
Evaluation of a Lipopolysaccharide and Resiquimod Combination as an Adjuvant with Inactivated Newcastle Disease Virus Vaccine in Chickens

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Birds and TLR Agonists
2.2. Bulk Production and Titration of NDV
2.3. Evaluation of Adjuvant Activity of TLR Agonists’ Combination
2.3.1. Preparation of Inactivated NDV Vaccine
2.3.2. Experimental Design
2.3.3. Evaluation of Humoral Immune Response
2.3.4. Evaluation of Cellular Immune Response
Lymphocyte Transformation Test (LTT)
Flow Cytometry Analysis
2.4. Challenge Study
2.5. Viral Shedding
2.6. Statistical Analysis
3. Results
3.1. Preparation of Inactivated NDV Vaccine
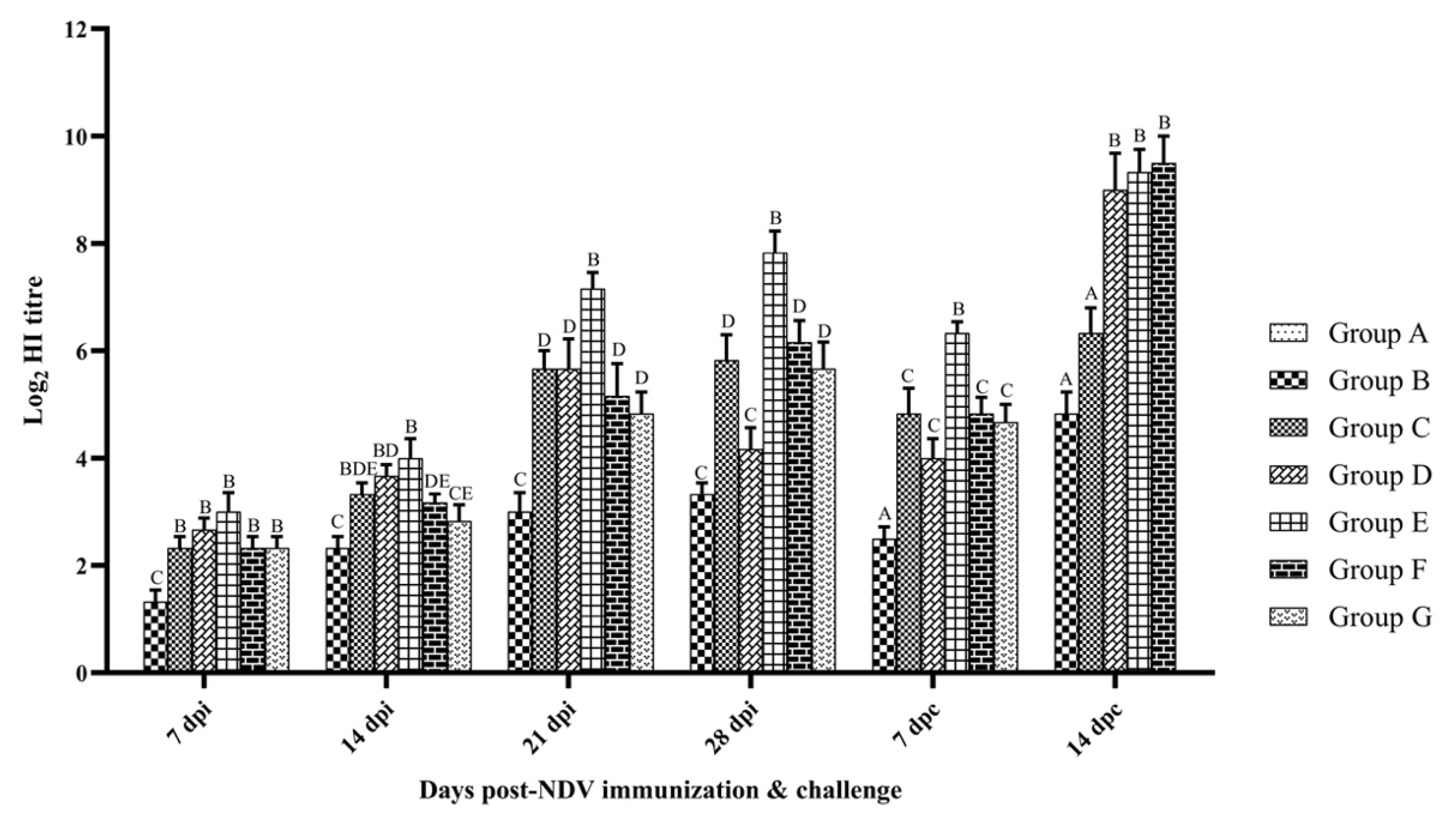
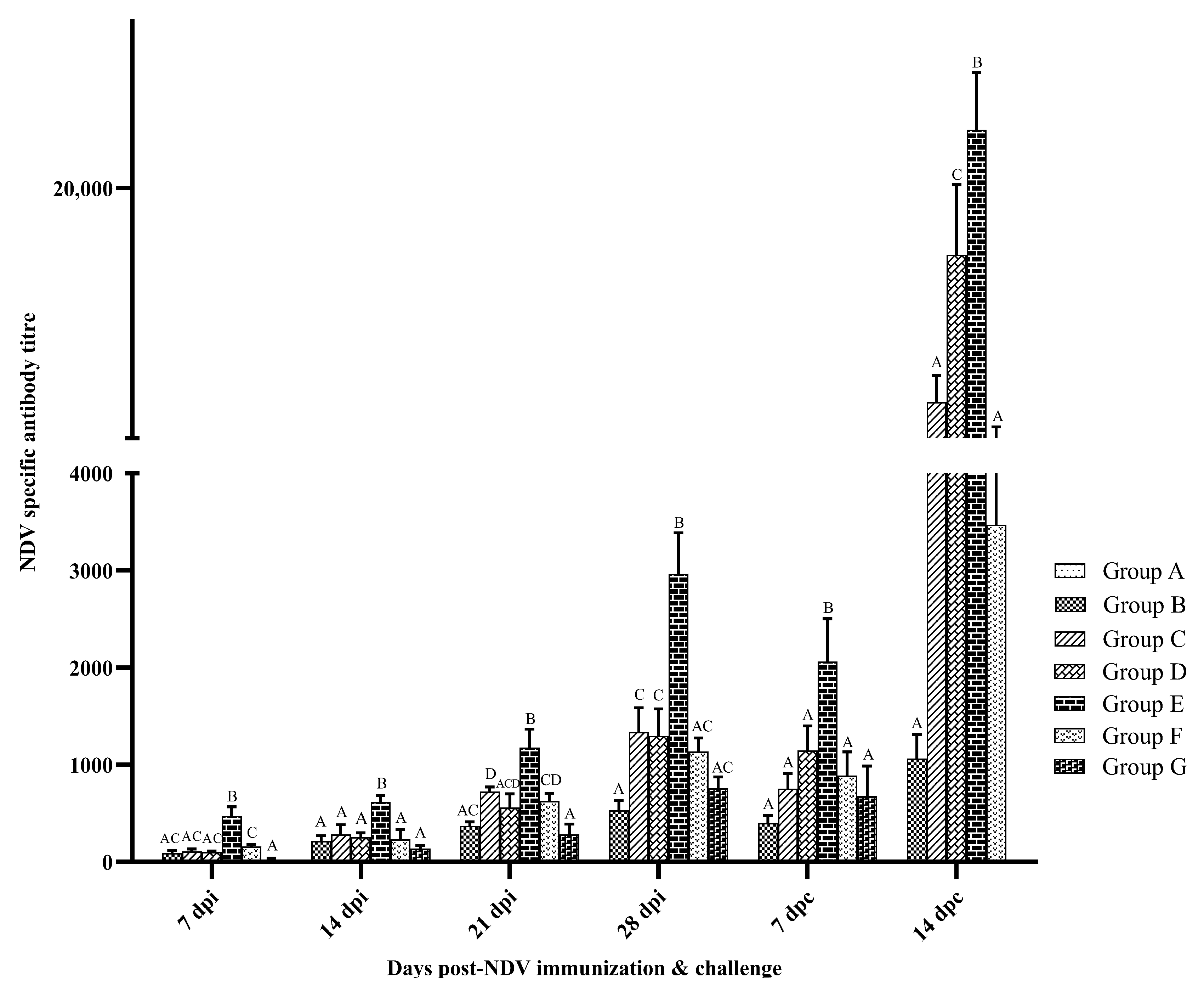
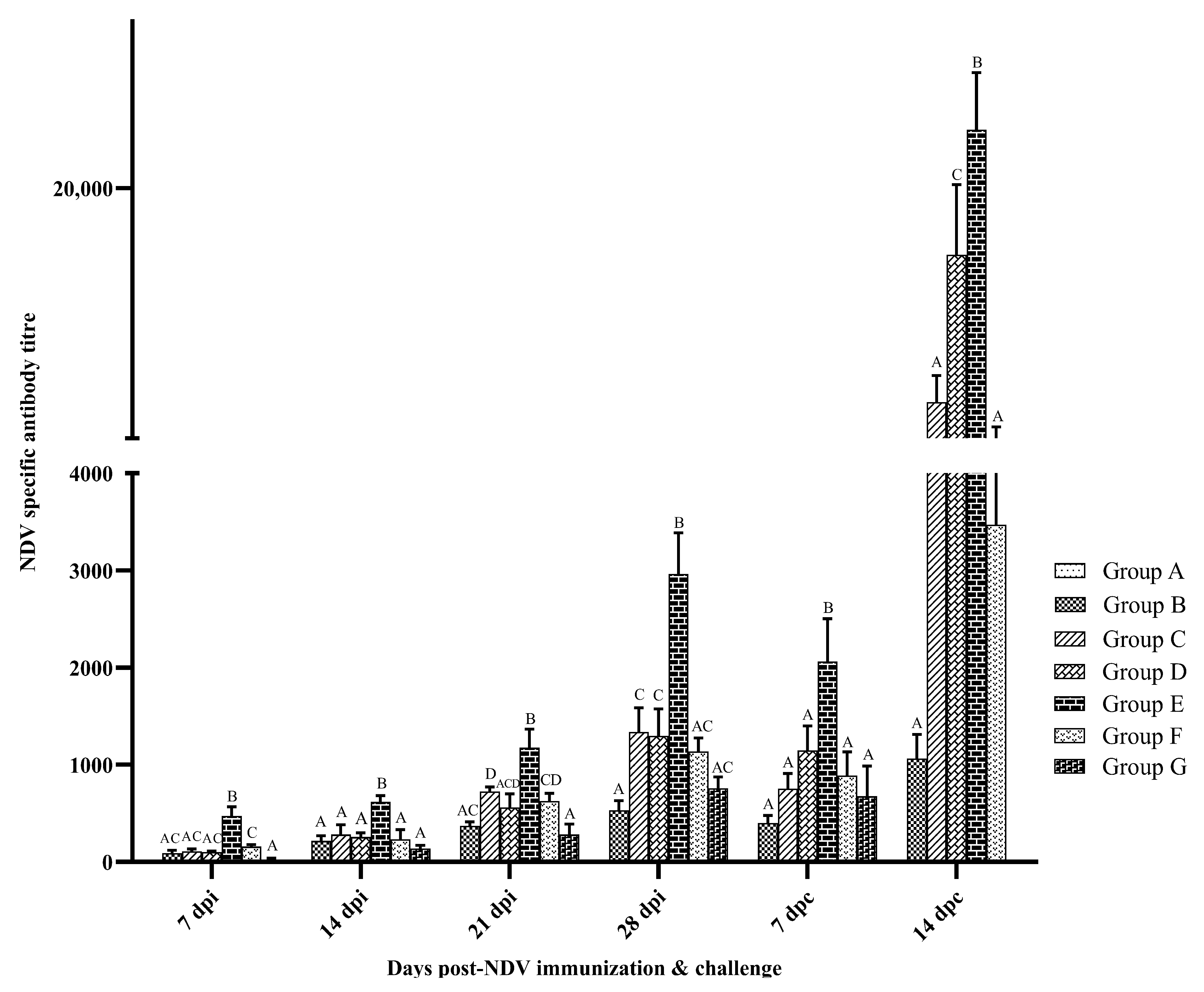
3.2. Humoral Immune Response
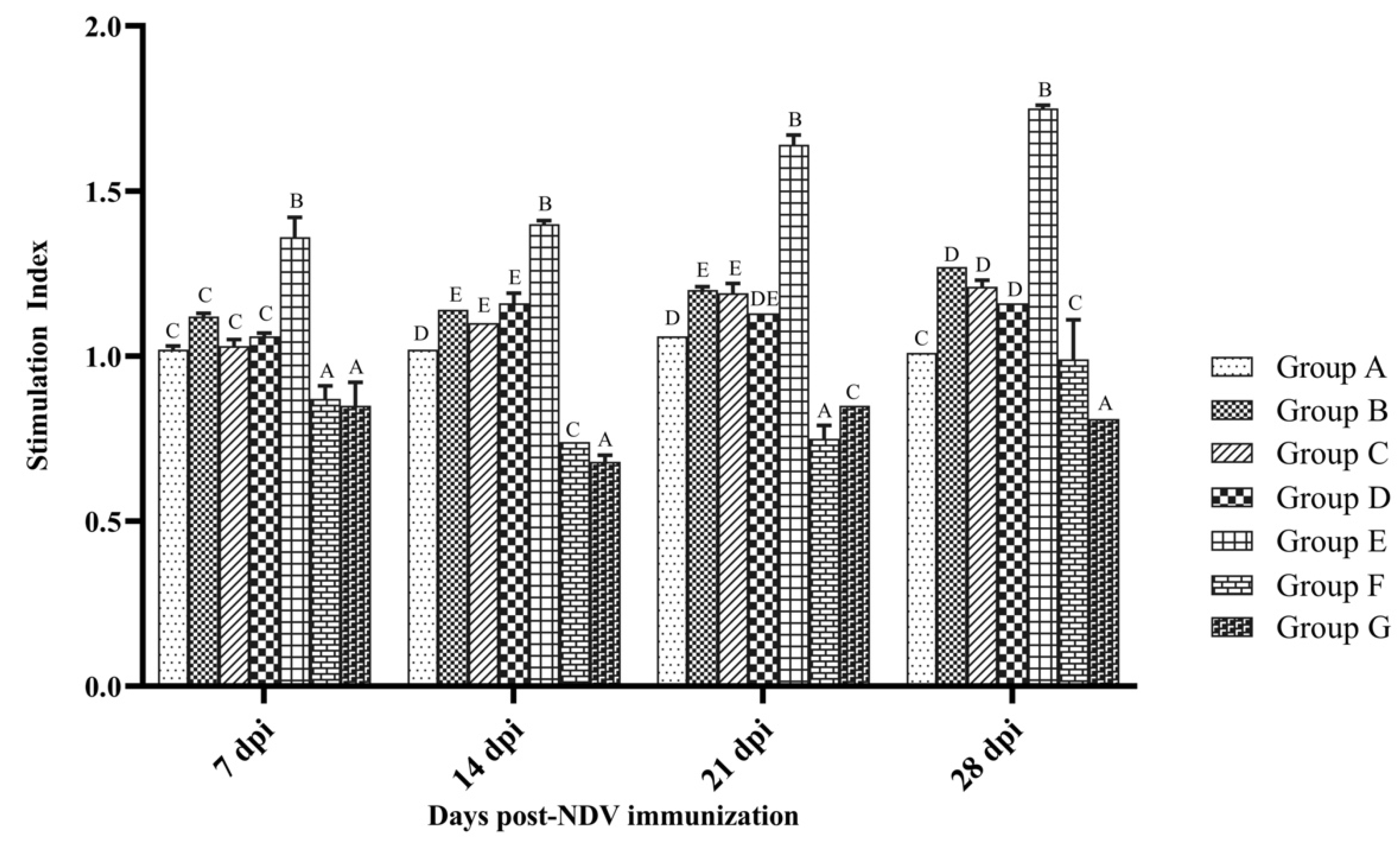
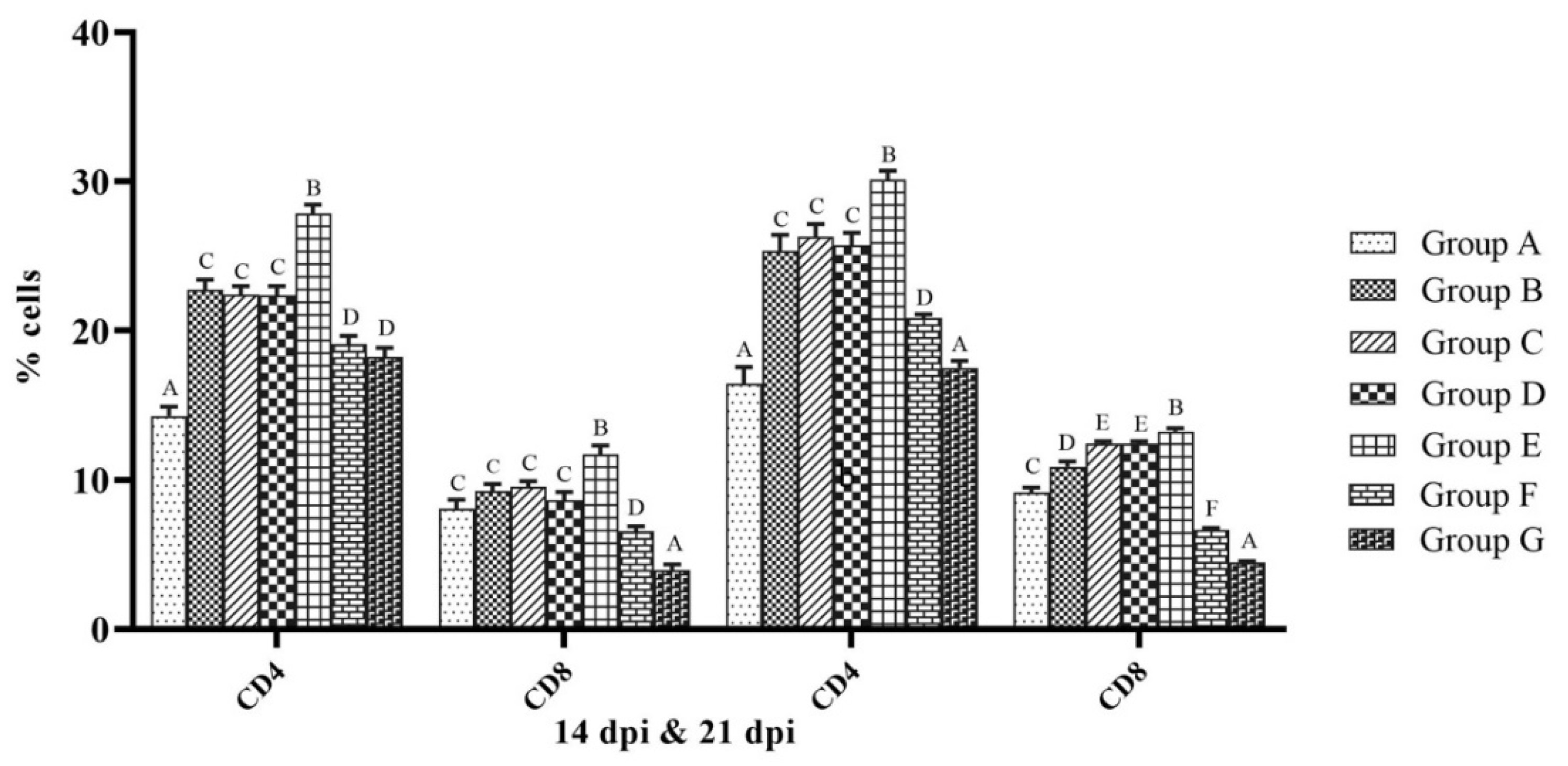
3.3. Cellular Immune Response
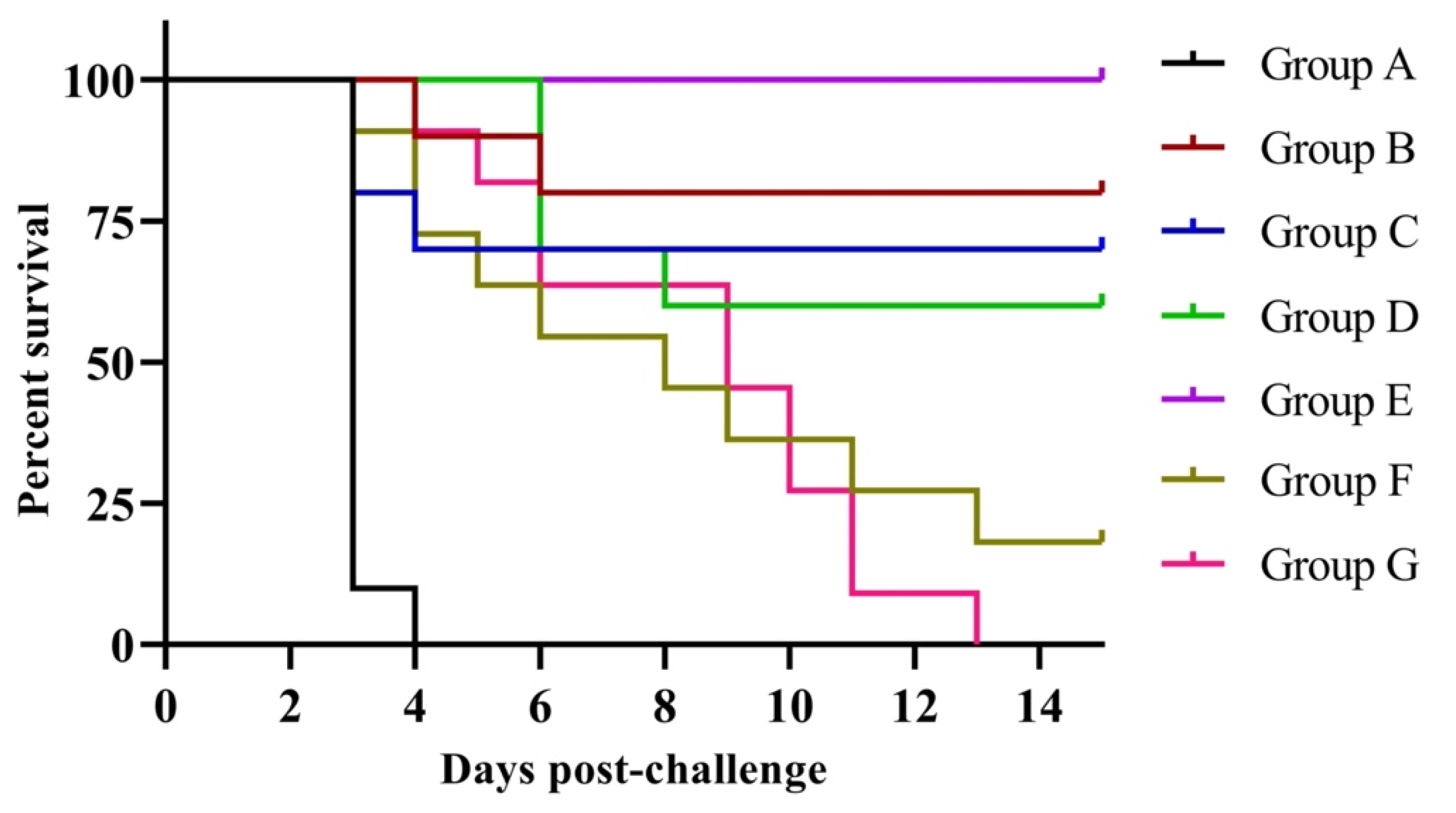
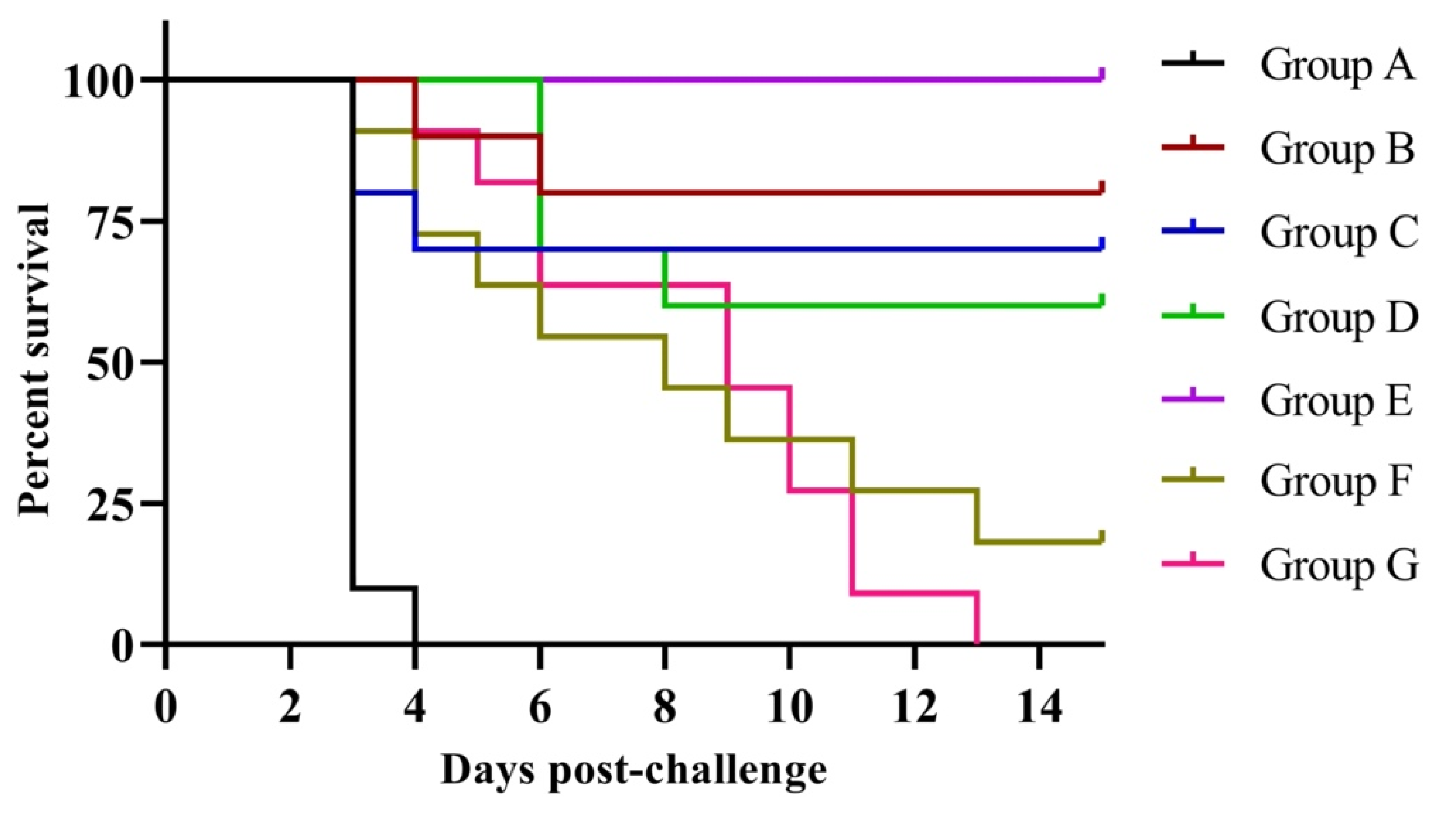
3.4. Protection against Clinical Signs, Mortality and Virus Shedding
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group $ (n = 10 or 11/Group) | Survived | Dead | Protection (%) |
|---|---|---|---|
| A | 0 | 10 * | 0 |
| B | 7 | 3 | 80 |
| C | 8 | 2 | 70 |
| D | 6 | 4 | 60 |
| E | 10 | 0 | 100 |
| F (11) | 2 | 9 | 18.18 |
| G (11) | 0 | 11 | 0 |
References
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Werling, D.; Jungi, T.W. Toll-like receptors linking innate and adaptive immune response. Vet. Immunol. Immunopathol. 2003, 91, 1–12. [Google Scholar] [CrossRef]
- Hou, B.; Reizis, B.; DeFranco, A.L. Toll-like receptors activate innate and adaptive immunity by using dendritic cell-intrinsic and-extrinsic mechanisms. Immunity 2008, 29, 272–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Cheng, A.; Wang, M. Innate sensing of viruses by pattern recognition receptors in birds. Vet. Res. 2013, 44, 82. [Google Scholar] [CrossRef] [Green Version]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; Huffel, C.V.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef] [Green Version]
