
Evaluation of the Immunomodulatory Effect of the Recombinant 14-3-3 and Major Antigen Proteins of Strongyloides stercoralis against an Infection by S. venezuelensis

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statements
2.2. S. venezuelensis and Experimental Mice Model
2.3. Cloning of S. stercoralis Recombinant 14-3-3 and Major Antigen Proteins
2.4. Expression and Purification of rSs14-3-3 and rSsMA Proteins in E. coli
2.5. Vaccination Experiment Protocol
2.6. Detection of Antibodies in Sera by Indirect ELISA
2.7. RNA Isolation, Reverse Transcription and Quantification of Cytokine mRNAs
2.8. Statistical Analysis
3. Results
3.1. Cloning, Expression and Purification of Recombinant Ss14-3-3 and SsMA Proteins from S. stercoralis
3.2. Immunization with rSs14-3-3 and rSsMA Proteins Do Not Induce Effective Protection against the Experimental Infection
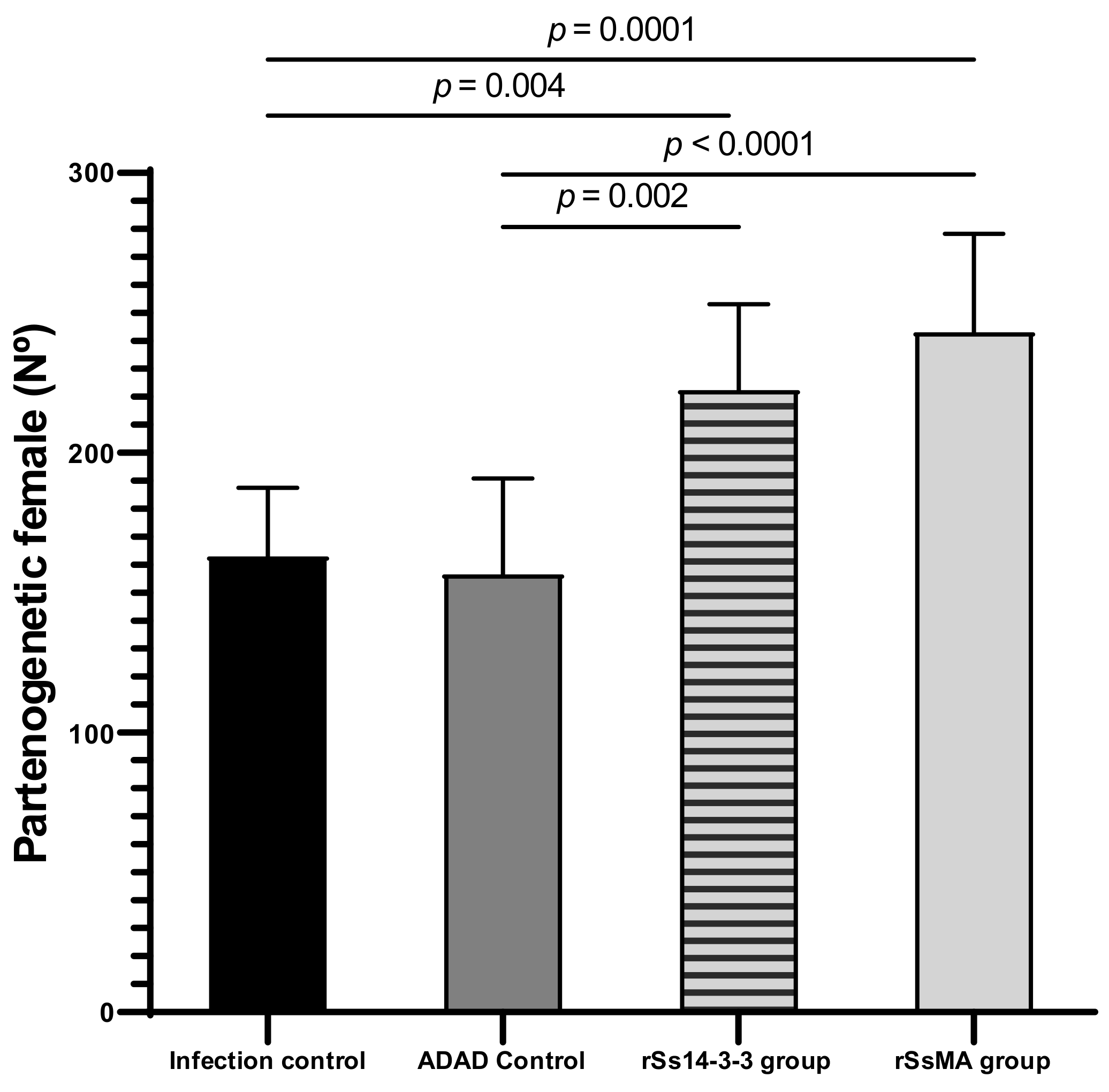
3.3. Splenic and Intestinal Indexes Reflect the Non-Protection of the Vaccine against Infection
3.4. Humoral Antibody Response to the Immunization of Mice with rSs14-3-3 and rSsMA Proteins and/or Infection with S. venezuelensis
3.5. Cytokine Profile in Mice after Immunization with rSs14-3-3 and rSsMA and Experimental Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Soil-Transmitted Helminth Infections. Newsroom. 10 January 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/soil-transmitted-helminth-infections (accessed on 1 June 2022).
