
Fiction and Facts about BCG Imparting Trained Immunity against COVID-19


 , , ,
, , ,
Abstract
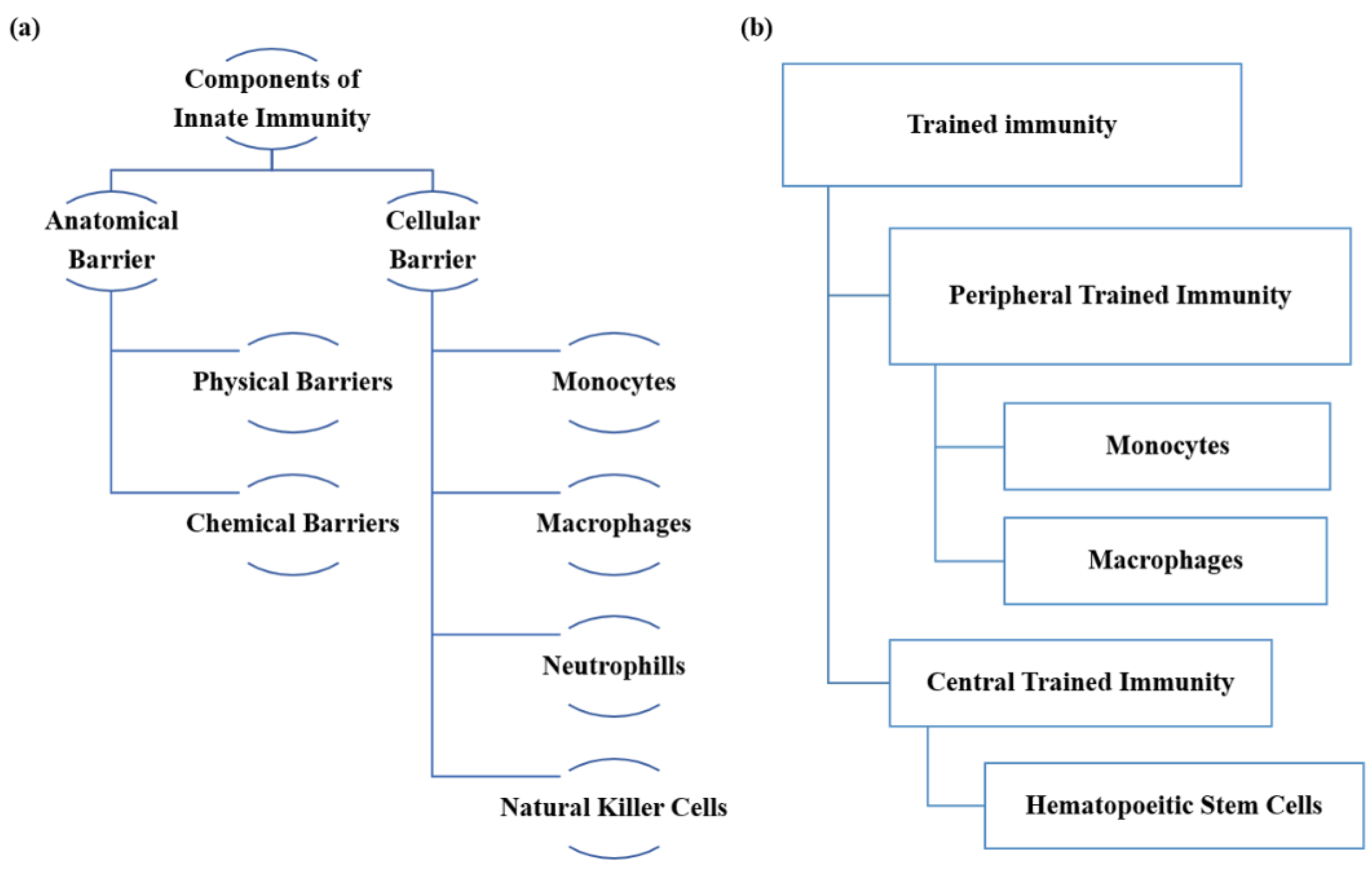
:1. Introduction
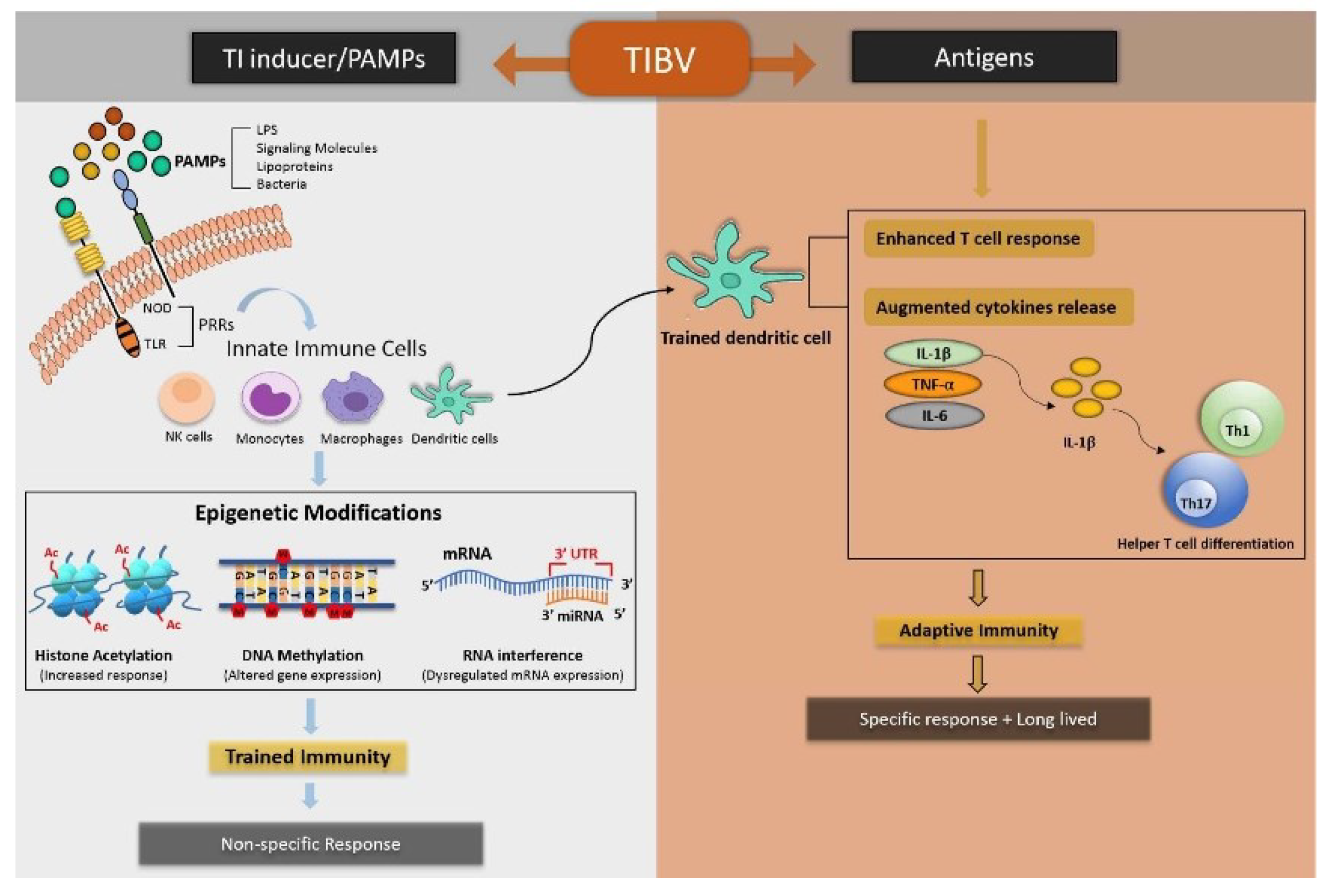
2. Induction of the activation of Trained Immunity
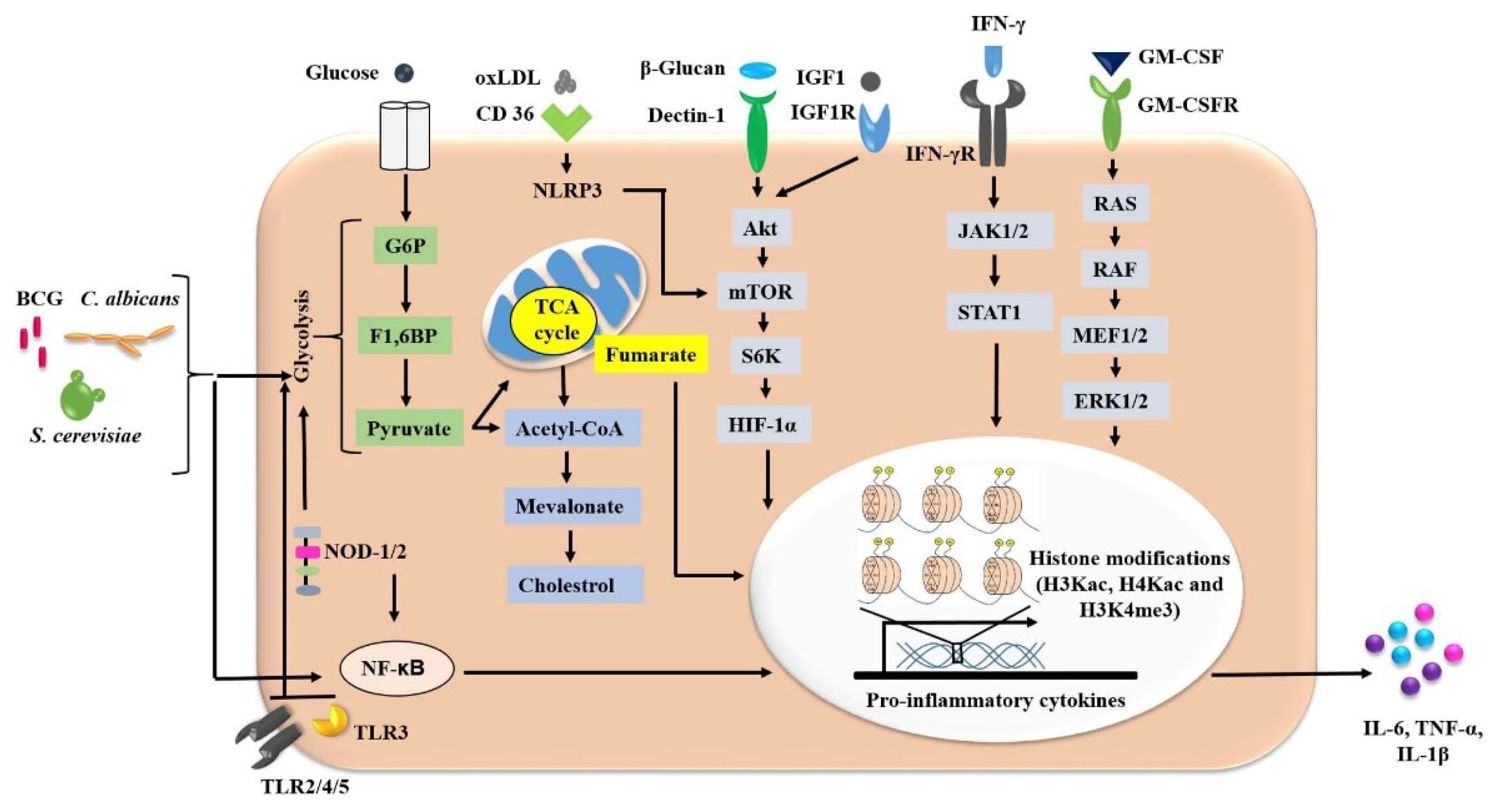
2.1. Trained-Immunity-Based Vaccines and Associated Mechanism
2.2. Clinical Applications of TIbVs
2.3. Role of Trained Immunity in Viral Diseases
2.4. Immune Response to SARS-CoV-2
2.5. BCG and Trained Immunity against COVID-19
3. Summary
4. Expert Opinion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Fortier, A.H.; Mock, B.A.; Meltzer, M.S.; Nacy, C.A. Mycobacterium bovis BCG-induced protection against cutaneous and systemic Leishmania major infections of mice. Infect. Immun. 1987, 55, 1707–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.; Ifrim, D.C.; Saeed, S.; Jacobs, C.; van Loenhout, J.; de Jong, D.; Stunnenberg, H.G.; et al. Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc. Natl. Acad. Sci. USA 2012, 109, 17537–17542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Subramaniam, R.; Chen, H.; Smith, A.; Keshava, S.; Shams, H. Boosting efferocytosis in alveolar space using BCG vaccine to protect host against influenza pneumonia. PLoS ONE 2017, 12, e0180143. [Google Scholar] [CrossRef] [PubMed]
- Floc’h, F.; Werner, G.H. Increased resistance to virus infections of mice inoculated with BCG (Bacillus calmette-guerin). Ann. D’immunol. 1976, 127, 173–186. [Google Scholar]
- Niobey, F.M.; Duchiade, M.P.; Vasconcelos, A.G.; de Carvalho, M.L.; Leal Mdo, C.; Valente, J.G. Risk factors for death caused by pneumonia in children younger than 1 year old in a metropolitan region of southeastern Brazil. A case- control study. Rev. Saude Publica 1992, 26, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Rieckmann, A.; Villumsen, M.; Sorup, S.; Haugaard, L.K.; Ravn, H.; Roth, A.; Baker, J.L.; Benn, C.S.; Aaby, P. Vaccinations against smallpox and tuberculosis are associated with better long-term survival: A Danish case-cohort study 1971–2010. Int. J. Epidemiol. 2017, 46, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Rieckmann, A.; Meyle, K.D.; Rod, N.H.; Baker, J.L.; Benn, C.S.; Aaby, P.; Sorup, S. Smallpox and BCG vaccination in childhood and cutaneous malignant melanoma in Danish adults followed from 18 to 49 years. Vaccine 2019, 37, 6730–6736. [Google Scholar] [CrossRef]
- Biering-Sørensen, S.; Aaby, P.; Lund, N.; Monteiro, I.; Jensen, K.J.; Eriksen, H.B.; Schaltz-Buchholzer, F.; Jørgensen, A.S.P.; Rodrigues, A.; Fisker, A.B.; et al. Early BCG-Denmark and Neonatal Mortality Among Infants Weighing <2500 g: A Randomized Controlled Trial. Clin. Infect. Dis. 2017, 65, 1183–1190. [Google Scholar] [CrossRef]