- Beutler, B. TLR4 as the mammalian endotoxin sensor. Curr. Top. Microbiol. Immunol. 2002, 270, 109–120. [Google Scholar]
- Thoelen, S.; De Clercq, N.; Tornieporth, N. A prophylactic hepatitis B vaccine with a novel adjuvant system. Vaccine 2001, 19, 2400–2403. [Google Scholar] [CrossRef]
- Harper, D.M.; Franco, E.L.; Wheeler, C.; Ferris, D.G.; Jenkins, D.; Schuind, A.; Zahaf, T.; Innis, B.; Naud, P.; De Carvalho, N.S.; et al. Efficacy of a bivalent L1 virus-like particle vaccine in prevention of infection with human papillomavirus types 16 and 18 in young women: A randomised controlled trial. Lancet 2004, 364, 1757–1765. [Google Scholar] [CrossRef]
- Tseng, L.P.; Chiou, C.J.; Chen, C.C.; Deng, M.C.; Chung, T.W.; Huang, Y.Y.; Liu, D.Z. Effect of lipopolysaccharide on intranasal administration of liposomal Newcastle disease virus vaccine to SPF chickens. Vet. Immunol. Immunopathol. 2009, 131, 285–289. [Google Scholar] [CrossRef]
- Lee, J.; Chuang, T.H.; Redecke, V.; She, L.; Pitha, P.M.; Carson, D.A.; Raz, E.; Cottam, H.B. Molecular basis for the immunostimulatory activity of guanine nucleoside analogs: Activation of Toll-like receptor 7. Proc. Natl. Acad. Sci. USA 2003, 100, 6646–6651. [Google Scholar] [CrossRef] [Green Version]
- Jurk, M.; Kritzler, A.; Schulte, B.; Tluk, S.; Schetter, C.; Krieg, A.M.; Vollmer, J. Modulating responsiveness of human TLR7 and 8 to small molecule ligands with T-rich phosphorothiate oligodeoxynucleotides. Eur. J. Immunol. 2006, 36, 1815–1826. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ohto, U.; Shibata, T.; Krayukhina, E.; Taoka, M.; Yamauchi, Y.; Tanji, H.; Isobe, T.; Uchiyama, S.; Miyake, K.; et al. Structural analysis reveals that Toll-like receptor 7 is a dual receptor for guanosine and single-stranded RNA. Immunity 2016, 45, 737–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachan, S.; Ramakrishnan, S.; Annamalai, A.; Sharma, B.K.; Malik, H.; Saravanan, B.C.; Jain, L.; Saxena, M.; Kumar, A.; Krishnaswamy, N. Adjuvant potential of resiquimod with inactivated Newcastle disease vaccine and its mechanism of action in chicken. Vaccine 2015, 33, 4526–4532. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Kim, Y.J. Signaling pathways downstream of pattern-recognition receptors and their cross talk. Annu. Rev. Biochem. 2007, 76, 447–480. [Google Scholar] [CrossRef]
- Tom, J.K.; Albin, T.J.; Manna, S.; Moser, B.A.; Steinhardt, R.C.; Esser-Kahn, A.P. Applications of immunomodulatory immune synergies to adjuvant discovery and vaccine development. Trends Biotechnol. 2019, 37, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, S.; Annamalai, A.; Sachan, S.; Kumar, A.; Sharma, B.K.; Govindaraj, E.; Chellappa, M.M.; Dey, S.; Krishnaswamy, N. Synergy of lipopolysaccharide and resiquimod on type I interferon, pro-inflammatory cytokine, Th1 and Th2 response in chicken peripheral blood mononuclear cells. Mol. Immunol. 2015, 64, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.K.; Mickelson, D.J.; Solberg, J.C.; Lipson, K.E.; Inglefield, J.R.; Alkan, S.S. TLR–TLR cross talk in human PBMC resulting in synergistic and antagonistic regulation of type-1 and 2 interferons, IL-12 and TNF-α. Int. Immunopharmacol. 2007, 7, 1111–1121. [Google Scholar] [CrossRef]
- Fischetti, L.; Zhong, Z.; Pinder, C.L.; Tregoning, J.S.; Shattock, R.J. The synergistic effects of combining TLR ligand based adjuvants on the cytokine response are dependent upon p38/JNK signalling. Cytokine 2017, 99, 287–296. [Google Scholar] [CrossRef]
