

Quasispecies Nature of RNA Viruses: Lessons from the Past


Abstract
:1. Preamble
1.1. Origin of Quasispecies
1.2. Contributors of Quasispecies Formation
2. Unique Features of the RNA Genome
3. Footprints of Quasispecies Evolution of Major RNA Viruses
3.1. Paramyxoviridae
3.2. Orthomyxoviridae
3.2.1. Avian Influenza
3.2.2. Mutations in Influenza Leading toward Quasispecies
3.3. Reteroviridae
3.4. Coronaviridae
3.5. Other Viruses
3.5.1. Hepatitis Virus
3.5.2. Ebolavirus
3.5.3. Poliovirus
3.5.4. Chikungunya Virus
4. Challenges and Discussions for Prospective Antiviral Treatments
5. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eigen, M. New concepts for dealing with the evolution of nucleic acids. Cold Spring Harb. Symp. Quant. Biol. 1987, 52, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Eigen, M. On the nature of virus quasispecies. Trends Microbiol. 1996, 4, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Eigen, M.; Schuster, P. A principle of natural self-organization. Sci. Nat. 1977, 64, 541–565. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.A. What is a quasispecies? Trends Ecol. Evol. 1992, 7, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Boerlijst, M.C.; Bonhoeffer, S.; Nowak, M.A. Viral quasi-species and recombination. Proc. R. Soc. B Biol. Sci. 1996, 263, 1577–1584. [Google Scholar]
- Jacobi, M.N.; Nordahl, M. Quasispecies and recombination. Theor. Popul. Biol. 2006, 70, 479–485. [Google Scholar] [CrossRef]
- Domingo, E.; Perales, C. Viral quasispecies. PLoS Genet. 2019, 15, e1008271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andino, R.; Domingo, E. Viral quasispecies. Virology 2015, 479, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauring, A.S.; Andino, R. Quasispecies theory and the behavior of RNA viruses. PLoS Patog. 2010, 6, e1001005. [Google Scholar] [CrossRef] [Green Version]
- Eigen, M. Selforganization of matter and the evolution of biological macromolecules. Sci. Nat. 1971, 58, 465–523. [Google Scholar] [CrossRef]
- Domingo, E.; García-Crespo, C.; Perales, C. Historical perspective on the discovery of the quasispecies concept. Annu. Rev. Virol. 2021, 8, 51–72. [Google Scholar] [CrossRef]
- Domingo, E.; Holland, J. RNA virus mutations and fitness for survival. Annu. Rev. Microbiol. 1997, 51, 151. [Google Scholar] [CrossRef] [PubMed]
- Ojosnegros, S.; Perales, C.; Mas, A.; Domingo, E. Quasispecies as a matter of fact: Viruses and beyond. Virus Res. 2011, 162, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Sheldon, J.; Perales, C. Viral quasispecies evolution. Microbiol. Mol. Biol. Rev. 2012, 76, 159–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingo, E.; Perales, C. Quasispecies and virus. Eur. Biophys. J. 2018, 47, 443–457. [Google Scholar] [CrossRef]
- Agol, V.I.; Gmyl, A.P. Emergency services of viral RNAs: Repair and remodeling. Microbiol. Mol. Biol. Rev. 2018, 82, e00067-17. [Google Scholar] [CrossRef] [Green Version]
- Sanjuán, R.; Nebot, M.R.; Chirico, N.; Mansky, L.M.; Belshaw, R.J. Viral mutation rates. J. Virol. 2010, 84, 9733–9748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttle, C.A. Marine viruses—Major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef]
- Elde, N.C.; Child, S.J.; Geballe, A.P.; Malik, H.S. Protein kinase R reveals an evolutionary model for defeating viral mimicry. Nature 2009, 457, 485–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enard, D.; Cai, L.; Gwennap, C.; Petrov, D.A. Viruses are a dominant driver of protein adaptation in mammals. eLife 2016, 5, e12469. [Google Scholar] [CrossRef] [PubMed]