- Hollm-Delgado, M.G.; Stuart, E.A.; Black, R.E. Acute lower respiratory infection among Bacille Calmette-Guerin (BCG)-vaccinated children. Pediatrics 2014, 133, e73–e81. [Google Scholar] [CrossRef] [Green Version]
- Ohrui, T.; Nakayama, K.; Fukushima, T.; Chiba, H.; Sasaki, H. Prevention of elderly pneumonia by pneumococcal, influenza and BCG vaccinations. Nihon Ronen Igakkai Zasshi. Jpn. J. Geriatr. 2005, 42, 34–36. [Google Scholar] [CrossRef] [Green Version]
- Wardhana; Datau, E.A.; Sultana, A.; Mandang, V.V.; Jim, E. The efficacy of Bacillus Calmette-Guerin vaccinations for the prevention of acute upper respiratory tract infection in the elderly. Acta Med. Indones. 2011, 43, 185–190. [Google Scholar] [PubMed]
- Stensballe, L.G.; Nante, E.; Jensen, I.P.; Kofoed, P.E.; Poulsen, A.; Jensen, H.; Newport, M.; Marchant, A.; Aaby, P. Acute lower respiratory tract infections and respiratory syncytial virus in infants in Guinea-Bissau: A beneficial effect of BCG vaccination for girls community based case-control study. Vaccine 2005, 23, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.I.; Dorta, M.L.; Pereira, A.J.; Bastos, R.P.; Oliveira, M.A.; Pinto, S.A.; Galdino, H., Jr.; Mayrink, W.; Barcelos, W.; Toledo, V.P.; et al. Increase of NK cells and proinflammatory monocytes are associated with the clinical improvement of diffuse cutaneous leishmaniasis after immunochemotherapy with BCG/Leishmania antigens. Am. J. Trop. Med. Hyg. 2009, 81, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Leentjens, J.; Kox, M.; Stokman, R.; Gerretsen, J.; Diavatopoulos, D.A.; van Crevel, R.; Rimmelzwaan, G.F.; Pickkers, P.; Netea, M.G. BCG Vaccination Enhances the Immunogenicity of Subsequent Influenza Vaccination in Healthy Volunteers: A Randomized, Placebo-Controlled Pilot Study. J. Infect. Dis. 2015, 212, 1930–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavan Kumar, N.; Padmapriyadarsini, C.; Rajamanickam, A.; Marinaik, S.B.; Nancy, A.; Padmanaban, S.; Akbar, N.; Murhekar, M.; Babu, S. Effect of BCG vaccination on proinflammatory responses in elderly individuals. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; van Crevel, R.; Netea, M.G. Trained immunity: Consequences for the heterologous effects of BCG vaccination. Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 29–35. [Google Scholar] [CrossRef]
- de Castro, M.J.; Pardo-Seco, J.; Martinón-Torres, F. Nonspecific (Heterologous) Protection of Neonatal BCG Vaccination Against Hospitalization Due to Respiratory Infection and Sepsis. Clin. Infect. Dis. 2015, 60, 1611–1619. [Google Scholar] [CrossRef] [Green Version]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Benn, C.S.; Joosten, L.A.; Jacobs, C.; van Loenhout, J.; Xavier, R.J.; Aaby, P.; van der Meer, J.W.; et al. Long-lasting effects of BCG vaccination on both heterologous Th1/Th17 responses and innate trained immunity. J. Innate Immun. 