- Olsen, A.; van Lieshout, L.; Marti, H.; Polderman, T.; Polman, K.; Steinmann, P.; Stothard, R.; Thybo, S.; Verweij, J.; Magnussen, P. Strongyloidiasis—The most neglected of the neglected tropical diseases? Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 967–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aramendia, A.A.; Anegagrie, M.; Zewdie, D.; Dacal, E.; Saugar, J.M.; Herrador, Z.; Hailu, T.; Yimer, M.; Periago, M.V.; Rodriguez, E.; et al. Epidemiology of intestinal helminthiases in a rural community of Ethiopia: Is it time to expand control programs to include Strongyloides stercoralis and the entire community? PLoS Negl. Trop. Dis. 2020, 14, e0008315. [Google Scholar] [CrossRef] [PubMed]
- Botero, D.; Restrepo, M. Parasitosis Humanas, 5th ed.; Corporación para Investigaciones Biológicas: Medellín, Colombia, 2012; pp. 162–177. [Google Scholar]
- Igual Adell, R.; Domínguez Márquez, V. Estrongiloidiasis: Epidemiología, manifestaciones clínicas y diagnóstico. Experiencia en una zona endémica: La comarca de La Safor (Valencia). Enferm. Infecc. Microb. Clin. 2007, 25, 38–44. [Google Scholar] [CrossRef]
- Viney, M.; Kikuchi, T. Strongyloides ratti and S. venezuelensis—Rodent models of Strongyloides infection. Parasitology 2017, 144, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruano, A.L.; Martín, T.; Pardo, J.; López-Abán, J.; Muro, A. Avances en el estudio sobre la estrongiloidosis. Enferm. Emerg. 2005, 7, 102–109. Available online: http://enfermedadesemergentes.com/articulos/a414/s-7-2-003.pdf (accessed on 2 February 2022).
