
A Novel Therapeutic Tumor Vaccine Targeting MUC1 in Combination with PD-L1 Elicits Specific Anti-Tumor Immunity in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Cell Lines
2.2. Protein Production and Purification
2.3. Lentiviral-Transduced Tumor Cell Lines
2.4. Preparation of Bone Marrow-Derived Dendritic Cells (BMDCs)
2.5. Tumor Model and Vaccination
2.6. Flow Cytometry Analysis
2.7. Antibody ELISA
2.8. Hematoxylin-Eosin (HE) Staining
2.9. Statistical Analysis
3. Results
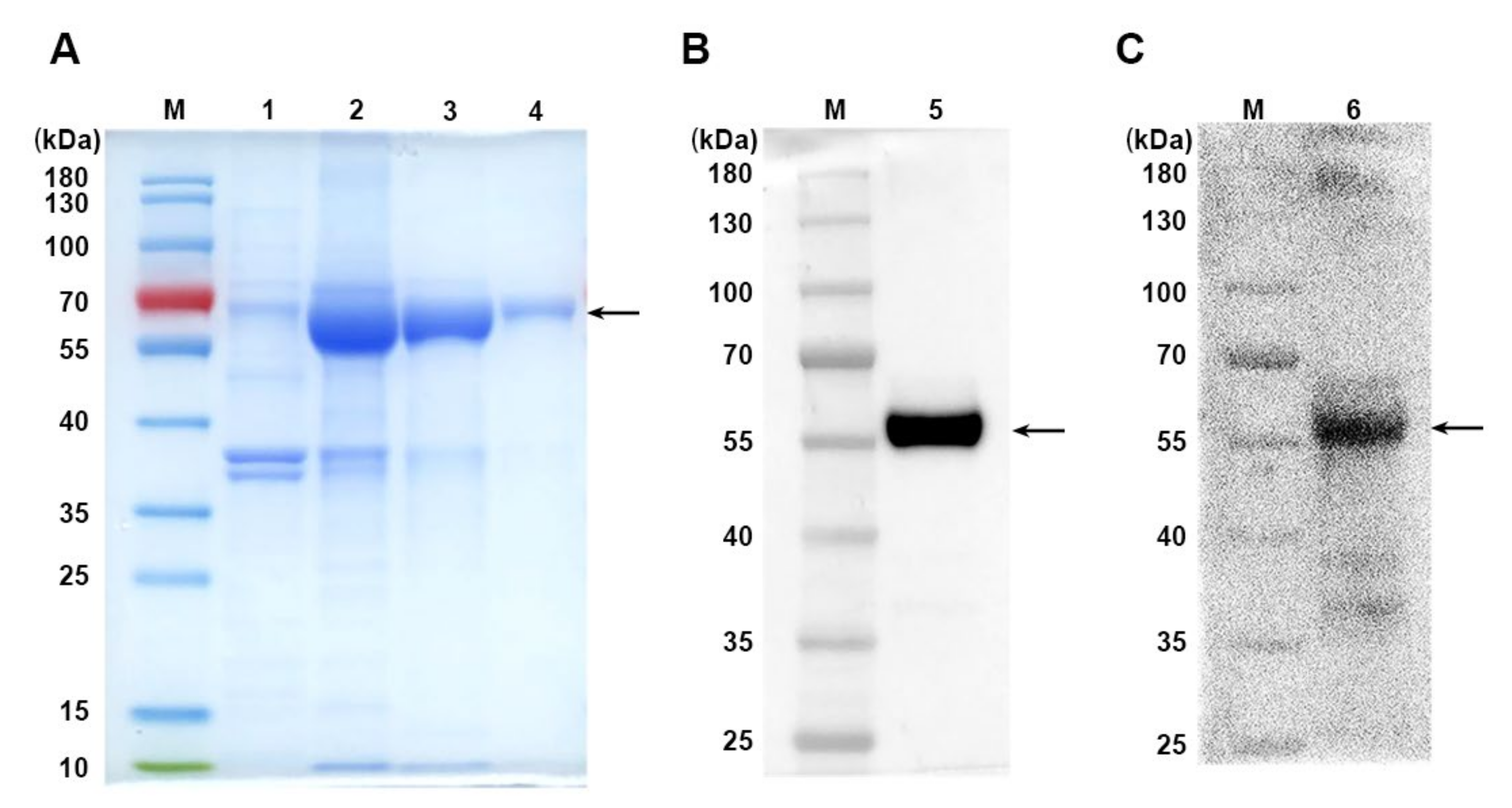
3.1. Production of Recombinant MUC1-PDL1-IgG1 Fc Immunogen (MUC1-Vax)
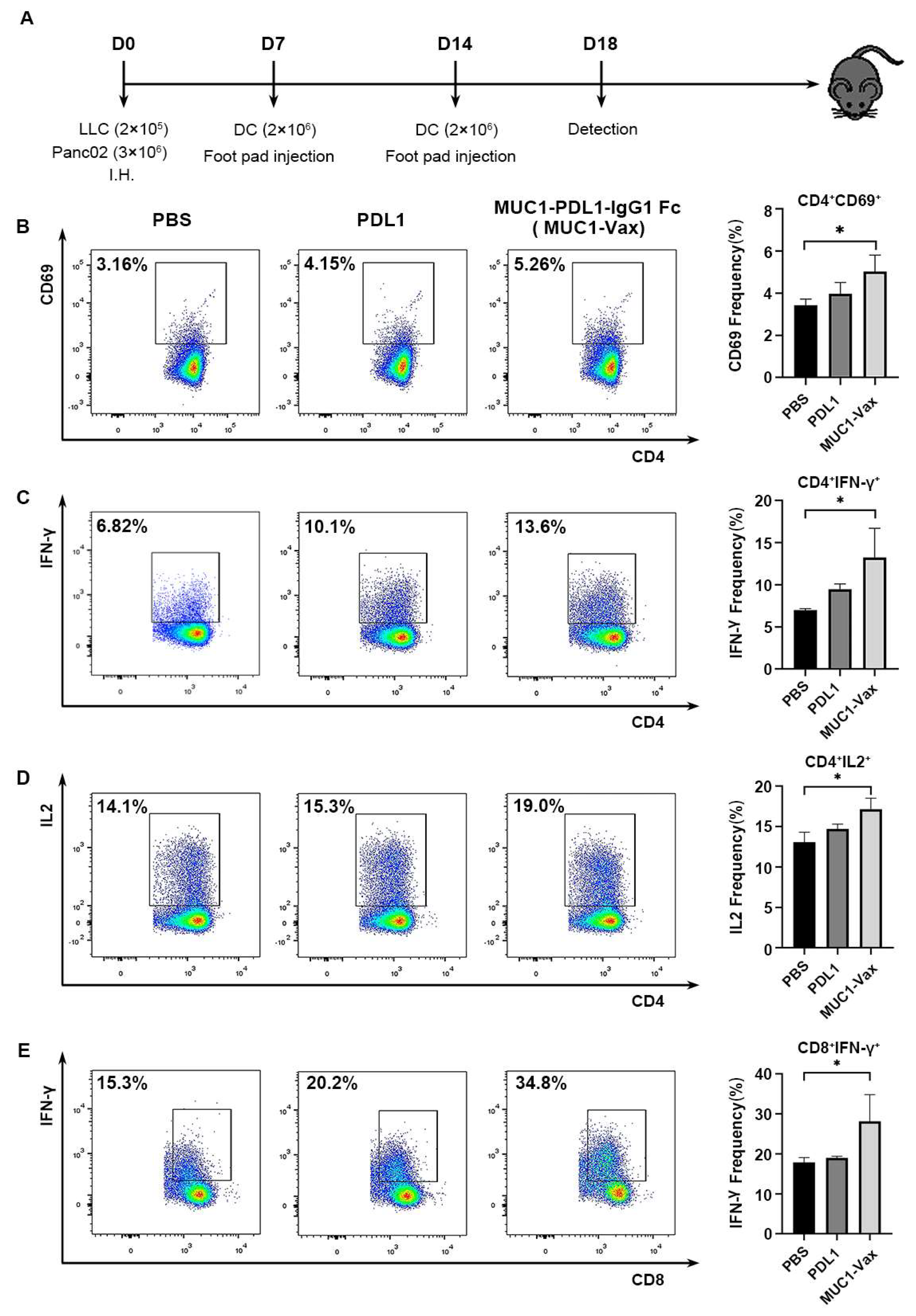
3.2. MUC1-Vax-Loaded DC Vaccine Induces Specific T Cell Activation and Cytokine Secretion
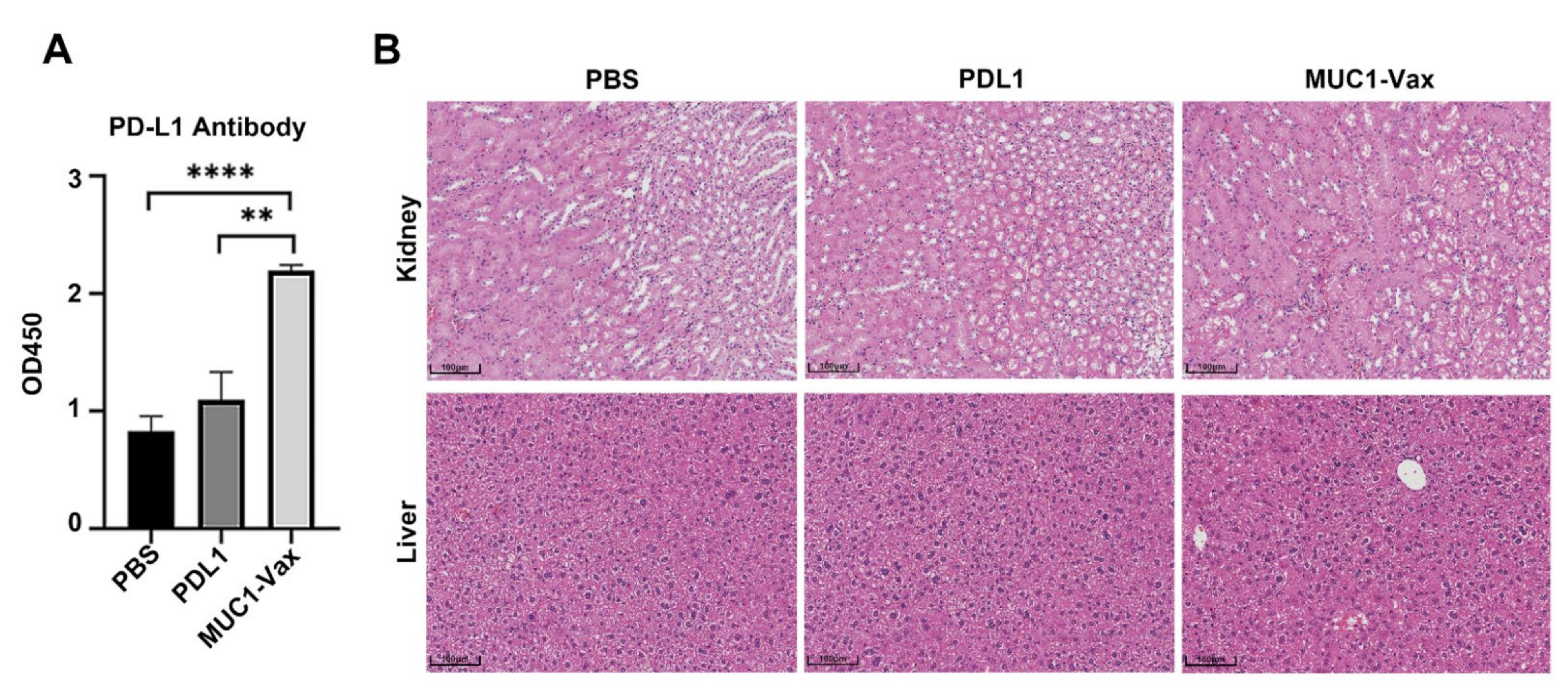
3.3. Anti-PD-L1 Antibody Response Induced by MUC1-Vax-Loaded DC Vaccine
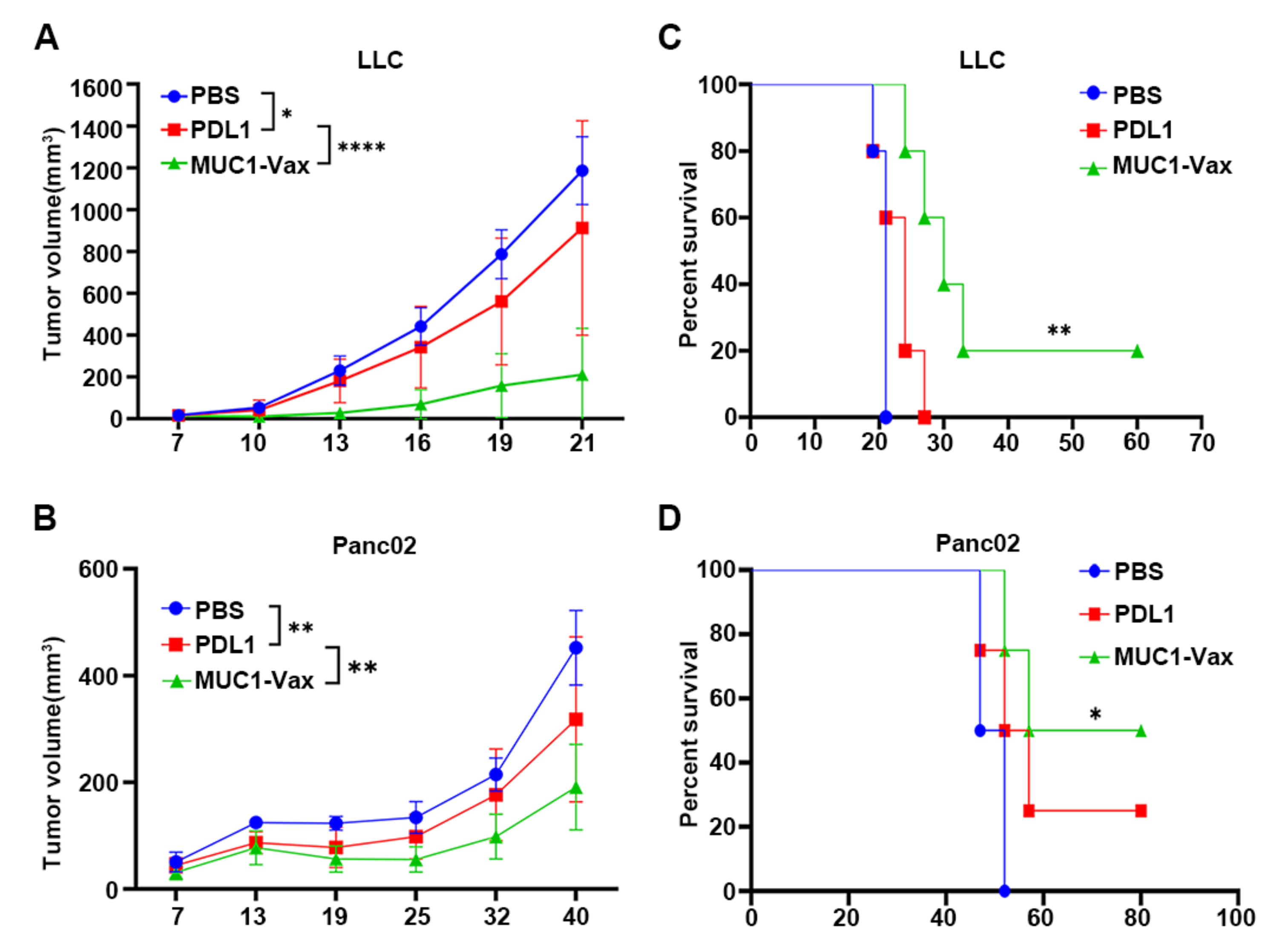
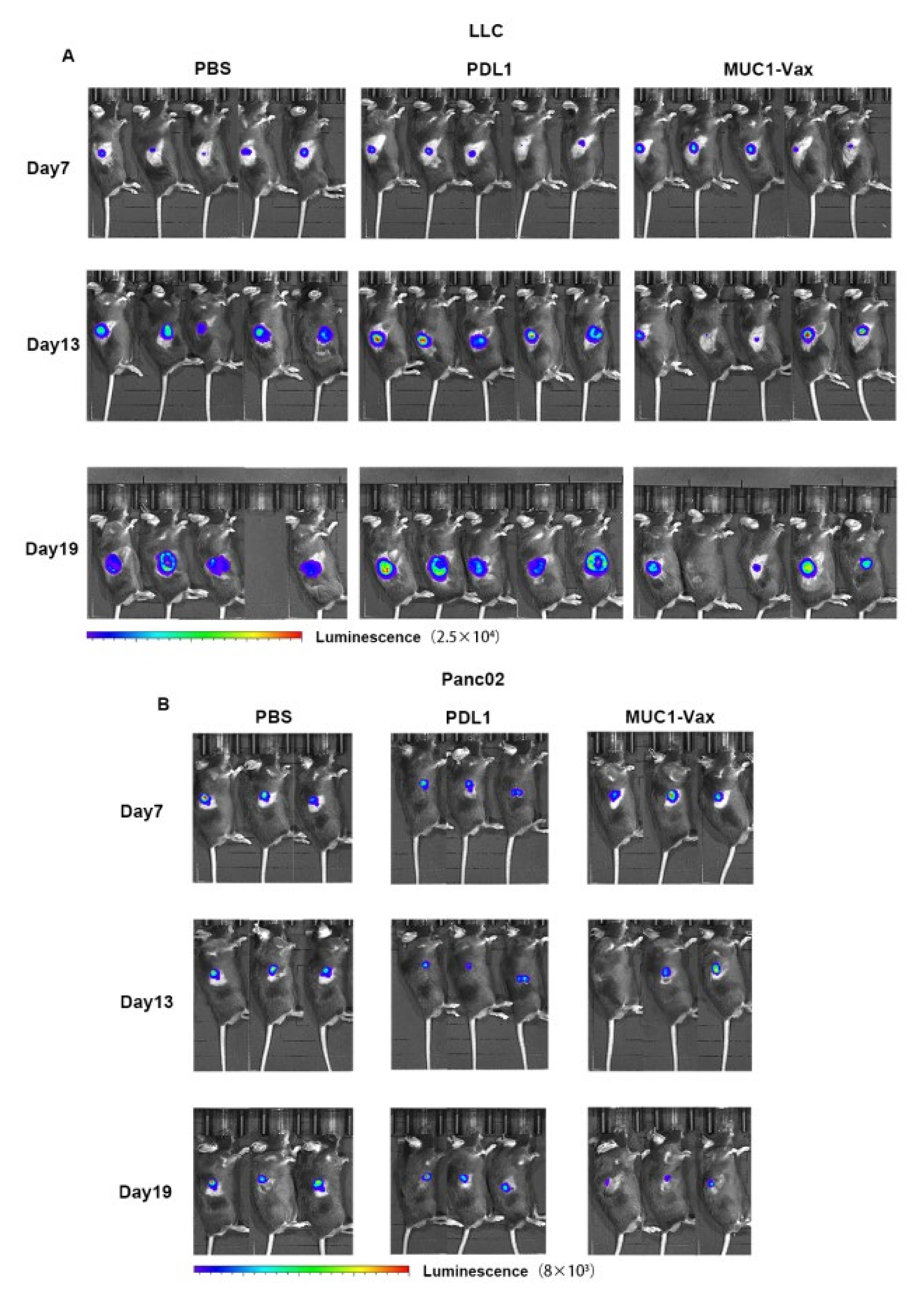
3.4. Inhibitory Effect of MUC1-Vax-DC Vaccine on Tumor Growth
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APCs | Antigen-presenting cells |
| CTL | Cytotoxic T lymphocyte |
| DCs | Dendritic cells |
| FACS | Fluorescence-activated cell sorting |
| FcγRs | Fcγ receptors |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| IFN | Interferon |
| IL | Interleukin |
| LPS | Lipopolysaccharide |
| mAb | Monoclonal antibody |
| MHC | Major histocompatibility complex |
| MUC1-Vax | MUC1-PDL1-IgG1 Fc fusion protein |
| PD-L1 | Programmed cell death ligand 1 |
| TAAs | Tumor-associated antigens |
| Th | helper T cell |
References
