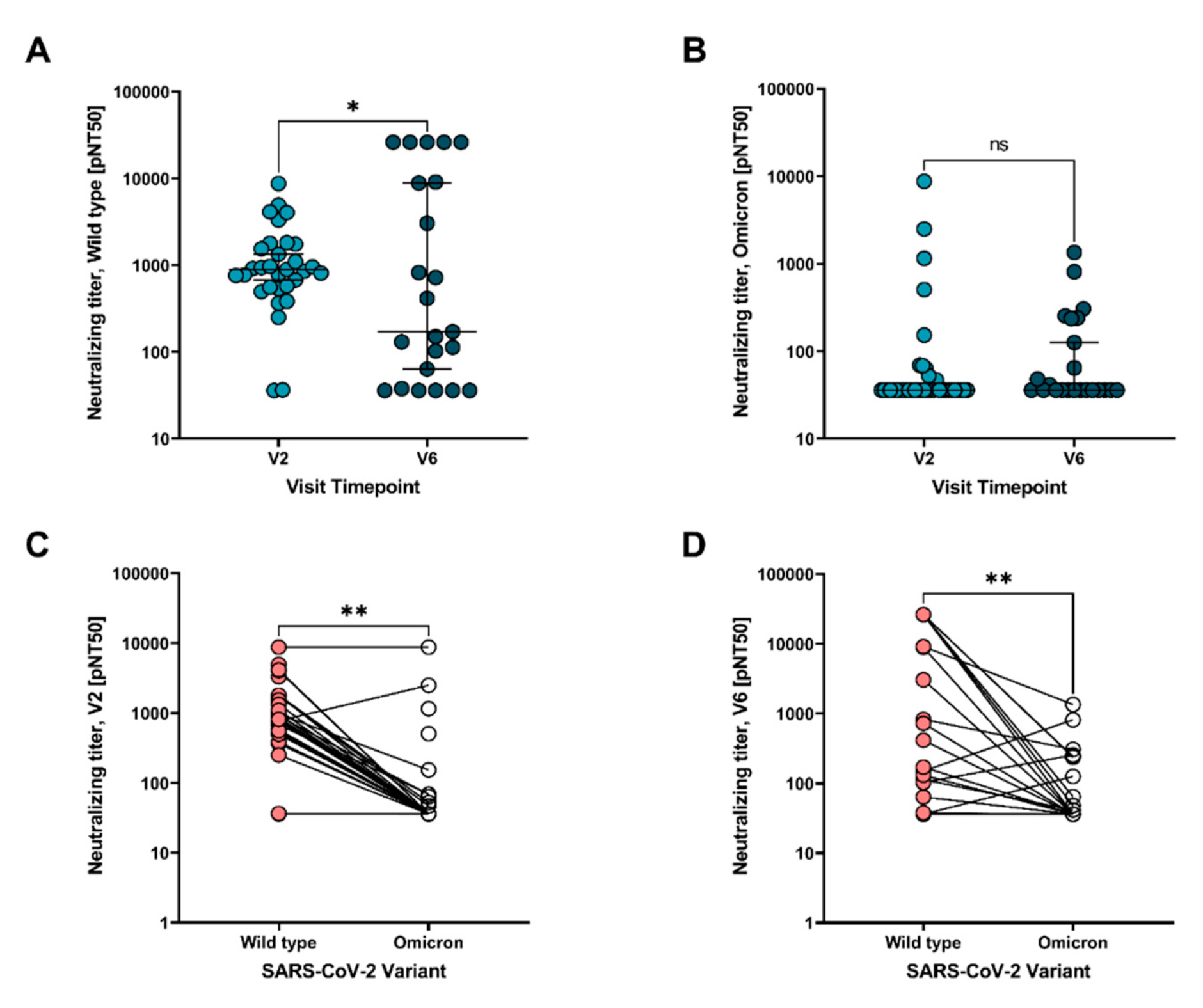
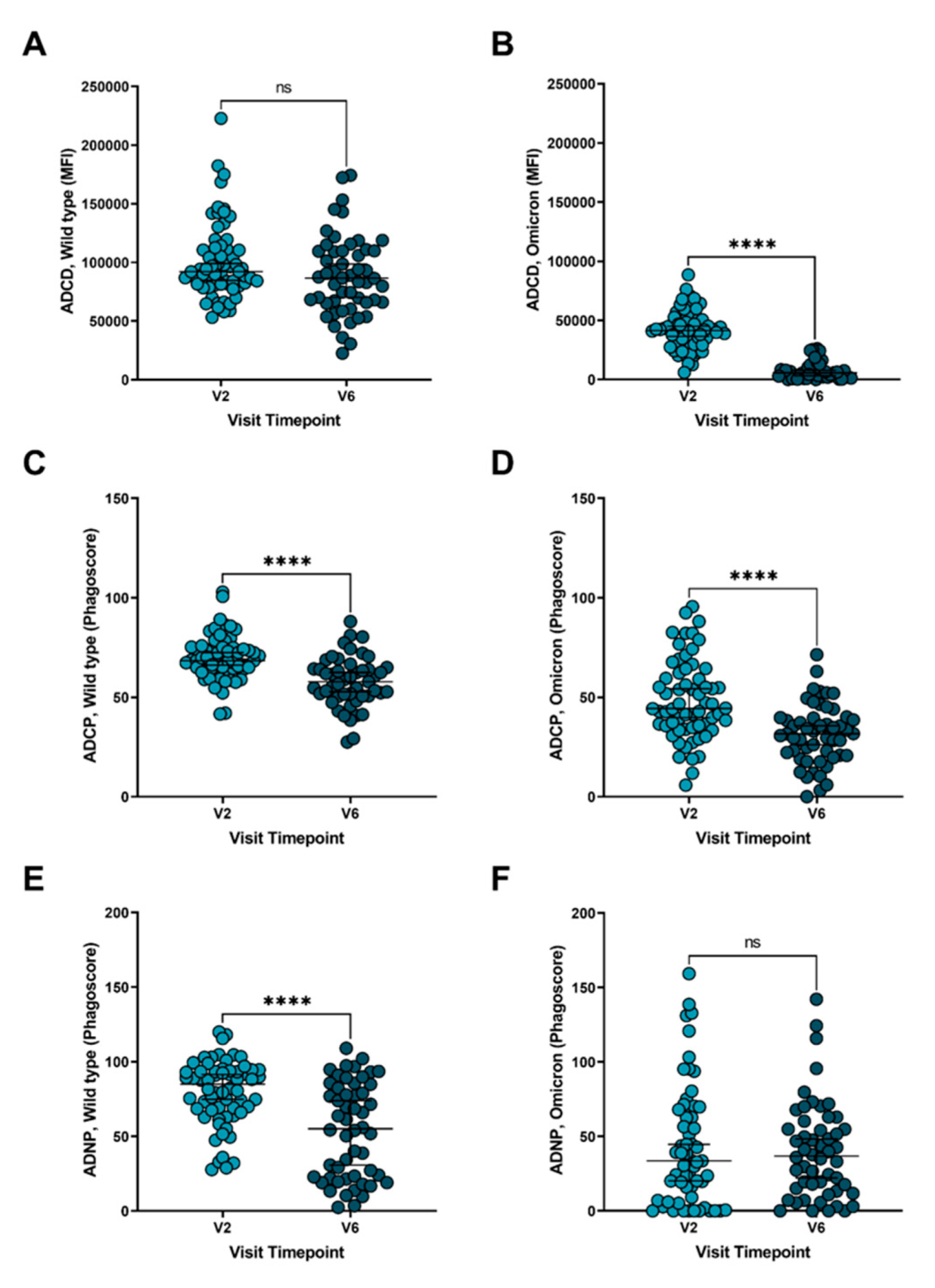
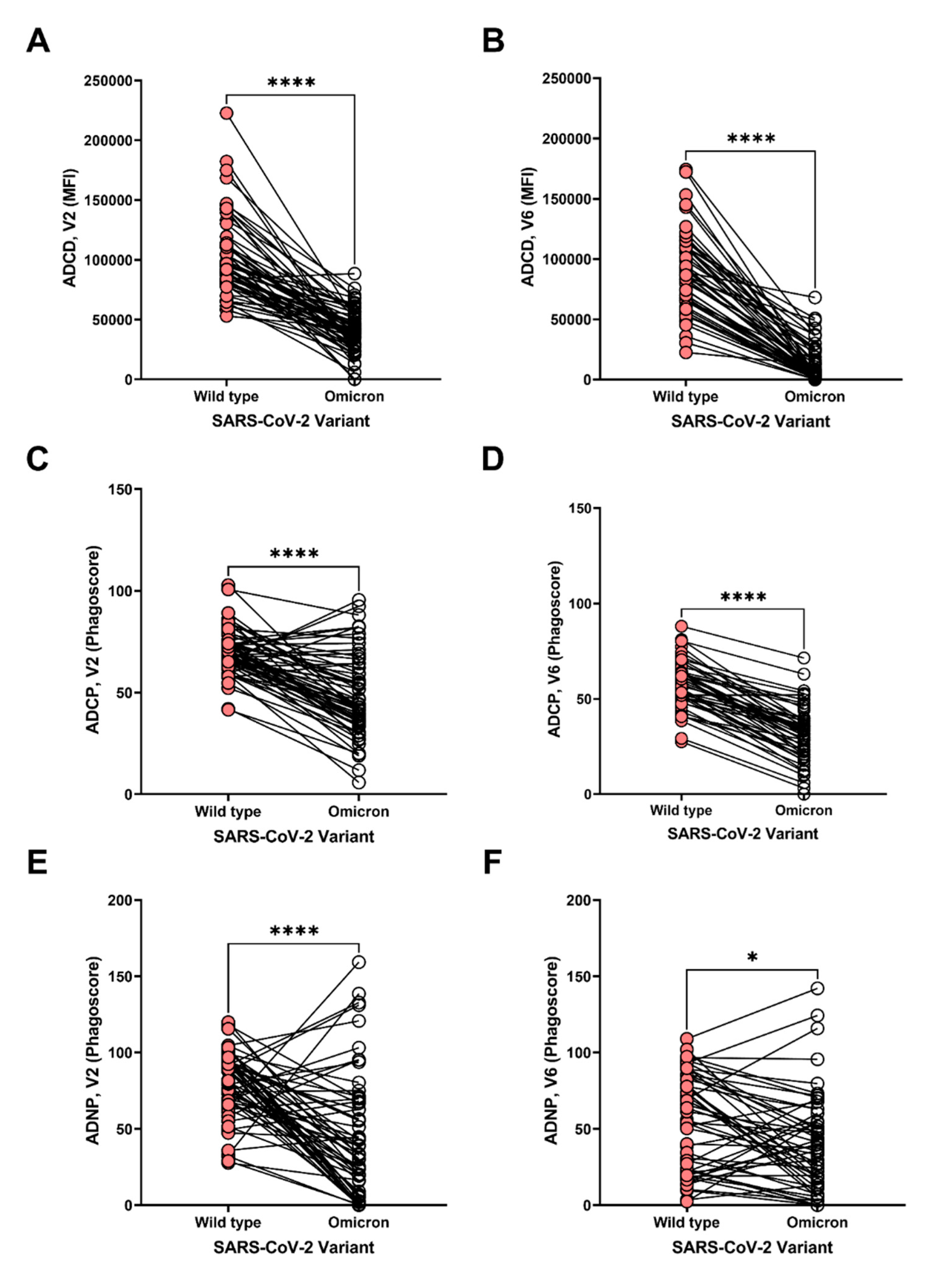
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Lipsitch, M.; Krammer, F.; Regev-Yochay, G.; Lustig, Y.; Balicer, R.D. SARS-CoV-2 breakthrough infections in vaccinated individuals: Measurement, causes and impact. Nat. Rev. Immunol. 2021, 22, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://services.aap.org/en/pages/2019-novel-coronavirus-covid-19-infections/children-and-covid-19-state-level-data-report/ (accessed on 12 February 2022).
- Available online: https://data.unicef.org/topic/child-survival/covid-19/ (accessed on 12 February 2022).
- Available online: https://www.cdc.gov/mis-c/hcp/ (accessed on 12 February 2022).