- Surendran, N.; Simmons, A.; Pichichero, M.E. TLR agonist combinations that stimulate Th type I polarizing responses from human neonates. Innate Immun. 2018, 24, 240–251. [Google Scholar] [CrossRef]
- Goff, P.H.; Hayashi, T.; Martínez-Gil, L.; Corr, M.; Crain, B.; Yao, S.; Cottam, H.B.; Chan, M.; Ramos, I.; Eggink, D.; et al. Synthetic Toll-like receptor 4 (TLR4) and TLR7 ligands as influenza virus vaccine adjuvants induce rapid, sustained, and broadly protective responses. J. Virol. 2015, 89, 3221–3235. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.X.; Brown, I.H.; Choi, K.S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: Mammals, Birds and Bees, Biological Standards Commission, 7th ed.; World Organization for Animal Health (OIE): Paris, France, 2012. [Google Scholar]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival: Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods. 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Gupta, S.K.; Deb, R.; Dey, S.; Chellappa, M.M. Toll-like receptor-based adjuvants: Enhancing the immune response to vaccines against infectious diseases of chicken. Expert Rev. Vaccines. 2014, 13, 909–925. [Google Scholar] [CrossRef] [PubMed]
- Bhadouriya, S.; Sharma, B.K.; Kakker, N.K.; Chhabra, R. Toll like receptors and cytokines as immunostimulatory adjuvants in poultry vaccines: Current status and future trends. World’s Poult. Sci. J. 2019, 75, 417–428. [Google Scholar] [CrossRef]
- He, H.; Genovese, K.J.; Nisbet, D.J.; Kogut, M.H. Synergy of CpG oligodeoxynucleotide and double-stranded RNA (poly I: C) on nitric oxide induction in chicken peripheral blood monocytes. Mol. Immunol. 2007, 44, 3234–3242. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Genovese, K.J.; Kogut, M.H. Modulation of chicken macrophage effector function by TH1/TH2 cytokines. Cytokine 2011, 53, 363–369. [Google Scholar] [CrossRef]