- Chen, D.S.; Mellman, I. Oncology meets immunology: The cancer-immunity cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keeffe, M.; Mok, W.H.; Radford, K.J. Human dendritic cell subsets and function in health and disease. Cell Mol. Life Sci. 2015, 72, 4309–4325. [Google Scholar] [PubMed]
- Lu, Y.; Shi, Y.; You, J. Strategy and clinical application of up-regulating cross presentation by DCs in anti-tumor therapy. J. Control. Release 2021, 341, 184–205. [Google Scholar] [CrossRef] [PubMed]
- Bol, K.F.; Schreibelt, G.; Gerritsen, W.R.; de Vries, I.J.M.; Figdor, C.G. Dendritic Cell–Based Immunotherapy: State of the Art and Beyond. Clin. Cancer Res. 2016, 22, 1897–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wculek, S.K.; Cueto, F.J.; Mujal, A.M.; Melero, I.; Krummel, M.F.; Sancho, D. Dendritic cells in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2020, 20, 7–24. [Google Scholar] [CrossRef]
- Wang, Y.; Xiang, Y.; Xin, V.W.; Wang, X.-W.; Peng, X.-C.; Liu, X.-Q.; Wang, D.; Li, N.; Cheng, J.-T.; Lyv, Y.-N.; et al. Dendritic cell biology and its role in tumor immunotherapy. J. Hematol. Oncol. 2020, 13, 107. [Google Scholar] [CrossRef]
- Li, B.; Hu, L. Cross-presentation of Exogenous Antigens. Transfus. Clin. Biol. 2019, 26, 346–351. [Google Scholar] [CrossRef]
- Macri, C.; Morgan, H.; Villadangos, J.A.; Mintern, J.D. Regulation of dendritic cell function by Fc-gamma-receptors and the neonatal Fc receptor. Mol. Immunol. 2021, 139, 193–201. [Google Scholar] [CrossRef]
- Hossain, M.K.; Wall, K.A. Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses. Cancers 2019, 11, 418. [Google Scholar] [CrossRef] [Green Version]