- Tenoever, B.R.; Ng, S.-L.; Chua, M.A.; McWhirter, S.M.; García-Sastre, A.; Maniatis, T. Multiple functions of the IKK-related kinase IKKε in interferon-mediated antiviral immunity. Sciences 2007, 315, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Suspene, R.; Renard, M.; Henry, M.; Guétard, D.; Puyraimond-Zemmour, D.; Billecocq, A.; Bouloy, M.; Tangy, F.; Vartanian, J.-P.; Wain-Hobson, S. Inversing the natural hydrogen bonding rule to selectively amplify GC-rich ADAR-edited RNAs. Nucleic Acids Res. 2008, 36, e72. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.V.; George, C.X.; Welch, M.J.; Liou, L.-Y.; Hahm, B.; Lewicki, H.; de la Torre, J.C.; Samuel, C.E.; Oldstone, M.B. RNA editing enzyme adenosine deaminase is a restriction factor for controlling measles virus replication that also is required for embryogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 331–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattaneo, R.; Schmid, A.; Eschle, D.; Baczko, K.; ter Meulen, V.; Billeter, M.A. Biased hypermutation and other genetic changes in defective measles viruses in human brain infections. Cell 1988, 55, 255–265. [Google Scholar] [CrossRef]
- Domingo, E.; Martin, V.; Perales, C.; Grande-Perez, A.; Garcia-Arriaza, J.; Arias, A. Viruses as quasispecies: Biological implications. In Quasispecies: Concept and Implications for Virology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 51–82. [Google Scholar]
- Ali, A.; Roossinck, M. Analysis of quasispecies variation in single and mixed viral infection. Virus Evol. 2017, 3, vex037. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ruiz, H.; Diaz, A.; Ahlquist, P. Intermolecular RNA recombination occurs at different frequencies in alternate forms of brome mosaic virus RNA replication compartments. Viruses 2018, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Genet. 2011, 9, 617–626. [Google Scholar] [CrossRef]
- Tentchev, D.; Verdin, E.; Marchal, C.; Jacquet, M.; Aguilar, J.M.; Moury, B.J. Evolution and structure of Tomato spotted wilt virus populations: Evidence of extensive reassortment and insights into emergence processes. J. Gen. Virol. 2011, 92, 961–973. [Google Scholar] [CrossRef]
- Kwon, S.-J.; Cho, Y.-E.; Kwon, O.-H.; Kang, H.-G.; Seo, J.-K. Resistance-breaking tomato spotted wilt virus variant that recently occurred in pepper in South Korea is a genetic reassortant. Plant Dis. 2021, 105, 2771–2775. [Google Scholar] [CrossRef]
- Gu, H.; Zhang, C.; Ghabrial, S.A. Novel naturally occurring Bean pod mottle virus reassortants with mixed heterologous RNA1 genomes. Virology 2007, 97, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Pita, J.; Fondong, V.; Sangare, A.; Otim-Nape, G.; Ogwal, S.; Fauquet, C. Recombination, pseudorecombination and synergism of geminiviruses are determinant keys to the epidemic of severe cassava mosaic disease in Uganda. J. Gen. Virol. 2001, 82, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-I.; Si, Y.-J.; Kwon, H.-I.; Kim, E.-H.; Park, S.-J.; Robles, N.J.; Nguyen, H.D.; Yu, M.-A.; Yu, K.-M.; Lee, Y.-J. Pathogenicity and genetic characterisation of a novel reassortant, highly pathogenic avian influenza (HPAI) H5N6 virus isolated in Korea, 2017. Eurosurveillance 2018, 23, 18–00045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höfer, P.; Engel, M.; Jeske, H.; Frischmuth, T. Host range limitation of a pseudorecombinant virus produced by two distinct bipartite geminiviruses. MPMI 1997, 10, 1019–1022. [Google Scholar] [CrossRef] [Green Version]
- Idris, A.; Mills-Lujan, K.; Martin, K.; Brown, J. Melon chlorotic leaf curl virus: Characterization and differential reassortment with closest relatives reveal adaptive virulence in the squash leaf curl virus clade and host shifting by the host-restricted bean calico mosaic virus. J. Virol. 2008, 82, 1959–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolan, P.T.; Whitfield, Z.J.; Andino, R. Mechanisms and concepts in RNA virus population dynamics and evolution. Annu. Rev. Virol. 2018, 5, 69–92. [Google Scholar] [CrossRef] [PubMed]
- Loewe, L. Negative Selection. Nat. Educ. 2008, 1, 59. [Google Scholar]