- Lucas Dato, A.; Pacheco-Tenza, M.I.; Borrajo Brunete, E.; Martínez López, B.; García López, M.; González Cuello, I.; Gregori Colomé, J.; Navarro Cots, M.; Saugar, J.M.; García-Vazquez, E.; et al. Strongyloidiasis in Southern Alicante (Spain): Comparative Retrospective Study of Autochthonous and Imported Cases. Pathogens 2020, 9, 601. [Google Scholar] [CrossRef]
- Barroso, M.; Salvador, F.; Sánchez-Montalvá, A.; Bosch-Nicolau, P.; Molina, I. Strongyloides stercoralis infection: A systematic review of endemic cases in Spain. PLoS Negl. Trop. Dis. 2019, 13, e0007230. [Google Scholar] [CrossRef]
- Martínez-Vázquez, C.; González Mediero, G.; Núñez, M.; Pérez, S.; García-Fernández, J.M.; Gimena, B. Strongyloides stercoralis en el sur de Galicia. Ann. Med. Interna. 2003, 20, 39–41. Available online: http://scielo.isciii.es/scielo.php?script=sci_arttext&pid=S0212-71992003000900008&lng=es (accessed on 1 June 2022). [CrossRef] [Green Version]
- Román-Sánchez, P.; Pastor-Guzmán, A.; Moreno-Guillén, S.; Igual-Adell, R.; Suñer-Generoso, S.; Tornero-Estébanez, C. High prevalence of Strongyloides stercoralis among farm workers on the mediterranean coast of Spain: Analysis of the predictive factors of infection in developed countries. Am. J. Trop. Med. Hyg. 2003, 69, 336–340. [Google Scholar] [CrossRef]
- Alcaraz, C.O.; Adell, R.I.; Sánchez, P.S.; Blasco, M.J.; Sánchez, O.A.; Auñón, A.S.; Calabuig, D.R. Characteristics and geographical profile of strongyloidiasis in healthcare area 11 of the Valencian community (Spain). J. Infect. 2004, 49, 152–158. [Google Scholar] [CrossRef]
- Marcilla, A.; Sotillo, J.; Pérez-Garcia, A.; Igual-Adell, R.; Valero, M.L.; Sánchez-Pino, M.M.; Bernal, D.; Muñoz-Antolí, C.; Trelis, M.; Toledo, R.; et al. Proteomic analysis of Strongyloides stercoralis L3 larvae. Parasitology 2010, 137, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Marcilla, A.; Garg, G.; Bernal, D.; Ranganathan, S.; Forment, J.; Ortiz, J.; Muñoz-Antolí, C.; Dominguez, M.V.; Pedrola, L.; Martinez-Blanch, J.; et al. The transcriptome analysis of Strongyloides stercoralis L3i larvae reveals targets for intervention in a neglected disease. PLoS Negl. Trop. Dis. 2012, 6, e1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siles-Lucas, M.; Gottstein, B. The 14-3-3 protein: A key molecule in parasites as in other organisms. Trends Parasitol. 2003, 19, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Rodpai, R.; Intapan, P.M.; Thanchomnang, T.; Sanpool, O.; Janwan, P.; Laummaunwai, P.; Wongkham, C.; Insawang, T.; Maleewong, W. Strongyloides stercoralis diagnostic polypeptides for human strongyloidiasis and their proteomic analysis. Parasitol. Res. 2016, 115, 4007–4012. [Google Scholar] [CrossRef] [PubMed]
- Rodpai, R.; Intapan, P.M.; Thanchomnang, T.; Sanpool, O.; Janwan, P.; Laummaunwai, P.; Wongkham, C.; Insawang, T.; Maleewong, W. Identification of antigenic proteins in Strongyloides stercoralis by proteomic analysis. Parasitol. Res. 2017, 116, 1687–1693. [Google Scholar] [CrossRef]
- Vicente, B.; López-Abán, J.; Rojas-Caraballo, J.; Pérez del Villar, L.; Hillyer, G.V.; Martínez-Fernández, A.R.; Muro, A. A Fasciola hepatica-derived fatty acid binding protein induces protection against schistosomiasis caused by Schistosoma bovis using the adjuvant adaptation (ADAD) vaccination system. Exp. Parasitol. 2014, 145, 145–151. [Google Scholar] [CrossRef]
- Vicente, B.; López-Abán, J.; Rojas-Caraballo, J.; del Olmo, E.; Fernández-Soto, P.; Muro, A. Protection against Schistosoma mansoni infection using a Fasciola hepatica-derived fatty acid binding protein from different delivery systems. Parasites Vector 2016, 9, 216. [Google Scholar] [CrossRef] [Green Version]