- Rudnik-Jansen, I.; Howard, K.A. FcRn expression in cancer: Mechanistic basis and therapeutic opportunities. J. Control. Release 2021, 337, 248–257. [Google Scholar] [CrossRef]
- Beckwith, D.M.; Cudic, M. Tumor-associated O-glycans of MUC1: Carriers of the glyco-code and targets for cancer vaccine design. Semin. Immunol. 2020, 47, 101389. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Papadimitriou, J.; Burchell, J.M.; Graham, R.; Beatson, R. Latest developments in MUC1 immunotherapy. Biochem. Soc. Trans. 2018, 46, 659–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Zhang, Z.; Zhang, S.; Zhu, P.; Ko, J.; Yung, K. MUC1: Structure, Function, and Clinic Application in Epithelial Cancers. Int. J. Mol. Sci. 2021, 22, 6567. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Cen, Q.; Lei, H. A review on development of MUC1-based cancer vaccine. Biomed. Pharmacother. 2020, 132, 110888. [Google Scholar] [CrossRef]
- Cheever, M.A.; Allison, J.P.; Ferris, A.S.; Finn, O.J.; Hastings, B.M.; Hecht, T.T.; Mellman, I.; Prindiville, S.A.; Viner, J.L.; Weiner, L.M.; et al. The Prioritization of Cancer Antigens: A National Cancer Institute Pilot Project for the Acceleration of Translational Research. Clin. Cancer Res. 2009, 15, 5323–5337. [Google Scholar] [CrossRef] [Green Version]
- Shindo, Y.; Hazama, S.; Maeda, Y.; Matsui, H.; Iida, M.; Suzuki, N.; Yoshimura, K.; Ueno, T.; Yoshino, S.; Sakai, K.; et al. Adoptive immunotherapy with MUC1-mRNA transfected dendritic cells and cytotoxic lymphocytes plus gemcitabine for unresectable pancreatic cancer. J. Transl. Med. 2014, 12, 175. [Google Scholar] [CrossRef] [Green Version]