2014, 6, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; van der Meer, J.W. Trained Immunity: An Ancient Way of Remembering. Cell Host Microbe 2017, 21, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Joosten, L.A.; Latz, E.; Mills, K.H.; Natoli, G.; Stunnenberg, H.G.; O’Neill, L.A.; Xavier, R.J. Trained immunity: A program of innate immune memory in health and disease. Science 2016, 352, 21. [Google Scholar] [CrossRef] [Green Version]
- Usher, N.T.; Chang, S.; Howard, R.S.; Martinez, A.; Harrison, L.H.; Santosham, M.; Aronson, N.E. Association of BCG Vaccination in Childhood With Subsequent Cancer Diagnoses: A 60-Year Follow-up of a Clinical Trial. JAMA Netw. Open 2019, 2. [Google Scholar] [CrossRef] [PubMed]
- Starr, S.E.; Visintine, A.M.; Tomeh, M.O.; Nahmias, A.J. Effects of immunostimulants on resistance of newborn mice to herpes simplex type 2 infection. Proc. Soc. Exp. Biol. Med. 1976, 152, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.C.; Ganguly, R.; Waldman, R.H. Nonspecific protection of mice against influenza virus infection by local or systemic immunization with Bacille Calmette-Guérin. J. Infect. Dis. 1977, 136, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Zens, K.D.; Chen, J.K.; Farber, D.L. Vaccine-generated lung tissue-resident memory T cells provide heterosubtypic protection to influenza infection. JCI Insight 2016, 1. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Quintin, J.; van der Meer, J.W. Trained immunity: A memory for innate host defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Mulder, W.J.M.; Ochando, J.; Joosten, L.A.B.; Fayad, Z.A.; Netea, M.G. Therapeutic targeting of trained immunity. Nat. Rev. Drug Discov. 2019, 18, 553–566. [Google Scholar] [CrossRef]
- Redelman-Sidi, G. Could BCG be used to protect against COVID-19? Nat. Rev. Urol. 2020, 17, 316–317. [Google Scholar] [CrossRef]
- Arts, R.J.; Blok, B.A.; Aaby, P.; Joosten, L.A.; de Jong, D.; van der Meer, J.W.; Benn, C.S.; van Crevel, R.; Netea, M.G. Long-term in vitro and in vivo effects of γ-irradiated BCG on innate and adaptive immunity. J. Leukoc. Biol. 2015, 98, 995–1001. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Netea, M.G. BCG-induced trained immunity: Can it offer protection against COVID-19? Nat. Rev. Immunol. 2020, 20, 335–337. [Google Scholar] [CrossRef]
- Sharma, A.R.; Batra, G.; Kumar, M.; Mishra, A.; Singla, R.; Singh, A.; Singh, R.S.; Medhi, B. BCG as a game-changer to prevent the infection and severity of COVID-19 pandemic? Allergol. Immunopathol. 2020, 48, 507–517. [Google Scholar] [CrossRef]
- WHO. Global Tuberculosis Report 2021; World Health Organization: Geneva, Switzerland, 2021; Licence: CC BY-NC-SA 3.0 IGO. Global TB report 2021; Available online: https://covid19.who.int/;WHO (accessed on 20 February 2022).