- Schneider, W.L.; Roossinck, M.J. Genetic diversity in RNA virus quasispecies is controlled by host-virus interactions. J. Virol. 2001, 75, 6566–6571. [Google Scholar] [CrossRef] [Green Version]
- Stern, A.; Bianco, S.; Te Yeh, M.; Wright, C.; Butcher, K.; Tang, C.; Nielsen, R.; Andino, R. Costs and benefits of mutational robustness in RNA viruses. Cell Rep. 2014, 8, 1026–1036. [Google Scholar] [CrossRef] [Green Version]
- Sanjuán, R.; Domingo-Calap, P.J. Mechanisms of viral mutation. Cell. Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef] [Green Version]
- Bradwell, K.; Combe, M.; Domingo-Calap, P.; Sanjuán, R. Correlation between mutation rate and genome size in riboviruses: Mutation rate of bacteriophage Qβ. Genetics 2013, 195, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Mandary, M.B.; Masomian, M.; Poh, C.L. Impact of RNA virus evolution on quasispecies formation and virulence. Int. J. Mol. Sci. 2019, 20, 4657. [Google Scholar] [CrossRef] [Green Version]
- Aleem, A.; Samad, A.B.A.; Slenker, A.K. Emerging variants of SARS-CoV-2 and novel therapeutics against coronavirus (COVID-19). In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Kingsbury, D.W.; Bratt, M.A.; Choppin, P.W.; Hanson, R.P.; Hosaka, Y.; ter Meulen, V.; Norrby, E.; Plowright, W.; Rott, R.; Wunner, W.H. Paramyxoviridae. Intervirology 1978, 10, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Rima, B.; Balkema-Buschmann, A.; Dundon, W.G.; Duprex, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.; Lee, B.; Rota, P. ICTV virus taxonomy profile: Paramyxoviridae. J. Gen. Virol. 2019, 100, 1593. [Google Scholar] [CrossRef] [PubMed]
- Jordan, I.K.; Sutter IV, B.A.; McClure, M.A. Molecular evolution of the Paramyxoviridae and Rhabdoviridae multiple-protein-encoding P gene. Mol. Biol. Evol. 2000, 17, 75–086. [Google Scholar] [CrossRef] [PubMed]
- Dahlberg, J.E.; Simon, E.H. Physical and genetic studies of Newcastle disease virus: Evidence for multiploid particles. Virology 1969, 38, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.L.; Amarasinghe, G.K.; Bányai, K.; Bào, Y.; Basler, C.F.; Bavari, S.; Bejerman, N.; Blasdell, K.R.; Briand, F.-X.; Briese, T. Taxonomy of the order Mononegavirales: Update 2016. Arch. Virol. 2016, 161, 2351–2360. [Google Scholar] [CrossRef] [Green Version]
- Sullender, W.M. Respiratory syncytial virus genetic and antigenic diversity. Am. Soc. Microbiol. 2000, 13, 1–15. [Google Scholar] [CrossRef]
- Bastien, N.; Liu, L.; Ward, D.; Taylor, T.; Li, Y.J. Genetic variability of the G glycoprotein gene of human metapneumovirus. J. Clin. Microbiol. 2004, 42, 3532–3537. [Google Scholar] [CrossRef] [Green Version]
- Yoon, K.-J. 23 Overview of Viruses. In Diseases of Swine, 11th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Ma, W. Orthomyxoviridae. In Veterinary Microbiology; John Wiley & Sons: Hoboken, NJ, USA, 2022; pp. 573–588. [Google Scholar]
- Lofgren, E.; Fefferman, N.H.; Naumov, Y.N.; Gorski, J.; Naumova, E.N. Influenza seasonality: Underlying causes and modeling theories. J. Virol. 2007, 81, 5429–5436. [Google Scholar] [CrossRef] [Green Version]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Kim, H.; Webster, R.G.; Webby, R.J. Influenza virus: Dealing with a drifting and shifting pathogen. Viral Immunol. 2018, 31, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Sharp, G.B.; Claas, E.C. Interspecies transmission of influenza viruses. Am. J. Respir. Crit. Care Med. 1995, 152, S25. [Google Scholar] [CrossRef] [PubMed]
- Dugan, V.G.; Chen, R.; Spiro, D.J.; Sengamalay, N.; Zaborsky, J.; Ghedin, E.; Nolting, J.; Swayne, D.E.; Runstadler, J.A.; Happ, G.M. The evolutionary genetics and emergence of avian influenza viruses in wild birds. PLoS Pathog. 2008, 4, e1000076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-T.; Linster, M.; Mendenhall, I.H.; Su, Y.C.; Smith, G.J. Avian influenza viruses in humans: Lessons from past outbreaks. Br. Med Bull. 2019, 132, 81–95. [Google Scholar] [CrossRef]