- Loveland, B.E.; Zhao, A.; White, S.; Gan, H.; Hamilton, K.; Xing, P.-X.; Pietersz, G.A.; Apostolopoulos, V.; Vaughan, H.; Karanikas, V.; et al. Mannan-MUC1–Pulsed Dendritic Cell Immunotherapy: A Phase I Trial in Patients with Adenocarcinoma. Clin. Cancer Res. 2006, 12, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Yi, M.; Niu, M.; Xu, L.; Luo, S.; Wu, K. Regulation of PD-L1 expression in the tumor microenvironment. J. Hematol. Oncol. 2021, 14, 10. [Google Scholar] [CrossRef]
- Han, Y.Y.; Liu, D.D.; Li, L.H. PD-1/PD-L1 pathway: Current researche in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar]
- Chen, J.; Liu, H.; Jehng, T.; Li, Y.; Chen, Z.; Lee, K.-D.; Shen, H.-T.; Jones, L.; Huang, X.F.; Chen, S.-Y. A Novel Anti-PD-L1 Vaccine for Cancer Immunotherapy and Immunoprevention. Cancers 2019, 11, 1909. [Google Scholar] [CrossRef] [Green Version]
- Munir, S.; Andersen, G.H.; Woetmann, A.; Ødum, N.; Becker, J.C.; Andersen, M.H. Cutaneous T cell lymphoma cells are targets for immune checkpoint ligand PD-L1-specific, cytotoxic T cells. Leukemia 2013, 27, 2251–2253. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.M.; Larsen, S.K.; Svane, I.M.; Andersen, M.H. Harnessing PD-L1-specific cytotoxic T cells for anti-leukemia immunotherapy to defeat mechanisms of immune escape mediated by the PD-1 pathway. Leukemia 2013, 28, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.M.; Svane, I.M.; Andersen, M.H. The stimulation of PD-L1-specific cytotoxic T lymphocytes can both directly and indirectly enhance antileukemic immunity. Blood Cancer J. 2014, 4, e230. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.M.; Borch, T.H.; Hansen, M.; Andersen, M.H. PD-L1-specific T cells. Cancer Immunol. Immunother. 2016, 65, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.-N.; Han, S.-B.; Hwang, I.-Y.; Kehrl, J.H. B Cells Productively Engage Soluble Antigen-Pulsed Dendritic Cells: Visualization of Live-Cell Dynamics of B Cell-Dendritic Cell Interactions. J. Immunol. 2005, 175, 7125–7134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruprecht, C.R.; Lanzavecchia, A. Toll-like receptor stimulation as a third signal required for activation of human naive B cells. Eur. J. Immunol. 2006, 36, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Evel-Kabler, K.; Strube, R.; Chen, S.-Y. Silencing of SOCS1 enhances antigen presentation by dendritic cells and antigen-specific anti-tumor immunity. Nat. Biotechnol. 2004, 22, 1546–1553. [Google Scholar] [CrossRef]
- Beatson, R.E.; Taylor-Papadimitriou, J.; Burchell, J.M. MUC1 immunotherapy. Immunotherapy 2010, 2, 305–327. [Google Scholar] [CrossRef]
- Cibrián, D.; Sánchez-Madrid, F. CD69: From activation marker to metabolic gatekeeper. Eur. J. Immunol. 2017, 47, 946–953. [Google Scholar] [CrossRef]
- Dredge, K.; Marriott, B.J.; Todryk, S.M.; Dalgleish, A.G. Adjuvants and the promotion of Th1-type cytokines in tumour immunotherapy. Cancer Immunol. Immunother. 2002, 51, 521–531. [Google Scholar] [CrossRef]