- Pahari, S.; Kaur, G.; Negi, S.; Aqdas, M.; Das, D.K.; Bashir, H.; Singh, S.; Nagare, M.; Khan, J.; Agrewala, J.N. Reinforcing the Functionality of Mononuclear Phagocyte System to Control Tuberculosis. Front. Immunol. 2018, 9, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bree, L.C.J.; Koeken, V.; Joosten, L.A.B.; Aaby, P.; Benn, C.S.; van Crevel, R.; Netea, M.G. Non-specific effects of vaccines: Current evidence and potential implications. Semin. Immunol. 2018, 39, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Walk, J.; de Bree, L.C.J.; Graumans, W.; Stoter, R.; van Gemert, G.J.; van de Vegte-Bolmer, M.; Teelen, K.; Hermsen, C.C.; Arts, R.J.W.; Behet, M.C.; et al. Outcomes of controlled human malaria infection after BCG vaccination. Nat. Commun. 2019, 10, 874. [Google Scholar] [CrossRef] [Green Version]
- Convit, J.; Ulrich, M.; Polegre, M.A.; Avila, A.; Rodríguez, N.; Mazzedo, M.I.; Blanco, B. Therapy of Venezuelan patients with severe mucocutaneous or early lesions of diffuse cutaneous leishmaniasis with a vaccine containing pasteurized Leishmania promastigotes and bacillus Calmette-Guerin: Preliminary report. Mem. Inst. Oswaldo Cruz 2004, 99, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Piñeiro, J.A.; Muntañola, P. Nonspecific immunotherapy with BCG vaccine in bladder tumors: A preliminary report. Eur. Urol. 1977, 3, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Barton, E.S.; White, D.W.; Cathelyn, J.S.; Brett-McClellan, K.A.; Engle, M.; Diamond, M.S.; Miller, V.L.; Virgin, H.W. Herpesvirus latency confers symbiotic protection from bacterial infection. Nature 2007, 447, 326–329. [Google Scholar] [CrossRef]
- Quintin, J.; Saeed, S.; Martens, J.H.A.; Giamarellos-Bourboulis, E.J.; Ifrim, D.C.; Logie, C.; Jacobs, L.; Jansen, T.; Kullberg, B.J.; Wijmenga, C.; et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe 2012, 12, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Saeed, S.; Quintin, J.; Kerstens, H.H.; Rao, N.A.; Aghajanirefah, A.; Matarese, F.; Cheng, S.C.; Ratter, J.; Berentsen, K.; van der Ent, M.A.; et al. Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science 2014, 345, 1251086. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Andrés, J.; Joosten, L.A.; Netea, M.G. Induction of innate immune memory: The role of cellular metabolism. Curr. Opin. Immunol. 2019, 56, 10–16. [Google Scholar] [CrossRef]
- Cheng, S.C.; Quintin, J.; Cramer, R.A.; Shepardson, K.M.; Saeed, S.; Kumar, V.; Giamarellos-Bourboulis, E.J.; Martens, J.H.; Rao, N.A.; Aghajanirefah, A.; et al. mTOR- and HIF-1α-mediated aerobic glycolysis as metabolic basis for trained immunity. Science 2014, 345, 1250684. [Google Scholar] [CrossRef] [Green Version]
- Guy, B. The perfect mix: Recent progress in adjuvant research. Nat. Rev. Microbiol. 2007, 5, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, I.; Tan, Y.; Di Gioia, M.; Broggi, A.; Ruan, J.; Shi, J.; Donado, C.A.; Shao, F.; Wu, H.; Springstead, J.R.; et al. An endogenous caspase-11 ligand elicits interleukin-1 release from living dendritic cells. Science 2016, 352, 1232–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aaby, P.; Gustafson, P.; Roth, A.; Rodrigues, A.; Fernandes, M.; Sodemann, M.; Holmgren, B.; Benn, C.S.; Garly, M.L.; Lisse, I.M.; et al. Vaccinia scars associated with better survival for adults. An observational study from Guinea-Bissau. Vaccine 2006, 24, 5718–5725. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.L.; Dave, S.; Schim van der Loeff, M.; da Costa, C.; Vincent, T.; Leligdowicz, A.; Benn, C.S.; Roth, A.; Ravn, H.; Lisse, I.M.; et al. Vaccinia scars associated with improved survival among adults in rural Guinea-Bissau. PLoS ONE 2006, 1, 101. [Google Scholar] [CrossRef] [PubMed]
- Mayr, A. Taking advantage of the positive side-effects of smallpox vaccination. J. Vet. Med. B Infect. Dis. Vet. Public Health 2004, 51, 199–201. [Google Scholar] [CrossRef]
- Kaufmann, E.; Sanz, J.; Dunn, J.L.; Khan, N.; Mendonça, L.E.; Pacis, A.; Tzelepis, F.; Pernet, E.; Dumaine, A.; Grenier, J.C.; et al. BCG Educates Hematopoietic Stem Cells to Generate Protective Innate Immunity against Tuberculosis. Cell 2018, 172, 176–190. [Google Scholar] [CrossRef] [Green Version]
- Gillard, G.O.; Bivas-Benita, M.; Hovav, A.H.; Grandpre, L.E.; Panas, M.W.; Seaman, M.S.; Haynes, B.F.; Letvin, N.L. Thy1+ NK [corrected] cells from vaccinia virus-primed mice confer protection against vaccinia virus challenge in the absence of adaptive lymphocytes. PLoS Pathog. 2011, 7, 4. [Google Scholar] [CrossRef]
- Kawai, K.; Miyazaki, J.; Joraku, A.; Nishiyama, H.; Akaza, H. Bacillus Calmette-Guerin (BCG) immunotherapy for bladder cancer: Current understanding and perspectives on engineered BCG vaccine. Cancer Sci. 2013, 104, 22–27. [Google Scholar] [CrossRef]
- Buffen, K.; Oosting, M.; Quintin, J.; Ng, A.; Kleinnijenhuis, J.; Kumar, V.; van de Vosse, E.; Wijmenga, C.; van Crevel, R.; Oosterwijk, E.; et al. Autophagy controls BCG-induced trained immunity and the response to intravesical BCG therapy for bladder cancer. PLoS Pathog. 2014, 10, e1004485. [Google Scholar] [CrossRef]
- Piedra, P.A.; Gaglani, M.J.; Kozinetz, C.A.; Herschler, G.B.; Fewlass, C.; Harvey, D.; Zimmerman, N.; Glezen, W.P. Trivalent live attenuated intranasal influenza vaccine administered during the 2003–2004 influenza type A (H3N2) outbreak provided immediate, direct, and indirect protection in children. Pediatrics 2007, 120, 2006–2836. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Lee, J.Y.; Jang, Y.H.; Seo, S.U.; Chang, J.; Seong, B.L. Non-specific Effect of Vaccines: Immediate Protection against Respiratory Syncytial Virus Infection by a Live Attenuated Influenza Vaccine. Front. Microbiol. 2018, 9, 83. [Google Scholar] [CrossRef] [PubMed]