- St Paul, M.; Paolucci, S.; Barjesteh, N.; Wood, R.D.; Sharif, S. Chicken erythrocytes respond to Toll-like receptor ligands by up-regulating cytokine transcripts. Res. Vet. Sci. 2013, 95, 87–91. [Google Scholar] [CrossRef]
- Gupta, S.K.; Deb, R.; Gaikwad, S.; Saravanan, R.; Mohan, C.M.; Dey, S. Recombinant flagellin and its cross-talk with lipopolysaccharide–Effect on pooled chicken peripheral blood mononuclear cells. Res. Vet. Sci. 2013, 95, 30–935. [Google Scholar] [CrossRef]
- Goff, P.H.; Hayashi, T.; He, W.; Yao, S.; Cottam, H.B.; Tan, G.S.; Crain, B.; Krammer, F.; Messer, K.; Pu, M.; et al. Synthetic toll-like receptor 4 (TLR4) and TLR7 ligands work additively via MyD88 to induce protective antiviral immunity in mice. J. Virol. 2017, 91, e01050-17. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, D.L.; Maraqa, A.D. Protective immunity against Newcastle disease: The role of antibodies specific to Newcastle disease virus polypeptides. Avian. Dis. 2000, 44, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Schwenk, R.; DeBot, M.; Porter, M.; Nikki, J.; Rein, L.; Spaccapelo, R.; Crisanti, A.; Wightman, P.D.; Ockenhouse, C.F.; Dutta, S. IgG2 antibodies against a clinical grade Plasmodium falciparum CSP vaccine antigen associate with protection against transgenic sporozoite challenge in mice. PLoS ONE 2014, 9, 111020. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.L.; Maraqa, A.D. Protective immunity against Newcastle disease: The role of cell-mediated immunity. Avian. Dis. 2000, 44, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Erf, G.F. Cell-mediated immunity in poultry. Poult. Sci. 2004, 83, 580–590. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Afonso, C.L.; Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 2013, 41, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.J.; Steinbach, S.; Clifford, D.; Baldwin, S.L.; Ireton, G.C.; Coler, R.N.; Reed, S.G.; Vordermeier, H.M. Immunisation with ID83 fusion protein induces antigen-specific cell mediated and humoral immune responses in cattle. Vaccine 2013, 31, 5250–5255. [Google Scholar] [CrossRef] [Green Version]
- McKay, P.F.; King, D.F.; Mann, J.F.; Barinaga, G.; Carter, D.; Shattock, R.J. TLR4 and TLR7/8 adjuvant combinations generate different vaccine antigen-specific immune outcomes in minipigs when administered via the ID or IN routes. PLoS ONE 2016, 11, e0148984. [Google Scholar] [CrossRef] [Green Version]
- Rostamian, M.; Niknam, H.M. Evaluation of the adjuvant effect of agonists of toll-like receptor 4 and 7/8 in a vaccine against leishmaniasis in BALB/c mice. Mol. Immunol. 2017, 91, 202–208. [Google Scholar] [CrossRef]
- Tacken, P.J.; Zeelenberg, I.S.; Cruz, L.J.; van Hout-Kuijer, M.A.; van de Glind, G.; Fokkink, R.G.; Lambeck, A.J.; Figdor, C.G. Targeted delivery of TLR ligands to human and mouse dendritic cells strongly enhances adjuvanticity. Blood 2011, 118, 6836–6844. [Google Scholar] [CrossRef] [Green Version]
- Fox, C.B.; Sivananthan, S.J.; Duthie, M.S.; Vergara, J.; Guderian, J.A.; Moon, E.; Coblentz, D.; Reed, S.G.; Carter, D. A nanoliposome delivery system to synergistically trigger TLR4 AND TLR7. J. Nanobiotechnol. 2014, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Ilyinskii, P.O.; Roy, C.J.; O’Neil, C.P.; Browning, E.A.; Pittet, L.A.; Altreuter, D.H.; Alexis, F.; Tonti, E.; Shi, J.; Basto, P.A.; et al. Adjuvant-carrying synthetic vaccine particles augment the immune response to encapsulated antigen and exhibit strong local immune activation without inducing systemic cytokine release. Vaccine 2014, 32, 2882–2895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madan-Lala, R.; Pradhan, P.; Roy, K. Combinatorial