- Knutson, K.L.; Disis, M.L. Tumor antigen-specific T helper cells in cancer immunity and immunotherapy. Cancer Immunol. Immunother. 2005, 54, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.M. T Cells: Soldiers and Spies—The Surveillance and Control of Effector T Cells by Regulatory T Cells. Clin. J. Am. Soc. Nephrol. 2015, 10, 2050–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacPherson, G.; Kushnir, N.; Wykes, M. Dendritic cells, B cells and the regulation of antibody synthesis. Immunol. Rev. 1999, 172, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Balázs, M.; Martin, F.; Zhou, T.; Kearney, J.F. Blood Dendritic Cells Interact with Splenic Marginal Zone B Cells to Initiate T-Independent Immune Responses. Immunity 2002, 17, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Sabado, R.L.; Balan, S.; Bhardwaj, N. Dendritic cell-based immunotherapy. Cell Res. 2017, 27, 74–95. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Zhou, L.; Mi, Q.-S.; Jiang, A. DC-Based Vaccines for Cancer Immunotherapy. Vaccines 2020, 8, 706. [Google Scholar] [CrossRef]
- Perez, C.R.; De Palma, M. Engineering dendritic cell vaccines to improve cancer immunotherapy. Nat. Commun. 2019, 10, 5408. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, M.; Nie, H.; Yuan, Y. PD-1 and PD-L1 in cancer immunotherapy: Clinical implications and future considerations. Hum. Vaccines Immunother. 2019, 15, 1111–1122. [Google Scholar] [CrossRef]