- Stephenson, T.; Shafran, R.; De Stavola, B.; Rojas, N.; Aiano, F.; Amin-Chowdhury, Z.; McOwat, K.; Simmons, R.; Zavala, M.; Consortium, C.; et al. Long COVID and the mental and physical health of children and young people: National matched cohort study protocol (the CloCk study). BMJ Open 2021, 11, e052838. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, L.D.; Newhams, M.M.; Olson, S.M.; Halasa, N.B.; Price, A.M.; Boom, J.A.; Sahni, L.C.; Kamidani, S.; Tarquinio, K.M.; Maddux, A.B.; et al. Effectiveness of BNT162b2 (Pfizer-BioNTech) mRNA Vaccination Against Multisystem Inflammatory Syndrome in Children Among Persons Aged 12–18 Years-United States, July–December 2021. Morb. Mortal. Wkly. Rep. 2022, 71, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Frenck, R.W., Jr.; Klein, N.P.; Kitchin, N.; Gurtman, A.; Absalon, J.; Lockhart, S.; Perez, J.L.; Walter, E.B.; Senders, S.; Bailey, R.; et al. Safety, Immunogenicity, and Efficacy of the BNT162b2 COVID-19 Vaccine in Adolescents. N. Engl. J. Med. 2021, 385, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Fowlkes, A.L.; Yoon, S.K.; Lutrick, K.; Gwynn, L.; Burns, J.; Grant, L.; Phillips, A.L.; Ellingson, K.; Ferraris, M.V.; LeClair, L.B.; et al. Effectiveness of 2-Dose BNT162b2 (Pfizer BioNTech) mRNA Vaccine in Preventing SARS-CoV-2 Infection among Children Aged 5–11 Years and Adolescents Aged 12–15 Years—PROTECT Cohort, July 2021–February 2022. Morb. Mortal. Wkly. Rep. 2022, 71, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.pfizer.com/news/press-release/press-release-detail/pfizer-and-biontech-provide-update-rolling-submission (accessed on 8 March 2022).
- Klein, N.P.; Stockwell, M.S.; Demarco, M.; Gaglani, M.; Kharbanda, A.B.; Irving, S.A.; Rao, S.; Grannis, S.J.; Dascomb, K.; Murthy, K.; et al. Effectiveness of COVID-19 Pfizer-BioNTech BNT162b2 mRNA Vaccination in Preventing COVID-19-Associated Emergency Department and Urgent Care Encounters and Hospitalizations Among Nonimmunocompromised Children and Adolescents Aged 5–17 Years-VISION Network, 10 States, April 2021–January 2022. Morb. Mortal. Wkly. Rep. 2022, 71, 352–358. [Google Scholar] [CrossRef]
- Lima, R.; Gootkind, E.F.; De la Flor, D.; Yockey, L.J.; Bordt, E.A.; D’Avino, P.; Ning, S.; Heath, K.; Harding, K.; Zois, J.; et al. Establishment of a pediatric COVID-19 biorepository: Unique considerations and opportunities for studying the impact of the COVID-19 pandemic on children. BMC Med. Res. Methodol. 2020, 20, 228. [Google Scholar] [CrossRef] [PubMed]
- Yonker, L.M.; Neilan, A.M.; Bartsch, Y.; Patel, A.B.; Regan, J.; Arya, P.; Gootkind, E.; Park, G.; Hardcastle, M.; St John, A.; et al. Pediatric SARS-CoV-2: Clinical Presentation, Infectivity, and Immune Responses. J. Pediatr. 2020, 227, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Beltran, W.F.; Lam, E.C.; St Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021, 184, 2372–2383. [Google Scholar] [CrossRef] [PubMed]
- Fischinger, S.; Fallon, J.K.; Michell, A.R.; Broge, T.; Suscovich, T.J.; Streeck, H.; Alter, G. A high-throughput, bead-based, antigen-specific assay to assess the ability of antibodies to induce complement activation. J. Immunol. Methods 2019, 473, 112630. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, M.E.; Moldt, B.; Wyatt, R.T.; Dugast, A.S.; McAndrew, E.; Tsoukas, S.; Jost, S.; Berger, C.T.; Sciaranghella, G.; Liu, Q.; et al. A robust, high-throughput assay to determine the phagocytic activity of clinical antibody samples. J. Immunol. Methods 2011, 366, 8–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karsten, C.B.; Mehta, N.; Shin, S.A.; Diefenbach, T.J.; Slein, M.D.; Karpinski, W.; Irvine, E.B.; Broge, T.; Suscovich, T.J.; Alter, G. A versatile high-throughput assay to characterize antibody-mediated neutrophil phagocytosis. J. Immunol. Methods 2019, 471, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Kurtovic, L.; Beeson, J.G. Complement Factors in COVID-19 Therapeutics and Vaccines. Trends Immunol. 