- Arts, R.J.W.; Moorlag, S.; Novakovic, B.; Li, Y.; Wang, S.Y.; Oosting, M.; Kumar, V.; Xavier, R.J.; Wijmenga, C.; Joosten, L.A.B.; et al. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell Host Microbe 2018, 23, 89–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arts, R.J.W.; Carvalho, A.; La Rocca, C.; Palma, C.; Rodrigues, F.; Silvestre, R.; Kleinnijenhuis, J.; Lachmandas, E.; Gonçalves, L.G.; Belinha, A.; et al. Immunometabolic Pathways in BCG-Induced Trained Immunity. Cell Rep. 2016, 17, 2562–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, D.E.; Cleary, D.W.; Clarke, S.C. Secondary Bacterial Infections Associated with Influenza Pandemics. Front. Microbiol. 2017, 8, 1041–1058. [Google Scholar] [CrossRef] [Green Version]
- Blok, B.A.; Arts, R.J.; van Crevel, R.; Benn, C.S.; Netea, M.G. Trained innate immunity as underlying mechanism for the long-term, nonspecific effects of vaccines. J. Leukoc. Biol. 2015, 98, 347–356. [Google Scholar] [CrossRef]
- Lorenzo-Gómez, M.F.; Padilla-Fernández, B.; García-Criado, F.J.; Mirón-Canelo, J.A.; Gil-Vicente, A.; Nieto-Huertos, A.; Silva-Abuin, J.M. Evaluation of a therapeutic vaccine for the prevention of recurrent urinary tract infections versus prophylactic treatment with antibiotics. Int. Urogynecol. J. 2013, 24, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Alecsandru, D.; Valor, L.; Sánchez-Ramón, S.; Gil, J.; Carbone, J.; Navarro, J.; Rodríguez, J.; Rodríguez-Sainz, C.; Fernández-Cruz, E. Sublingual therapeutic immunization with a polyvalent bacterial preparation in patients with recurrent respiratory infections: Immunomodulatory effect on antigen-specific memory CD4+ T cells and impact on clinical outcome. Clin. Exp. Immunol. 2011, 164, 1365–2249. [Google Scholar] [CrossRef]
- Krone, B.; Kölmel, K.F.; Henz, B.M.; Grange, J.M. Protection against melanoma by vaccination with Bacille Calmette-Guerin (BCG) and/or vaccinia: An epidemiology-based hypothesis on the nature of a melanoma risk factor and its immunological control. Eur. J. Cancer 2005, 41, 104–117. [Google Scholar] [CrossRef]
- Wang, D.; Sun, S.; Heidari, M. Marek’s disease vaccine activates chicken macrophages. J. Vet. Sci. 2018, 19, 375–383. [Google Scholar] [CrossRef]
- Bardenheier, B.H.; McNeil, M.M.; Wodi, A.P.; McNicholl, J.M.; DeStefano, F. Risk of Nontargeted Infectious Disease Hospitalizations Among US Children Following Inactivated and Live Vaccines, 2005-2014. Clin. Infect. Dis. 2017, 65, 729–737. [Google Scholar] [CrossRef]
- La Torre, G.; Saulle, R.; Unim, B.; Meggiolaro, A.; Barbato, A.; Mannocci, A.; Spadea, A. The effectiveness of measles-mumps-rubella (MMR) vaccination in the prevention of pediatric hospitalizations for targeted and untargeted infections: A retrospective cohort study. Hum. Vaccines Immunother. 2017, 13, 1879–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorup, S.; Benn, C.S.; Poulsen, A.; Krause, T.G.; Aaby, P.; Ravn, H. Live vaccine against measles, mumps, and rubella and the risk of hospital admissions for nontargeted infections. JAMA 2014, 311, 826–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tielemans, S.; de Melker, H.E.; Hahne, S.J.M.; Boef, A.G.C.; van der Klis, F.R.M.; Sanders, E.A.M.; van der Sande, M.A.B.; Knol, M.J. Non-specific effects of measles, mumps, and rubella (MMR) vaccination in high income setting: Population based cohort study in the Netherlands. Bmj 2017, 358, j3862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, E.A.; Boulos, R.; Halsey, N.A.; Boulos, L.M.; Boulos, C. Childhood survival in Haiti: Protective effect of measles vaccination. Pediatrics 1990, 85, 188–194. [Google Scholar] [CrossRef]
- Seppala, E.; Viskari, H.; Hoppu, S.; Honkanen, H.; Huhtala, H.; Simell, O.; Ilonen, J.; Knip, M.; Hyoty, H. Viral interference induced by live attenuated virus vaccine (OPV) can prevent otitis media. Vaccine 2011, 29, 8615–8618. [Google Scholar] [CrossRef]
- Upfill-Brown, A.; Taniuchi, M.; Platts-Mills, J.A.; Kirkpatrick, B.; Burgess, S.L.; Oberste, M.S.; Weldon, W.; Houpt, E.; Haque, R.; Zaman, K.; et al. Nonspecific Effects of Oral Polio Vaccine on Diarrheal Burden and Etiology Among Bangladeshi Infants. Clin. Infect. Dis. 2017, 65, 414–419. [Google Scholar] [CrossRef] [Green Version]
- Kline, K.A.; Bowdish, D.M. Infection in an aging population. Curr. Opin. Microbiol. 2016, 29, 63–67. [Google Scholar] [CrossRef]
- Novakovic, B.; Habibi, E.; Wang, S.Y.; Arts, R.J.W.; Davar, R.; Megchelenbrink, W.; Kim, B.; Kuznetsova, T.; Kox, M.; Zwaag, J.; et al. β-Glucan Reverses the Epigenetic State of LPS-Induced Immunological Tolerance. Cell 2016, 167, 1354–1368. [Google Scholar] [CrossRef] [Green Version]
- Błach-Olszewska, Z. Innate immunity: Cells, receptors, and signaling pathways. Arch. Immunol. Ther. Exp. 2005, 53, 245–253. [Google Scholar]
- Kirwan, S.E.; Burshtyn, D.N. Regulation of natural killer cell activity. Curr. Opin. Immunol. 2007, 19, 46–54. [Google Scholar] [CrossRef]
- Nieto, A.; Mazon, A.; Nieto, M.; Calderon, R.; Calaforra, S.; Selva, B.; Uixera, S.; Palao, M.J.; Brandi, P.; Conejero, L.; et al. Bacterial Mucosal Immunotherapy with MV130 Prevents Recurrent Wheezing in Children: A Randomized, Double-blind, Placebo-controlled Trial. Am. J. Respir. Crit. Care Med. 2021, 204, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Moorlag, S.; Arts, R.J.W.; van Crevel, R.; Netea, M.G. Non-specific effects of BCG vaccine on viral infections. Clin. Microbiol. Infect. 2019, 25, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.; Jacobs, C.; Xavier, R.J.; van der Meer, J.W.; van Crevel, R.; Netea, M.G. BCG-induced trained immunity in NK cells: Role for non-specific protection to infection. Clin. Immunol. 2014, 155, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelman, A.E.; LaRosa, D.F.; Zhang, J.; Walsh, P.T.; Choi, Y.; Sunyer, J.O.; Turka, L.A. The adaptor molecule MyD88 activates PI-3 kinase signaling in CD4+ T cells and enables CpG oligodeoxynucleotide-mediated costimulation. Immunity 2006, 25, 783–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.C.; Beilke, J.N.; Lanier, L.L. Adaptive immune features of natural killer cells. Nature 2009, 457, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Machiels, B.; Dourcy, M.; Xiao, X.; Javaux, J.; Mesnil, C.; Sabatel, C.; Desmecht, D.; Lallemand, F.; Martinive, P.; Hammad, H.; et al. A gammaherpesvirus provides protection against allergic asthma by inducing the replacement of resident alveolar macrophages with regulatory monocytes. Nat. Immunol. 2017, 18, 1310–1320. [Google Scholar] [CrossRef]