- Gong, J.; Chehrazi-Raffle, A.; Reddi, S.; Salgia, R. Development of PD-1 and PD-L1 inhibitors as a form of cancer immunotherapy: A comprehensive review of registration trials and future considerations. J. Immunother. Cancer 2018, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, I.; Vinuesa, C. Dendritic cells, BAFF, and APRIL: Innate players in adaptive antibody responses. Immunity 2002, 17, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Bi, Y.; Xue, L.; Zhang, Y.; Yang, H.; Chen, X.; Lu, Y.; Zhang, Z.; Liu, H.; Wang, X.; et al. Dendritic cell SIRT1-HIF1alpha axis programs the differentiation of CD4+ T cells through IL-12 and TGF-beta1. Proc. Natl. Acad. Sci. USA 2015, 112, E957–E965. [Google Scholar] [PubMed] [Green Version]
- van Gulijk, M.; Dammeijer, F.; Aerts, J.; Vroman, H. Combination Strategies to Optimize Efficacy of Dendritic Cell-Based Immunotherapy. Front Immunol. 2018, 9, 2759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Paulete, A.R.; Cueto, F.J.; Martínez-López, M.; Labiano, S.; Morales-Kastresana, A.; Rodriguez-Ruiz, M.E.; Jure-Kunkel, M.; Azpilikueta, A.; Aznar, M.A.; Quetglas, J.I.; et al. Cancer Immunotherapy with Immunomodulatory Anti-CD137 and Anti-PD-1 Monoclonal Antibodies Requires BATF3-Dependent Dendritic Cells. Cancer Discov. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlom, J. Therapeutic Cancer Vaccines: Current Status and Moving Forward. JNCI: J. Natl. Cancer Inst. 2012, 104, 599–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teitz-Tennenbaum, S.; Li, Q.; Davis, M.A.; Wilder-Romans, K.; Hoff, J.; Li, M.; Chang, A.E. Radiotherapy Combined with Intratumoral Dendritic Cell Vaccination Enhances the Therapeutic Efficacy of Adoptive T-cell Transfer. J. Immunother. 2009, 32, 602–612. [Google Scholar] [CrossRef] [Green Version]