- Legarda-Ceballos, A.L.; López-Abán, J.; Del Olmo, E.; Escarcena, R.; Bustos, L.A.; Rojas-Caraballo, J.; Vicente, B.; Fernández-Soto, P.; San Feliciano, A.; Muro, A. In vitro and in vivo evaluation of 2-aminoalkanol and 1,2-alkanediamine derivatives against Strongyloides venezuelensis. Parasites Vectors 2016, 9, 364. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Fernández, A.R.; Nogal-Ruiz, J.J.; López-Abán, J.; Ramajo, V.; Oleaga, A.; Manga-González, Y.; Hillyer, G.V.; Muro, A. Vaccination of mice and sheep with Fh12 FABP from Fasciola hepatica using the new adjuvant/immunomodulator system ADAD. Vet. Parasitol. 2004, 126, 287–298. [Google Scholar] [CrossRef]
- Vlaminck, J.; López-Abán, J.; Ruano, A.L.; del Olmo, E.; Muro, A. Vaccination against Strongyloides venezuelensis with homologue antigens using new immunomodulators. J. Parasitol. 2010, 96, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Ye, Z.; Chang, Y.; Huang, S.; Song, J.; Lu, F. Trichinella spiralis co-infection exacerbates Plasmodium berghei malaria-induced hepatopathy. Parasites Vectors 2020, 13, 440. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Manente, F.A.; Quinello, C.; Ferreira, L.S.; de Andrade, C.R.; Jellmayer, J.A.; Portuondo, D.L.; Batista-Duharte, A.; Carlos, I.Z. Experimental sporotrichosis in a cyclophosphamide-induced immunosuppressed mice model. Med. Mycol. 2018, 56, 711–722. [Google Scholar] [CrossRef]
- Karanam, L.S.K.; Basavraj, G.K.; Papireddy, C.K.R. Strongyloides stercoralis Hyper infection Syndrome. Indian J. Surg. 2020, 12, 1–5. [Google Scholar] [CrossRef]
- Czeresnia, J.M.; Weiss, L.M. Strongyloides stercoralis . Lung 2022, 200, 141–148. [Google Scholar] [CrossRef]
- Perera, D.; Ndao, M. Promising Technologies in the Field of Helminth Vaccines. Front. Immunol. 2021, 12, 711650. [Google Scholar] [CrossRef]
- Culma, M.F. Strongyloides stercoralis proteome: A reverse approach to the identification of potential immunogenic candidates. Microb. Pathog. 2021, 152, 104545. [Google Scholar] [CrossRef]
- Siles-Lucas, M.; Uribe, N.; López-Abán, J.; Vicente, B.; Orfao, A.; Nogal-Ruiz, J.J.; Feliciano, A.S.; Muro, A. The Schistosoma bovis Sb14-3-3zeta recombinant protein cross-protects against Schistosoma mansoni in BALB/c mice. Vaccine 2007, 25, 7217–7223. [Google Scholar] [CrossRef]
- Rojas-Caraballo, J.; López-Abán, J.; Pérez del Villar, L.; Vizcaíno, C.; Vicente, B.; Fernández-Soto, P.; del Olmo, E.; Patarroyo, M.A.; Muro, A. In vitro and in vivo studies for assessing the immune response and protection-inducing ability conferred by Fasciola hepatica-derived synthetic peptides containing B- and T-cell epitopes. PLoS ONE 2014, 9, e105323. [Google Scholar] [CrossRef] [Green Version]
- Trelis, M.; Galiano, A.; Bolado, A.; Toledo, R.; Marcilla, A.; Bernal, D. Subcutaneous injection of exosomes reduces symptom severity and mortality induced by Echinostoma caproni infection in BALB/c mice. Int. J. Parasitol. 2016, 46, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Molehin, A.J.; Gray, S.A.; Turner, C.; Davis, J.; Zhang, W.; Khatoon, S.; Rattan, M.; Kernen, R.; Peterson, C.; Sennoune, S.R.; et al. Process Development of Sj-p80: A Low-Cost Transmission-Blocking Veterinary Vaccine for Asiatic Schistosomiasis. Front. Immunol. 2021, 11, 578715. [Google Scholar] [CrossRef] [PubMed]
- Gause, W.C.; Wynn, T.A.; Allen, J.E. Type 2 immunity and wound healing: Evolutionary refinement of adaptive immunity by helminths. Nat. Rev. Immunol. 2013, 13, 607–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, J.E.; Sutherland, T.E. Host protective roles of type 2 immunity: Parasite killing and tissue repair, flip sides of the same coin. Semin. Immunol. 2014, 26, 329–340. [Google Scholar] [CrossRef] [PubMed]