- Chan, P.K. Outbreak of avian influenza A (H5N1) virus infection in Hong Kong in 1997. Clin. Infect. Dis. 2002, 34, S58–S64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morens, D.M.; Taubenberger, J.K.; Folkers, G.K.; Fauci, A.S. Pandemic influenza’s 500th anniversary. Clin. Infect. Dis. 2010, 51, 1442–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taubenberger, J.K.; Morens, D.M.; Fauci, A.S.J.J. The next influenza pandemic: Can it be predicted? JAMA 2007, 297, 2025–2027. [Google Scholar] [CrossRef] [Green Version]
- Worobey, M.; Han, G.-Z.; Rambaut, A. Genesis and pathogenesis of the 1918 pandemic H1N1 influenza A virus. Proc. Natl. Acad. Sci. USA 2014, 111, 8107–8112. [Google Scholar] [CrossRef] [Green Version]
- Reid, A.H.; Taubenberger, J.K.; Fanning, T.G. Evidence of an absence: The genetic origins of the 1918 pandemic influenza virus. Nat. Rev. Microbiol. 2004, 2, 909–914. [Google Scholar] [CrossRef]
- Johnson, N.P.; Mueller, J. Updating the accounts: Global mortality of the 1918–1920 “Spanish” influenza pandemic. Bull. Hist. Med. 2002, 76, 105–115. [Google Scholar] [CrossRef]
- Morens, D.M.; Taubenberger, J.K.; Fauci, A.S. The persistent legacy of the 1918 influenza virus. N. Engl. J. Med. 2009, 361, 225–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaoka, Y.; Krauss, S.; Webster, R.G.J. Avian-to-human transmission of the PB1 gene of influenza A viruses in the 1957 and 1968 pandemics. J. Virol. 1989, 63, 4603–4608. [Google Scholar] [CrossRef] [PubMed]
- Easterbrook, J.D.; Kash, J.C.; Sheng, Z.M.; Qi, L.; Gao, J.; Kilbourne, E.D.; Eichelberger, M.C.; Taubenberger, J.K. Immunization with 1976 swine H1N1-or 2009 pandemic H1N1-inactivated vaccines protects mice from a lethal 1918 influenza infection. Influenza 2011, 5, 198–205. [Google Scholar] [CrossRef]
- Simonsen, L.; Spreeuwenberg, P.; Lustig, R.; Taylor, R.J.; Fleming, D.M.; Kroneman, M.; Van Kerkhove, M.D.; Mounts, A.W.; Paget, W.J. Global mortality estimates for the 2009 Influenza Pandemic from the GLaMOR project: A modeling study. PLoS Med. 2013, 10, e1001558. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef] [Green Version]
- Sederdahl, B.K.; Williams, J.V. Epidemiology and clinical characteristics of influenza C virus. Viruses 2020, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Domingo, E.; Menéndez-Arias, L.; Holland, J. RNA virus fitness. Rev. Med. Virol. 1997, 7, 87–96. [Google Scholar] [CrossRef]
- Coffin, J.M.; Hughes, S.H.; Varmus, H.E. Retroviruses; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Kurth, R.; Bannert, N. Retroviruses: Molecular Biology, Genomics and Pathogenesis; Horizon Scientific Press: Norfolk, UK, 2010. [Google Scholar]
- Vargiu, L.; Rodriguez-Tomé, P.; Sperber, G.O.; Cadeddu, M.; Grandi, N.; Blikstad, V.; Tramontano, E.; Blomberg, J. Classification and characterization of human endogenous retroviruses; mosaic forms are common. Retrovirology 2016, 13, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, S.R. Cellular immune responses to retroviruses. In Retrovirus-Cell Interactions; Elsevier: Amsterdam, The Netherlands, 2018; pp. 401–420. [Google Scholar]
- Coffin, C.S.; Mulrooney-Cousins, P.M.; van Marle, G.; Roberts, J.P.; Michalak, T.I.; Terrault, N.A. Hepatitis B virus quasispecies in hepatic and extrahepatic viral reservoirs in liver transplant recipients on prophylactic therapy. Transplantation 2011, 17, 955–962. [Google Scholar] [CrossRef]
- Ho, D.D.; Neumann, A.U.; Perelson, A.S.; Chen, W.; Leonard, J.M.; Markowitz, M. Rapid turnover of plasma virions and CD4 lymphocytes in HIV-1 infection. Nature 1995, 373, 123–126. [Google Scholar] [CrossRef]