- Yao, Y.; Jeyanathan, M.; Haddadi, S.; Barra, N.G.; Vaseghi-Shanjani, M.; Damjanovic, D.; Lai, R.; Afkhami, S.; Chen, Y.; Dvorkin-Gheva, A.; et al. Induction of Autonomous Memory Alveolar Macrophages Requires T Cell Help and Is Critical to Trained Immunity. Cell 2018, 175, 1634–1650.e17. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Liu, D.X. The ER stress sensor IRE1 and MAP kinase ERK modulate autophagy induction in cells infected with coronavirus infectious bronchitis virus. Virology 2019, 533, 34–44. [Google Scholar] [CrossRef]
- Imai, Y.; Kuba, K.; Rao, S.; Huan, Y.; Guo, F.; Guan, B.; Yang, P.; Sarao, R.; Wada, T.; Leong-Poi, H.; et al. Angiotensin-converting enzyme 2 protects from severe acute lung failure. Nature 2005, 436, 112–116. [Google Scholar] [CrossRef]
- Chu, H.; Chan, J.F.; Wang, Y.; Yuen, T.T.; Chai, Y.; Hou, Y.; Shuai, H.; Yang, D.; Hu, B.; Huang, X.; et al. Comparative Replication and Immune Activation Profiles of SARS-CoV-2 and SARS-CoV in Human Lungs: An Ex Vivo Study With Implications for the Pathogenesis of COVID-19. Clin. Infect. Dis. 2020, 71, 1400–1409. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045. [Google Scholar] [CrossRef]
- Zhou, Y.; Fu, B.; Zheng, X.; Wang, D.; Zhao, C.; Qi, Y.; Sun, R.; Tian, Z.; Xu, X.; Wei, H. Pathogenic T-cells and inflammatory monocytes incite inflammatory storms in severe COVID-19 patients. Natl. Sci. Rev. 2020, 7, 998–1002. [Google Scholar] [CrossRef] [Green Version]
- Totura, A.L.; Whitmore, A.; Agnihothram, S.; Schäfer, A.; Katze, M.G.; Heise, M.T.; Baric, R.S. Toll-Like Receptor 3 Signaling via TRIF Contributes to a Protective Innate Immune Response to Severe Acute Respiratory Syndrome Coronavirus Infection. mBio 2015, 6, e00638-15. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.Y.; Huang, Y.; Ganesh, L.; Leung, K.; Kong, W.P.; Schwartz, O.; Subbarao, K.; Nabel, G.J. pH-dependent entry of severe acute respiratory syndrome coronavirus is mediated by the spike glycoprotein and enhanced by dendritic cell transfer through DC-SIGN. J. Virol. 2004, 78, 5642–5650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vankadari, N.; Wilce, J.A. Emerging WuHan (COVID-19) Coronavirus: Glycan Shield and Structure Prediction of Spike Glycoprotein and Its Interaction with Human CD26. Emerg. Microbes Infect. 2020, 9, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, C.; Schlossman, S.F. The structure and function of CD26 in the T-cell immune response. Immunol. Rev. 1998, 161, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Codo, A.C.; Davanzo, G.G.; Monteiro, L.B.; de Souza, G.F.; Muraro, S.P.; Virgilio-da-Silva, J.V.; Prodonoff, J.S.; Carregari, V.C.; de Biagi Junior, C.A.O.; Crunfli, F.; et al. Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis. Cell Metab. 2020, 32, 498–499. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Zhang, B.; Chassaing, B.; Shi, Z.; Uchiyama, R.; Zhang, Z.; Denning, T.L.; Crawford, S.E.; Pruijssers, A.J.; Iskarpatyoti, J.A.; Estes, M.K.; et al. Viral infection. Prevention and cure of rotavirus infection via TLR5/NLRC4-mediated production of IL-22 and IL-18. Science 2014, 346, 861–865. [Google Scholar] [CrossRef] [Green Version]
- Golonka, R.M.; Saha, P.; Yeoh, B.S.; Chattopadhyay, S.; Gewirtz, A.T.; Joe, B.; Vijay-Kumar, M. Harnessing innate immunity to eliminate SARS-CoV-2 and ameliorate COVID-19 disease. Physiol. Genomics. 2020, 52, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Basak, P.; Sachdeva, N.; Dayal, D. Can BCG vaccine protect against COVID-19 via trained immunity and tolerogenesis? BioEssays News Rev. Mol. Cell. Dev. Biol. 2020, 43, e2000200. [Google Scholar] [CrossRef]
- Mohamed Hussein, A.A.R.; Salem, M.R.; Salman, S.; Abdulrahim, A.F.; Al Massry, N.A.; Saad, M.; Dahman, N.B.H.; Negida, A. Correlation between COVID-19 case fatality rate and percentage of BCG vaccination: Is it true the vaccine is protective? Egypt. J. Bronchol. 2020, 14, 25. [Google Scholar] [CrossRef]
- Green, C.M.; Fanucchi, S.; Fok, E.T.; Moorlag, S.J.C.F.M.; Dominguez-Andres, J.; Negishi, Y.; Joosten, L.A.B.; Netea, M.G.; Mhlanga, M.M. COVID-19: A model correlating BCG vaccination to protection from mortality implicates trained immunity. medRxiv 2020. [Google Scholar] [CrossRef]
- Covián, C.; Retamal-Díaz, A.; Bueno, S.M.; Kalergis, A.M. Could BCG Vaccination Induce Protective Trained Immunity for SARS-CoV-2? Front. Immunol. 2020, 11, 970. [Google Scholar] [CrossRef] [PubMed]
- Rakshit, S.; Adiga, V.; Ahmed, A.; Parthiban, C.; Kumar, N.; Shivalingaiah, S.; Rao, S.; D’Souza, G.; Dwarkanath, P.; Dias, M.; et al. BCG revaccination qualitatively and quantitatively enhances SARS-CoV-2 spike-specific neutralizing antibody and T cell responses induced by the COVISHIELDTM vaccine in SARS-CoV-2 seronegative young Indian adults. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Kumar, N.P.; Padmapriyadarsini, C.; Rajamanickam, A.; Bhavani, P.K.; Nancy, A.; Jayadeepa, B.; Selvaraj, N.; Asokan, D.; Renji, R.M.; Venkataramani, V.; et al. BCG vaccination induces enhanced frequencies of memory T cells and altered plasma levels of common γc cytokines in elderly individuals. PLoS ONE 2021, 16, e0258743. [Google Scholar] [CrossRef] [PubMed]
- Covián, C.; Fernández-Fierro, A.; Retamal-Díaz, A.; Díaz, F.E.; Vasquez, A.E.; Lay, M.K.; Riedel, C.A.; González, P.A.; Bueno, S.M.; Kalergis, A.M. BCG-Induced Cross-Protection and Development of Trained Immunity: Implication for Vaccine Design. Front. Immunol. 2019, 10, 2806. [Google Scholar] [CrossRef] [PubMed]