- Ilieva, K.M.; Correa, I.; Josephs, D.H.; Karagiannis, P.; Egbuniwe, I.U.; Cafferkey, M.J.; Spicer, J.F.; Harries, M.; Nestle, F.O.; Lacy, K.E.; et al. Effects of BRAF Mutations and BRAF Inhibition on Immune Responses to Melanoma. Mol. Cancer Ther. 2014, 13, 2769–2783. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, J.; Zeng, W.; Jia, J.; Shi, Y.; Wang, D.; Dong, J.; Fang, Z.; He, J.; Yang, X.; Zhang, R.; et al. A Novel Therapeutic Tumor Vaccine Targeting MUC1 in Combination with PD-L1 Elicits Specific Anti-Tumor Immunity in Mice. Vaccines 2022, 10, 1092. https://doi.org/10.3390/vaccines10071092
Pan J, Zeng W, Jia J, Shi Y, Wang D, Dong J, Fang Z, He J, Yang X, Zhang R, et al. A Novel Therapeutic Tumor Vaccine Targeting MUC1 in Combination with PD-L1 Elicits Specific Anti-Tumor Immunity in Mice. Vaccines. 2022; 10(7):1092. https://doi.org/10.3390/vaccines10071092
Chicago/Turabian StylePan, Jiayi, Wuyi Zeng, Jiangtao Jia, Yi Shi, Danni Wang, Jun Dong, Zixuan Fang, Jiashan He, Xinyu Yang, Rong Zhang, and et al. 2022. "A Novel Therapeutic Tumor Vaccine Targeting MUC1 in Combination with PD-L1 Elicits Specific Anti-Tumor Immunity in Mice" Vaccines 10, no. 7: 1092. https://doi.org/10.3390/vaccines10071092
APA StylePan, J., Zeng, W., Jia, J., Shi, Y., Wang, D., Dong, J., Fang, Z., He, J., Yang, X., Zhang, R., He, M., Huang, M., Fu, B., Zhong, B., & Liu, H. (2022). A Novel Therapeutic Tumor Vaccine Targeting MUC1 in Combination with PD-L1 Elicits Specific Anti-Tumor Immunity in Mice. Vaccines, 10(7), 1092. https://doi.org/10.3390/vaccines10071092