- Hu, W.-S.; Temin, H.M. Genetic consequences of packaging two RNA genomes in one retroviral particle: Pseudodiploidy and high rate of genetic recombination. Proc. Natl. Acad. Sci. USA 1990, 87, 1556–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jetzt, A.E.; Yu, H.; Klarmann, G.J.; Ron, Y.; Preston, B.D.; Dougherty, J.P. High rate of recombination throughout the human immunodeficiency virus type 1 genome. J. Virol. 2000, 74, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Briones, C.; Domingo, E. Minority report: Hidden memory genomes in HIV-1 quasispecies and possible clinical implications. AIDS Rev. 2008, 10, 93–109. [Google Scholar] [PubMed]
- Wade, J.; Sterjovski, J.; Gray, L.; Roche, M.; Chiavaroli, L.; Ellett, A.; Jakobsen, M.R.; Cowley, D.; da Fonseca Pereira, C.; Saksena, N. Enhanced CD4+ cellular apoptosis by CCR5-restricted HIV-1 envelope glycoprotein variants from patients with progressive HIV-1 infection. Virology 2010, 396, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; McLane, M.F.; Ratner, L.; O’Brien, W.; Collman, R.; Essex, M.; Lee, T.-H. Killing of primary CD4+ T cells by non-syncytium-inducing macrophage-tropic human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 1994, 91, 10237–10241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hütter, G.; Nowak, D.; Mossner, M.; Ganepola, S.; Müßig, A.; Allers, K.; Schneider, T.; Hofmann, J.; Kücherer, C.; Blau, O. Long-term control of HIV by CCR5 Delta32/Delta32 stem-cell transplantation. N. Engl. J. Med. 2009, 360, 692–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björndal, A.; Deng, H.; Jansson, M.; Fiore, J.R.; Colognesi, C.; Karlsson, A.; Albert, J.; Scarlatti, G.; Littman, D.R.; Fenyö, E.M. Coreceptor usage of primary human immunodeficiency virus type 1 isolates varies according to biological phenotype. J. Virol. 1997, 71, 7478–7487. [Google Scholar] [CrossRef] [Green Version]
- Goetz, M.B.; Leduc, R.; Kostman, J.R.; Labriola, A.M.; Lie, Y.; Weidler, J.; Coakley, E.; Bates, M.; Luskin-Hawk, R.J. Relationship between HIV co-receptor tropism and disease progression in persons with untreated chronic HIV infection. J. Acquir. Immune Defic. Syndr. 2009, 50, 259. [Google Scholar] [CrossRef] [Green Version]
- Centlivre, M.; Sala, M.; Wain-Hobson, S.; Berkhout, B. In HIV-1 pathogenesis the die is cast during primary infection. Aids 2007, 21, 1–11. [Google Scholar] [CrossRef]
- Chun, T.-W.; Davey, R.T.; Ostrowski, M.; Shawn Justement, J.; Engel, D.; Mullins, J.I.; Fauci, A.S. Relationship between pre-existing viral reservoirs and the re-emergence of plasma viremia after discontinuation of highly active anti-retroviral therapy. Nat. Med. 2000, 6, 757–761. [Google Scholar] [CrossRef]
- Delobel, P.; Sandres-Sauné, K.; Cazabat, M.; L’Faqihi, F.-E.; Aquilina, C.; Obadia, M.; Pasquier, C.; Marchou, B.; Massip, P.; Izopet, J. Persistence of distinct HIV-1 populations in blood monocytes and naive and memory CD4 T cells during prolonged suppressive HAART. Aids 2005, 19, 1739–1750. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Minkah, N.; Leipzig, J.; Wang, G.; Arens, M.Q.; Tebas, P.; Bushman, F.D. DNA bar coding and pyrosequencing to identify rare HIV drug resistance mutations. Nucleic Acids Res. 2007, 35, e91. [Google Scholar] [CrossRef] [PubMed]
- Mardis, E.R. The impact of next-generation sequencing technology on genetics. Trends Genet. 2008, 24, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinones-Mateu, M.; Arts, E.J. Virus fitness: Concept, quantification, and application to HIV population dynamics. In Quasispecies: Concept and Implications for Virology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 83–140. [Google Scholar]
- McAllister, J.; Casino, C.; Davidson, F.; Power, J.; Lawlor, E.; Yap, P.L.; Simmonds, P.; Smith, D.B. Long-term evolution of the hypervariable region of hepatitis C virus in a common-source-infected cohort. J. Virol. 1998, 72, 4893–4905. [Google Scholar] [CrossRef] [Green Version]