- Soto, J.A.; Díaz, F.E.; Retamal-Díaz, A.; Gálvez, N.M.S.; Melo-González, F.; Piña-Iturbe, A.; Ramírez, M.A.; Bohmwald, K.; González, P.A.; Bueno, S.M.; et al. BCG-Based Vaccines Elicit Antigen-Specific Adaptive and Trained Immunity against SARS-CoV-2 and Andes orthohantavirus. Vaccines 2022, 10, 721. [Google Scholar] [CrossRef] [PubMed]
- Giamarellos-Bourboulis, E.J.; Tsilika, M.; Moorlag, S.; Antonakos, N.; Kotsaki, A.; Domínguez-Andrés, J.; Kyriazopoulou, E.; Gkavogianni, T.; Adami, M.E.; Damoraki, G.; et al. Activate: Randomized Clinical Trial of BCG Vaccination against Infection in the Elderly. Cell 2020, 183, 315–323.e9. [Google Scholar] [CrossRef]
- Stensballe, L.G.; Ravn, H.; Birk, N.M.; Kjaergaard, J.; Nissen, T.N.; Pihl, G.T.; Thostesen, L.M.; Greisen, G.; Jeppesen, D.L.; Kofoed, P.E.; et al. BCG Vaccination at Birth and Rate of Hospitalization for Infection Until 15 Months of Age in Danish Children: A Randomized Clinical Multicenter Trial. J. Pediatric Infect. Dis. Soc. 2019, 8, 213–220. [Google Scholar] [CrossRef]
- Kjaergaard, J.; Birk, N.M.; Nissen, T.N.; Thostesen, L.M.; Pihl, G.T.; Benn, C.S.; Jeppesen, D.L.; Pryds, O.; Kofoed, P.E.; Aaby, P.; et al. Nonspecific effect of BCG vaccination at birth on early childhood infections: A randomized, clinical multicenter trial. Pediatric Res. 2016, 80, 681–685. [Google Scholar] [CrossRef] [Green Version]
- Hamiel, U.; Kozer, E.; Youngster, I. SARS-CoV-2 Rates in BCG-Vaccinated and Unvaccinated Young Adults. JAMA 2020, 323, 2340–2341. [Google Scholar] [CrossRef]
- Hegarty, P.K.; Sfakianos, J.P.; Giannarini, G.; DiNardo, A.R.; Kamat, A.M. COVID-19 and Bacillus Calmette-Guérin: What is the Link? Eur. Urol. Oncol. 2020, 3, 259–261. [Google Scholar] [CrossRef]
- Lindestam Arlehamn, C.S.; Sette, A.; Peters, B. Lack of evidence for BCG vaccine protection from severe COVID-19. Proc. Natl. Acad. Sci. USA 2020, 117, 25203–25204. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Schlitzer, A.; Placek, K.; Joosten, L.A.B.; Schultze, J.L. Innate and Adaptive Immune Memory: An Evolutionary Continuum in the Host’s Response to Pathogens. Cell Host Microbe 2019, 25, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kantor, I.N. BCG versus COVID-19? Medicina 2020, 80, 292–294. [Google Scholar] [PubMed]
- Cernuschi, T.; Malvolti, S.; Nickels, E.; Friede, M. Bacillus Calmette-Guerin (BCG) vaccine: A global assessment of demand and supply balance. Vaccine 2018, 36, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Goonetilleke, N.P.; McShane, H.; Hannan, C.M.; Anderson, R.J.; Brookes, R.H.; Hill, A.V. Enhanced immunogenicity and protective efficacy against Mycobacterium tuberculosis of bacille Calmette-Guerin vaccine using mucosal administration and boosting with a recombinant modified vaccinia virus Ankara. J. Immunol. 2003, 171, 1602–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darrah, P.A.; Zeppa, J.J.; Maiello, P.; Hackney, J.A.; Wadsworth, M.H., 2nd; Hughes, T.K.; Pokkali, S.; Swanson, P.A., 2nd; Grant, N.L.; Rodgers, M.A.; et al. Prevention of tuberculosis in macaques after intravenous BCG immunization. Nature 2020, 577, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Verreck, F.A.W.; Tchilian, E.Z.; Vervenne, R.A.W.; Sombroek, C.C.; Kondova, I.; Eissen, O.A.; Sommandas, V.; van der Werff, N.M.; Verschoor, E.; Braskamp, G.; et al. Variable BCG efficacy in rhesus populations: Pulmonary BCG provides protection where standard intra-dermal vaccination fails. Tuberculosis 2017, 104, 46–57. [Google Scholar] [CrossRef]
- Yitbarek, K.; Abraham, G.; Girma, T.; Tilahun, T.; Woldie, M. The effect of Bacillus Calmette-Guérin (BCG) vaccination in preventing severe infectious respiratory diseases other than TB: Implications for the COVID-19 pandemic. Vaccine 2020, 38, 6374–6380. [Google Scholar] [CrossRef]
- Morawska, M. Reasons and consequences of COVID-19 vaccine failure in patients with chronic lymphocytic leukemia. Eur. J. Haematol. 2021, 108, 91–98. [Google Scholar] [CrossRef]
- Reischig, T.; Kacer, M.; Vlas, T.; Drenko, P.; Kielberger, L.; Machova, J.; Topolcan, O.; Kucera, R.; Kormunda, S. Insufficient response to mRNA SARS-CoV-2 vaccine and high incidence of severe COVID-19 in kidney transplant recipients during pandemic. Am. J. Transplant. Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2021, 22, 801–812. [Google Scholar] [CrossRef]
- Madhi, S.A.; Baillie, V.; Cutland, C.L.; Voysey, M.; Koen, A.L.; Fairlie, L.; Padayachee, S.D.; Dheda, K.; Barnabas, S.L.; Bhorat, Q.E.; et al. Efficacy of the ChAdOx1 nCoV-19 Covid-19 Vaccine against the B.1.351 Variant. N. Engl. J. Med. 2021, 384, 1885–1898. [Google Scholar] [CrossRef] [PubMed]
- Verma, I.; Grover, A. Antituberculous vaccine development: A perspective for the endemic world. Expert Rev. Vaccines 2009, 8, 1547–1553. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cross-Protection | Model | Remarks |
|---|---|---|
| L. major | Mice | 30–50% [1] |
| C. albicans | Mice | 100% survival in BCG-vaccinated mice vs. 30% in control mice [2] |
| Influenza A | Mice | 100% of mice immunized intranasally with BCG survived lethal IAV infection [3] |
| HSV1 | Mice | BCG-inoculated mice: 41%; survival in control: 18% [4] |
| Pneumonia | Children (case-controlled study) | 50% protection [5] |
| Sepsis | Case–control study | Reduced childhood mortality; better long-term survival [6] |
| Cutaneous malignant melanoma | Case–cohort study | No strong beneficial effect [7] |
| Pneumonia and sepsis | Randomized controlled trials | 43% reduction in infectious disease mortality; 38% reduction within neonatal period [8] |
| Respiratory tract infections | Data analysis based on Demographic and Health Surveys data | 17% to 37% risk in reduction [9] |
| Elderly pneumonia | Clinical trials | The risk of pneumonia was significantly decreased [10] |
| Acute URTIs respiratory tract infection | Humans | Protective effect on URTIs [11] |
| Acute lower respiratory tract infection (ALRI) | Infants | BCG vaccination may have a non-targeted protective effect against ALRI [12] |