2021, 42, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Carreño, J.M.; Alshammary, H.; Tcheou, J.; Singh, G.; Raskin, A.J.; Kawabata, H.; Sominsky, L.A.; Clark, J.J.; Adelsberg, D.C.; Bielak, D.A.; et al. Activity of convalescent and vaccine serum against SARS-CoV-2 Omicron. Nature 2021, 602, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, Y.C.; Wang, C.; Zohar, T.; Fischinger, S.; Atyeo, C.; Burke, J.S.; Kang, J.; Edlow, A.G.; Fasano, A.; Baden, L.R.; et al. Humoral signatures of protective and pathological SARS-CoV-2 infection in children. Nat. Med. 2021, 27, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Canaday, D.H.; Oyebanji, O.A.; Keresztesy, D.; Payne, M.; Wilk, D.; Carias, L.; Aung, H.; Denis, K.S.; Lam, E.C.; Rowley, C.F.; et al. Significant Reduction in Vaccine-Induced Antibody Levels and Neutralization Activity among Healthcare Workers and Nursing Home Residents 6 Months Following Coronavirus Disease 2019 BNT162b2 mRNA Vaccination. Clin. Infect. Dis. 2021, ciab963. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.cdc.gov/coronavirus/2019-ncov/vaccines/booster-shot.html (accessed on 13 March 2022).
- Accorsi, E.K.; Britton, A.; Fleming-Dutra, K.E.; Smith, Z.R.; Shang, N.; Derado, G.; Miller, J.; Schrag, S.J.; Verani, J.R. Association Between 3 Doses of mRNA COVID-19 Vaccine and Symptomatic Infection Caused by the SARS-CoV-2 Omicron and Delta Variants. JAMA 2022, 327, 639–651. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Characteristics | Cohort (n = 77) |
|---|---|
| Age at Enrollment, mean (SD) | 14 (1.8) |
| Male, number (%) | 41 (53) |
| Hispanic, number (%) | 15 (19) |
| Race, number (%) | |
| White | 53 (69) |
| Black | 2 (3) |
| Asian | 1 (1) |
| Other | 16 (21) |
| Unknown | 5 (6) |
| Time Since Dose 1 and V1 Draw, average days (SD) | 18 (3) |
| Time Since Dose 2 and V2 Draw, average days (SD) | 18 (5) |
| Time Since Dose 2 and V6 Draw, average months (SD) | 6.7 (0.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burns, M.D.; Boribong, B.P.; Bartsch, Y.C.; Loiselle, M.; St. Denis, K.J.; Sheehan, M.L.; Chen, J.W.; Davis, J.P.; Lima, R.; Edlow, A.G.; et al. Durability and Cross-Reactivity of SARS-CoV-2 mRNA Vaccine in Adolescent Children. Vaccines 2022, 10, 492. https://doi.org/10.3390/vaccines10040492
Burns MD, Boribong BP, Bartsch YC, Loiselle M, St. Denis KJ, Sheehan ML, Chen JW, Davis JP, Lima R, Edlow AG, et al. Durability and Cross-Reactivity of SARS-CoV-2 mRNA Vaccine in Adolescent Children. Vaccines. 2022; 10(4):492. https://doi.org/10.3390/vaccines10040492
Chicago/Turabian StyleBurns, Madeleine D., Brittany P. Boribong, Yannic C. Bartsch, Maggie Loiselle, Kerri J. St. Denis, Maegan L. Sheehan, Jessica W. Chen, Jameson P. Davis, Rosiane Lima, Andrea G. Edlow, and et al. 2022. "Durability and Cross-Reactivity of SARS-CoV-2 mRNA Vaccine in Adolescent Children" Vaccines 10, no. 4: 492. https://doi.org/10.3390/vaccines10040492
APA StyleBurns, M. D., Boribong, B. P., Bartsch, Y. C., Loiselle, M., St. Denis, K. J., Sheehan, M. L., Chen, J. W., Davis, J. P., Lima, R., Edlow, A. G., Fasano, A., Balazs, A. B., Alter, G., & Yonker, L. M. (2022). Durability and Cross-Reactivity of SARS-CoV-2 mRNA Vaccine in Adolescent Children. Vaccines, 10(4), 492. https://doi.org/10.3390/vaccines10040492