- Wainberg, M.A.; Zaharatos, G.J.; Brenner, B.G. Development of antiretroviral drug resistance. N. Engl. J. Med. 2011, 365, 637–646. [Google Scholar] [CrossRef]
- Domingo, E. RNA virus evolution and the control of viral disease. In Progress in Drug Research; Birkhäuser: Basel, Switzerland, 1989; pp. 93–133. [Google Scholar]
- Ribeiro, R.M.; Bonhoeffer, S. Production of resistant HIV mutants during antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2000, 97, 7681–7686. [Google Scholar] [CrossRef] [Green Version]
- Havlir, D.V.; Eastman, S.; Gamst, A.; Richman, D.D. Nevirapine-resistant human immunodeficiency virus: Kinetics of replication and estimated prevalence in untreated patients. J. Virol. 1996, 70, 7894–7899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agudo, R.; Ferrer-Orta, C.; Arias, A.; De La Higuera, I.; Perales, C.; Perez-Luque, R.; Verdaguer, N.; Domingo, E. A multi-step process of viral adaptation to a mutagenic nucleoside analogue by modulation of transition types leads to extinction-escape. PLoS Pathog. 2010, 6, e1001072. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Peng, F.; Wang, R.; Guan, K.; Jiang, T.; Xu, G.; Sun, J.; Chang, C. The deadly coronaviruses: The 2003 SARS pandemic and the 2020 novel coronavirus epidemic in China. J. Autoimmun. 2020, 109, 102434. [Google Scholar] [CrossRef]
- Algaissi, A.A.; Alharbi, N.K.; Hassanain, M.; Hashem, A.M. Preparedness and response to COVID-19 in Saudi Arabia: Building on MERS experience. J. Infect. Public Health 2020, 13, 834–838. [Google Scholar] [CrossRef]
- Karamitros, T.; Papadopoulou, G.; Bousali, M.; Mexias, A.; Tsiodras, S.; Mentis, A. SARS-CoV-2 exhibits intra-host genomic plasticity and low-frequency polymorphic quasispecies. J. Clin. Virol. 2020, 131, 104585. [Google Scholar] [CrossRef]
- Sharma, A.; Tiwari, S.; Deb, M.K.; Marty, J.L. Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2): A global pandemic and treatment strategies. Int. J. Antimicrob. Agents 2020, 56, 106054. [Google Scholar] [CrossRef] [PubMed]
- Nyayanit, D.A.; Yadav, P.D.; Kharde, R.; Shete-Aich, A. Quasispecies analysis of the SARS-CoV-2 from representative clinical samples: A preliminary analysis. Indian J. Med. Res. 2020, 152, 105. [Google Scholar]
- Petersen, E.; Koopmans, M.; Go, U.; Hamer, D.H.; Petrosillo, N.; Castelli, F.; Storgaard, M.; Al Khalili, S.; Simonsen, L. Comparing SARS-CoV-2 with SARS-CoV and influenza pandemics. Lancet Infect. Dis. 2020, 20, e238–e244. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Lian, X.; Su, X.; Wu, W.; Marraro, G.A.; Zeng, Y. From SARS and MERS to COVID-19: A brief summary and comparison of severe acute respiratory infections caused by three highly pathogenic human coronaviruses. Respir. Res. 2020, 21, 224. [Google Scholar] [CrossRef] [PubMed]
- Terada, Y.; Kawachi, K.; Matsuura, Y.; Kamitani, W. MERS coronavirus nsp1 participates in an efficient propagation through a specific interaction with viral RNA. Virology 2017, 511, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Rowe, L.A.; Frace, M.; Stevens, J.; Abedi, G.R.; Elnile, O.; Banassir, T.; Al-Masri, M.; Watson, J.T.; Assiri, A.J. Spike gene deletion quasispecies in serum of patient with acute MERS-CoV infection. J. Med. Virol. 2017, 89, 542–545. [Google Scholar] [CrossRef]
- Park, D.; Huh, H.J.; Kim, Y.J.; Son, D.-S.; Jeon, H.-J.; Im, E.-H.; Kim, J.-W.; Lee, N.Y.; Kang, E.-S.; Kang, C.I. Analysis of intrapatient heterogeneity uncovers the microevolution of Middle East respiratory syndrome coronavirus. Mol. Case Stud. 2016, 2, a001214. [Google Scholar] [CrossRef] [Green Version]