delivery of dual and triple TLR agonists via polymeric pathogen-like particles synergistically enhances innate and adaptive immune responses. Sci. Rep. 2017, 7, 2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Short, K.K.; Miller, S.M.; Walsh, L.; Cybulski, V.; Bazin, H.; Evans, J.T.; Burkhart, D. Co-encapsulation of synthetic lipidated TLR4 and TLR7/8 agonists in the liposomal bilayer results in a rapid, synergistic enhancement of vaccine-mediated humoral immunity. J. Control. Release 2019, 315, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Sato-Kaneko, F.; Yao, S.; Lao, F.S.; Shpigelman, J.; Messer, K.; Pu, M.; Shukla, N.M.; Cottam, H.B.; Chan, M.; Chu, P.J.; et al. A novel synthetic dual agonistic liposomal TLR4/7 adjuvant promotes broad immune responses in an influenza vaccine with minimal reactogenicity. Front. Immunol. 2020, 11, 1207. [Google Scholar] [CrossRef]
- Jayawardane, G.W.L.; Spradbrow, P.B. Mucosal immunity in chickens vaccinated with the V4 strain of Newcastle disease virus. Vet. Microbiol. 1995, 46, 69–77. [Google Scholar] [CrossRef]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar] [CrossRef]
- Miller, P.J.; Estevez, C.; Yu, Q.; Suarez, D.L.; King, D.J. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis. 2009, 53, 39–49. [Google Scholar] [CrossRef]



| Groups (n=10 or 11/Group) * | Vaccine (Dose/Bird) | TLR Agonist(s) (Dose/Bird) |
|---|---|---|
| Control (A) | PBS | - |
| Commercial vaccine (B) | Commercial oil adjuvanted inactivated NDV vaccine (recommended dose) | - |
| Vaccine alone (C) | Inactivated NDV vaccine (≥107 EID50) | - |
| Vaccine + TLR4 agonist (D) | Inactivated NDV vaccine (≥107 EID50) | LPS (50 µg) |
| Vaccine + TLR7 agonist (E) | Inactivated NDV vaccine (≥107 EID50) | R-848 (50 µg) |
| Vaccine + TLR4 and TLR7 agonists in low dose (F) | Inactivated NDV vaccine (≥107 EID50) | LPS (25 µg) + R-848 (25 µg) |