- McSorley, H.J.; Chayé, M.A.M.; Smits, H.H. Worms: Pernicious parasites or allies against allergies? Parasite Immunol. 2019, 41, e12574. [Google Scholar] [CrossRef]
- Maizels, R.M.; Yazdanbakhsh, M. Immune regulation by helminth parasites: Cellular and molecular mechanisms. Nat. Rev. Immunol. 2003, 3, 733–744. [Google Scholar] [CrossRef] [Green Version]
- Behnke, J.M.; Barnard, C.J.; Wakelin, D. Understanding Chronic Nematode Infections: Evolutionary Considerations, Current Hypotheses and The Way Forward. Int. J. Parasitol. 1992, 22, 861–907. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, R.; Dincer, H.E.; Whittemore, D. Strongyloides stercoralis hyperinfection in a patient with rheumatoid arthritis after anti-TNF-α therapy. J. Clin. Rheumatol. 2007, 13, 150–152. [Google Scholar] [CrossRef]
- Artis, D.; Humphreys, N.E.; Bancroft, A.J.; Rothwell, N.J.; Potten, C.S.; Grencis, R.K. Tumor Necrosis Factor Is a Critical Component of Interleukin 13-mediated Protective T Helper Cell Type 2 Responses during Helminth Infection. J. Exp. Med. 1999, 190, 953–962. [Google Scholar] [CrossRef] [Green Version]
- Khaliq, M.F.; Ihle, R.E.; Perry, J. Immunosuppression with Antitumour Necrosis Factor Therapy Leading to Strongyloides Hyperinfection Syndrome. Case Rep. Infect Dis. 2018, 2018, 6341680. [Google Scholar] [CrossRef]
- Prior-Español, Á.; Roure, S.; Mateo, L.; Martínez-Morillo, M. Anaemia and Eosinophilia Secondary to Strongyloides in a Patient Treated with Anti-TNF-α. Reumatol. Clin. 2020, 16, 508–509. [Google Scholar] [CrossRef] [PubMed]
- Masoori, L.; Falak, R.; Mokhtarian, K.; Bandehpour, M.; Razmjou, E.; Jalallou, N.; Jafarian, F.; Akhlaghi, L.; Meamar, A.R. Production of recombinant 14-3-3 protein and determination of its immunogenicity for application in serodiagnosis of strongyloidiasis. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rivas, G.; Rivaya, B.; Romaní, N.; Wang, J.; Alcaide, M.; Matas, L. Diagnosis of soil-transmitted helminth infections. An unsolved problem in the omics era. Enferm. Infecc. Microbiol. Clin. 2019, 37, 20–25. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Palencia, L.F.; Trelis, M.; López-Abán, J.; Galiano, A.; Vicente, B.; del Olmo, E.; Muro, A.; Bernal, D.; Marcilla, A. Evaluation of the Immunomodulatory Effect of the Recombinant 14-3-3 and Major Antigen Proteins of Strongyloides stercoralis against an Infection by S. venezuelensis. Vaccines 2022, 10, 1292. https://doi.org/10.3390/vaccines10081292
Sánchez-Palencia LF, Trelis M, López-Abán J, Galiano A, Vicente B, del Olmo E, Muro A, Bernal D, Marcilla A. Evaluation of the Immunomodulatory Effect of the Recombinant 14-3-3 and Major Antigen Proteins of Strongyloides stercoralis against an Infection by S. venezuelensis. Vaccines. 2022; 10(8):1292. https://doi.org/10.3390/vaccines10081292
Chicago/Turabian StyleSánchez-Palencia, Liz F., María Trelis, Julio López-Abán, Alicia Galiano, Belén Vicente, Esther del Olmo, Antonio Muro, Dolores Bernal, and Antonio Marcilla. 2022. "Evaluation of the Immunomodulatory Effect of the Recombinant 14-3-3 and Major Antigen Proteins of Strongyloides stercoralis against an Infection by S. venezuelensis" Vaccines 10, no. 8: 1292. https://doi.org/10.3390/vaccines10081292
APA StyleSánchez-Palencia, L. F., Trelis, M., López-Abán, J., Galiano, A., Vicente, B., del Olmo, E., Muro, A., Bernal, D., & Marcilla, A. (2022). Evaluation of the Immunomodulatory Effect of the Recombinant 14-3-3 and Major Antigen Proteins of Strongyloides stercoralis against an Infection by S. venezuelensis. Vaccines, 10(8), 1292. https://doi.org/10.3390/vaccines10081292