- Scobey, T.; Yount, B.L.; Sims, A.C.; Donaldson, E.F.; Agnihothram, S.S.; Menachery, V.D.; Graham, R.L.; Swanstrom, J.; Bove, P.F.; Kim, J.D. Reverse genetics with a full-length infectious cDNA of the Middle East respiratory syndrome coronavirus. Proc. Natl. Acad. Sci. USA 2013, 110, 16157–16162. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Zhang, Z.; Wang, F.-S. SARS-associated coronavirus quasispecies in individual patients. N. Engl. J. Med. 2004, 350, 1366–1367. [Google Scholar] [CrossRef]
- Rueca, M.; Bartolini, B.; Gruber, C.E.M.; Piralla, A.; Baldanti, F.; Giombini, E.; Messina, F.; Marchioni, L.; Ippolito, G.; Di Caro, A. Compartmentalized replication of SARS-Cov-2 in upper vs. lower respiratory tract assessed by whole genome quasispecies analysis. Microorganisms 2020, 8, 1302. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, A.J. Hepatitis Viruses; University of Texas Medical Branch: Galveston, TX, USA, 1996. [Google Scholar]
- Tsukiyama-Kohara, K.; Kohara, M. Hepatitis C virus: Viral quasispecies and genotypes. Int. J. Mol. Sci. 2017, 19, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevaliez, S.; Pawlotsky, J.-M. HCV Genome and Life Cycle; Horizon Bioscience: Norfolk, UK, 2006. [Google Scholar]
- Zawilińska, B.; Kosz-Vnenchak, M. General introduction into the Ebola virus biology and disease. Folia Med. Cracov. 2014, 54, 57–65. [Google Scholar] [PubMed]
- Giovanetti, M.; Grifoni, A.; Lo Presti, A.; Cella, E.; Montesano, C.; Zehender, G.; Colizzi, V.; Amicosante, M.; Ciccozzi, M.J. Amino acid mutations in Ebola virus glycoprotein of the 2014 epidemic. J. Med. Virol. 2015, 87, 893–898. [Google Scholar] [CrossRef] [PubMed]
- Diehl, W.E.; Lin, A.E.; Grubaugh, N.D.; Carvalho, L.M.; Kim, K.; Kyawe, P.P.; McCauley, S.M.; Donnard, E.; Kucukural, A.; McDonel, P. Ebola virus glycoprotein with increased infectivity dominated the 2013–2016 epidemic. Cell 2016, 167, 1088–1098.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Crone, L.; Dietzel, E.; Paijo, J.; González-Hernández, M.; Nehlmeier, I.; Kalinke, U.; Becker, S.; Pöhlmann, S.J. A polymorphism within the internal fusion loop of the Ebola virus glycoprotein modulates host cell entry. J. Virol. 2017, 91, e00177-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.K.; Lim, S.-Y.; Lee, S.M.; Cunningham, J.M. Biochemical basis for increased activity of Ebola glycoprotein in the 2013–16 epidemic. Cell Host Microbe 2017, 21, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Fitzsimmons, W.J.; Woods, R.J.; McCrone, J.T.; Woodman, A.; Arnold, J.J.; Yennawar, M.; Evans, R.; Cameron, C.E.; Lauring, A.S. A speed–fidelity trade-off determines the mutation rate and virulence of an RNA virus. PLoS Biol. 2018, 16, e2006459. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Dolan, P.T.; Goldstein, E.F.; Li, M.; Farkov, M.; Brodsky, L.; Andino, R. Poliovirus intrahost evolution is required to overcome tissue-specific innate immune responses. Nat. Commun. 2017, 8, 375. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C. Arrival of chikungunya virus in the new world: Prospects for spread and impact on public health. PLoS Negl. Trop. Dis. 2014, 8, e2921. [Google Scholar] [CrossRef] [Green Version]
- Leparc-Goffart, I.; Nougairede, A.; Cassadou, S.; Prat, C.; De Lamballerie, X. Chikungunya in the Americas. Lancet 2014, 383, 514. [Google Scholar] [CrossRef]
- Riemersma, K.K.; Steiner, C.; Singapuri, A.; Coffey, L.L. Chikungunya virus fidelity variants exhibit differential attenuation and population diversity in cell culture and adult mice. PLoS Negl. Trop. Dis. 2019, 93, e01606–e01618. [Google Scholar] [CrossRef] [Green Version]
- Jaimipak, T.; Yoksan, S.; Ubol, S.; Pulmanausahakul, R. Small plaque size variant of Chikungunya primary isolate showed reduced virulence in mice. Asian Pac. J. Allergy Immunol. 2018, 36, 201–205. [Google Scholar]
- Woo, H.-J.; Reifman, J. A quantitative quasispecies theory-based model of virus escape mutation under immune selection. Proc. Natl. Acad. Sci. USA 2012, 109, 12980–12985. [Google Scholar] [CrossRef]