| Leishmania amazonensis | Humans | A strong association between the increase of the frequency of innate immune system cells and the healing of lesions [13] |
| Influenza virus | Human study | Combined vaccination of BCG and influenza improved immunity against pandemic influenza A (H1N1) [14] |
| SARS-CoV-2 | Healthy elderly individuals | BCG vaccination down-regulates circulating inflammatory markers IL-10 and IL-33 and does not lead to increased inflammation in elderly individuals [15] |
| S. No | Clinical Trial Number | Location | Title | Date of Recruitment | Interventions |
|---|---|---|---|---|---|
| 1 | NCT04328441 | Netherlands | Reducing Health Care Workers’ Absenteeism in COVID-19 Pandemic Through BCG Vaccine (BCG-CORONA) | 31 March 2020 | BCG vaccine vs. placebo |
| 2 | NCT04659941 | Brazil | Use of BCG Vaccine as a Preventive Measure for COVID-19 in Health Care Workers (ProBCG) | 9 December 2020 | BCG Vaccine |
| 3 | NCT04347876 | Egypt | Outcome of COVID-19 Cases Based on Tuberculin Test: Can Previous BCG Alter the Prognosis? | 15 April 2020 | Diagnostic Test: Tuberculin test |
| 4 | NCT04348370 | USA | BCG Vaccine for Health Care Workers as Defense Against COVID-19 (BADAS) | 16 April 2020 | BCG vaccine vs. placebo |
| 5 | NCT04350931 | Egypt | Application of BCG Vaccine for Immune Prophylaxis Among Egyptian Healthcare Workers During the Pandemic of COVID-19 | 17 April 2020 | BCG vaccine vs. placebo |
| 6 | NCT04362124 | Columbia | Performance Evaluation of BCG Vaccination in Healthcare Personnel to Reduce the Severity of SARS-CoV-2 Infection | 24 April 2020 | BCG vaccine vs. placebo |
| 7 | NCT04369794 | Brazil | COVID-19: BCG As Therapeutic Vaccine, Transmission Limitation, and Immunoglobulin Enhancement (BATTLE) | 30 April 2020 | BCG vaccine vs. placebo |
| 8 | NCT04373291 | Denmark | Using BCG Vaccine to Protect Health Care Workers in the COVID-19 Pandemic | 4 May 2020 | BCG-Denmark vs. saline |
| 9 | NCT04379336 | South Africa | BCG Vaccination for Healthcare Workers in COVID-19 Pandemic | 7 May 2020 | BCG vaccine vs. placebo |
| 10 | NCT04384549 | France | Efficacy of BCG Vaccination in the Prevention of COVID19 Via the Strengthening of Innate Immunity in Health Care Workers (COVID-BCG) | 12 May 2020 | BCG vaccine vs. placebo |
| 11 | NCT04414267 | Netherlands | Bacillus Calmette-Guerin Vaccination to Prevent COVID-19 (ACTIVATEII) | 4 June 2020 | BCG vaccine vs. placebo |
| 12 | NCT04461379 | Mexico | Prevention, Efficacy and Safety of BCG Vaccine in COVID-19 Among Healthcare Workers | 8 July 2020 | BCG vaccine vs. placebo |
| 13 | NCT04475302 | India | BCG Vaccine in Reducing Morbidity and Mortality in Elderly Individuals in COVID-19 Hotspots | 17 July 2020 | BCG vaccine (Freeze-dried) |
| 14 | NCT04534803 | USA | BCG Against COVID-19 for Prevention and Amelioration of Severity Trial (BAC to the PAST) | 1 September 2020 | BCG vaccine vs. placebo |
| 15 | NCT04537663 | Netherlands | Prevention of Respiratory Tract Infection And COVID-19 Through BCG Vaccination in Vulnerable Older Adults (BCGPRIME) | 3 September 2020 | BCG vaccine vs. placebo |
| 16 | NCT04542330 | Denmark | Using BCG to Protect Senior Citizens During the COVID-19 Pandemic | 9 September 2020 | BCG-Denmark vs. saline |
| 17 | NCT04327206 | Australia | Efficacy of BCG Vaccination in the Prevention of COVID19 Via the Strengthening of Innate Immunity in Health Care Workers (BRACE) | 31 March 2020 | BCG vaccine vs. 0.9% NaCl |
| 18 | NCT04632537 | United States | BCG Vaccination to Prevent COVID-19 (NUEVA) | 17 November 2020 | Tice BCG vs. saline |
| 19 | NL8547 | Netherlands | Reducing Hospital Admission of Elderly in Sars-Cov-2 Pandemic Via the Induction of Trained Immunity By Bacillus Calmette-Guerin Vaccination, A Randomized Controlled Trial (BCG-CORONA Elderly) | May 2020 | BCG vs. placebo |
| 20 | CTRI/2020/05/025013 | India | Evaluation of BCG as potential therapy for COVID-19 | 6 May 2020 | BCG vaccine vs. saline |
| 21 | NCT04641858 | Denmark | BCG to Reduce Absenteeism Among Health Care Workers During the COVID-19 Pandemic (EDCTP) | 24 November 2020 | BCG vaccine Danish strain vs. saline |
| 22 | IRCT202004 11047019N1 | Iran | Investigating the Effect of BCG Vaccine on Preventing COVID-19 Infection in Healthcare Staff Exposed to SARS-CoV-2 | May 2020 | BCG vaccine vs. saline |
| 23 | EUCTR2020-001888- 90-DK | Denmark | To Reduce Absenteeism among Health Care Workers with Direct Patient Contact during the COVID-19 Pandemic | 30 April 2020 | BCG Danish strain vs. Placebo |
| 24 | EUCTR2020-002503- 19-GB | Spain; Australia; Netherlands; United Kingdom | BCG Vaccination to Reduce the Impact of COVID-19 on Health Care Workers | 8 July 2020 | BCG vs. Placebo |
| 25 | CTRI/2020/04/024833 | India | Effect of BCG-Denmark (Green Signal) on Prevention of COVID-19 Infection in Health Care Workers—A Double-Blind Randomized Controlled trial | 1 May 2020 | BCG vs. saline |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, G.; Singh, S.; Nanda, S.; Zafar, M.A.; Malik, J.A.; Arshi, M.U.; Lamba, T.; Agrewala, J.N. Fiction and Facts about BCG Imparting Trained Immunity against COVID-19. Vaccines 2022, 10, 1006. https://doi.org/10.3390/vaccines10071006
Kaur G, Singh S, Nanda S, Zafar MA, Malik JA, Arshi MU, Lamba T, Agrewala JN. Fiction and Facts about BCG Imparting Trained Immunity against COVID-19. Vaccines. 2022; 10(7):1006. https://doi.org/10.3390/vaccines10071006
Chicago/Turabian StyleKaur, Gurpreet, Sanpreet Singh, Sidhanta Nanda, Mohammad Adeel Zafar, Jonaid Ahmad Malik, Mohammad Umar Arshi, Taruna Lamba, and Javed Naim Agrewala. 2022. "Fiction and Facts about BCG Imparting Trained Immunity against COVID-19" Vaccines 10, no. 7: 1006. https://doi.org/10.3390/vaccines10071006
APA StyleKaur, G., Singh, S., Nanda, S., Zafar, M. A., Malik, J. A., Arshi, M. U., Lamba, T., & Agrewala, J. N. (2022). Fiction and Facts about BCG Imparting Trained Immunity against COVID-19. Vaccines, 10(7), 1006. https://doi.org/10.3390/vaccines10071006