| Vaccine + TLR4 and TLR7 agonists in high dose (G) | Inactivated NDV vaccine (≥107 EID50) | LPS (50 µg) + R-848 (50 µg) |
| Group $ (n = 6/Group) | Days Post-Challenge | ||
|---|---|---|---|
| 4 | 7 | 14 | |
| A | Died | Died | Died |
| B | 100 (6) | 100 (6) | 100 (6) |
| C | 100 (6) | 100 (6) | 66.67 (4) |
| D | 66.67 (4) | 66.67 (4) | 33.33 (2) |
| E | 0 (0) | 0 (0) | 0 (0) |
| F | 66.67 (4) | 66.67 (4) | 33.33 (2) * |
| G | 66.67 (4) | 66.67 (4) | Died |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, B.K.; Ramakrishnan, S.; Kaliappan, A.; Singh, M.; Kumar, A.; Dandapat, S.; Dey, S.; Chellappa, M.M. Evaluation of a Lipopolysaccharide and Resiquimod Combination as an Adjuvant with Inactivated Newcastle Disease Virus Vaccine in Chickens. Vaccines 2022, 10, 894. https://doi.org/10.3390/vaccines10060894
Sharma BK, Ramakrishnan S, Kaliappan A, Singh M, Kumar A, Dandapat S, Dey S, Chellappa MM. Evaluation of a Lipopolysaccharide and Resiquimod Combination as an Adjuvant with Inactivated Newcastle Disease Virus Vaccine in Chickens. Vaccines. 2022; 10(6):894. https://doi.org/10.3390/vaccines10060894
Chicago/Turabian StyleSharma, Bal Krishan, Saravanan Ramakrishnan, Abinaya Kaliappan, Mithilesh Singh, Ajay Kumar, Satyabrata Dandapat, Sohini Dey, and Madhan Mohan Chellappa. 2022. "Evaluation of a Lipopolysaccharide and Resiquimod Combination as an Adjuvant with Inactivated Newcastle Disease Virus Vaccine in Chickens" Vaccines 10, no. 6: 894. https://doi.org/10.3390/vaccines10060894
APA StyleSharma, B. K., Ramakrishnan, S., Kaliappan, A., Singh, M., Kumar, A., Dandapat, S., Dey, S., & Chellappa, M. M. (2022). Evaluation of a Lipopolysaccharide and Resiquimod Combination as an Adjuvant with Inactivated Newcastle Disease Virus Vaccine in Chickens. Vaccines, 10(6), 894. https://doi.org/10.3390/vaccines10060894