- Figlerowicz, M.; Alejska, M.; Kurzyńska-Kokorniak, A.; Figlerowicz, M. Genetic variability: The key problem in the prevention and therapy of RNA-based virus infections. Med. Res. Rev. 2003, 23, 488–518. [Google Scholar] [CrossRef] [PubMed]
- Gleed, M.L.; Ioannidis, H.; Kolocouris, A.; Busath, D.D. Resistance-mutation (N31) effects on drug orientation and channel hydration in amantadine-bound influenza A M2. J. Phys. Chem. B 2015, 119, 11548–11559. [Google Scholar] [CrossRef] [PubMed]
- McClellan, K.; Perry, C.M. Oseltamivir. Drugs 2001, 61, 263–283. [Google Scholar] [CrossRef] [PubMed]
- Castanotto, D.; Rossi, J. The promises and pitfalls of RNA-interference-based therapeutics. Nature 2009, 457, 426–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presloid, J.B.; Novella, I.S. RNA viruses and RNAi: Quasispecies implications for viral escape. Viruses 2015, 7, 3226–3240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Huang, W.; Wang, Y.; Guan, W.; Li, R.; Yang, Z.; Zhong, N. Gene silencing of β-galactosamide α-2, 6-sialyltransferase 1 inhibits human influenza virus infection of airway epithelial cells. BMC Microbiol. 2014, 14, 78. [Google Scholar] [CrossRef] [Green Version]
- Gitlin, L.; Stone, J.K.; Andino, R.J. Poliovirus escape from RNA interference: Short interfering RNA-target recognition and implications for therapeutic approaches. J. Virol. 2005, 79, 1027–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Xue, Y.; Wang, B.; Du, J.; Jin, Q. Broad-spectrum antiviral activity of RNA interference against four genotypes of Japanese encephalitis virus based on single microRNA polycistrons. PLoS ONE 2011, 6, e26304. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.A.; Richardson, C.D. Hepatitis C virus replicons escape RNA interference induced by a short interfering RNA directed against the NS5b coding region. J. Virol. 2005, 79, 7050–7058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullins, J.I.; Heath, L.; Hughes, J.P.; Kicha, J.; Styrchak, S.; Wong, K.G.; Rao, U.; Hansen, A.; Harris, K.S.; Laurent, J.-P. Mutation of HIV-1 genomes in a clinical population treated with the mutagenic nucleoside KP1461. PLoS ONE 2011, 6, e15135. [Google Scholar] [CrossRef]
- Crowder, S.; Kirkegaard, K. Trans-dominant inhibition of RNA viral replication can slow growth of drug-resistant viruses. Nat. Genet. 2005, 37, 701–709. [Google Scholar] [CrossRef]
- Spagnolo, J.F.; Rossignol, E.; Bullitt, E.; Kirkegaard, K. Enzymatic and nonenzymatic functions of viral RNA-dependent RNA polymerases within oligomeric arrays. RNA 2010, 16, 382–393. [Google Scholar] [CrossRef] [Green Version]
- Perales, C.; Agudo, R.; Tejero, H.; Manrubia, S.C.; Domingo, E. Potential benefits of sequential inhibitor-mutagen treatments of RNA virus infections. PLoS Pathog. 2009, 5, e1000658. [Google Scholar] [CrossRef] [Green Version]
- Grande-Pérez, A.; Lázaro, E.; Lowenstein, P.; Domingo, E.; Manrubia, S.C. Suppression of viral infectivity through lethal defection. Proc. Natl. Acad. Sci. USA 2005, 102, 4448–4452. [Google Scholar] [CrossRef] [Green Version]
- Ojosnegros, S.; Agudo, R.; Sierra, M.; Briones, C.; Sierra, S.; González-López, C.; Domingo, E.; Cristina, J. Topology of evolving, mutagenized viral populations: Quasispecies expansion, compression, and operation of negative selection. BMC Evol. Biol. 2008, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Perales, C.; Iranzo, J.; Manrubia, S.C.; Domingo, E. The impact of quasispecies dynamics on the use of therapeutics. Trends Microbiol. 2012, 20, 595–603. [Google Scholar] [CrossRef]
- Perales, C.; Agudo, R.; Domingo, E. Counteracting quasispecies adaptability: Extinction of a ribavirin-resistant virus mutant by an alternative mutagenic treatment. PLoS ONE 2009, 4, e5554. [Google Scholar] [CrossRef] [PubMed]

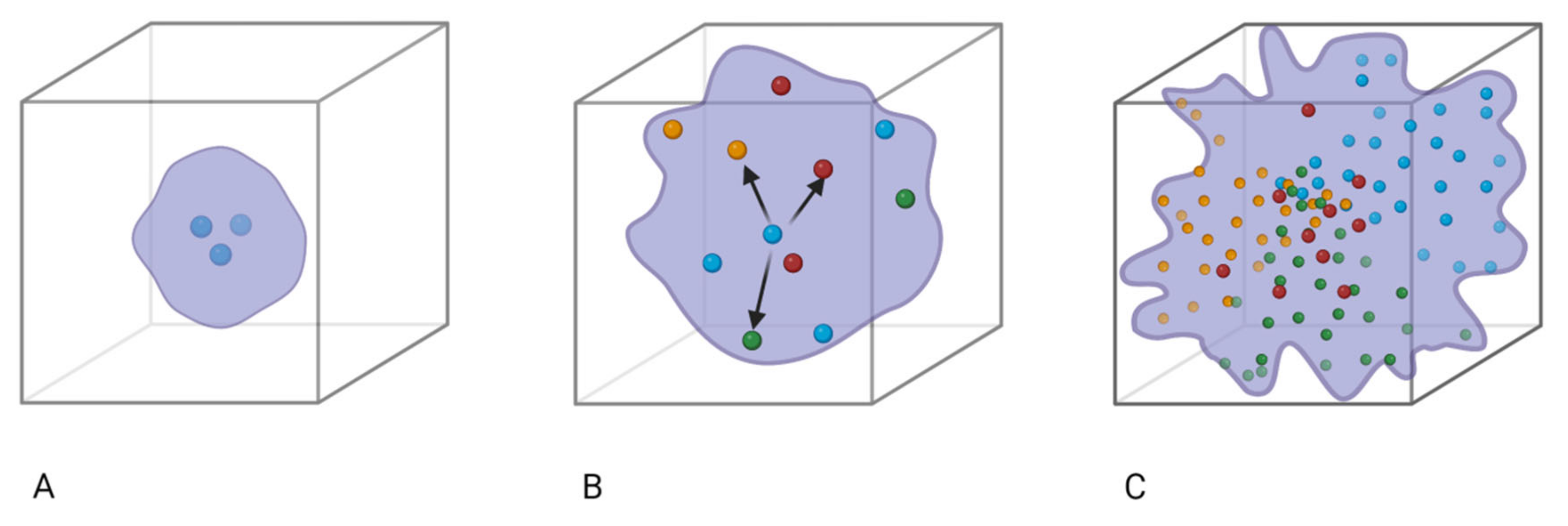
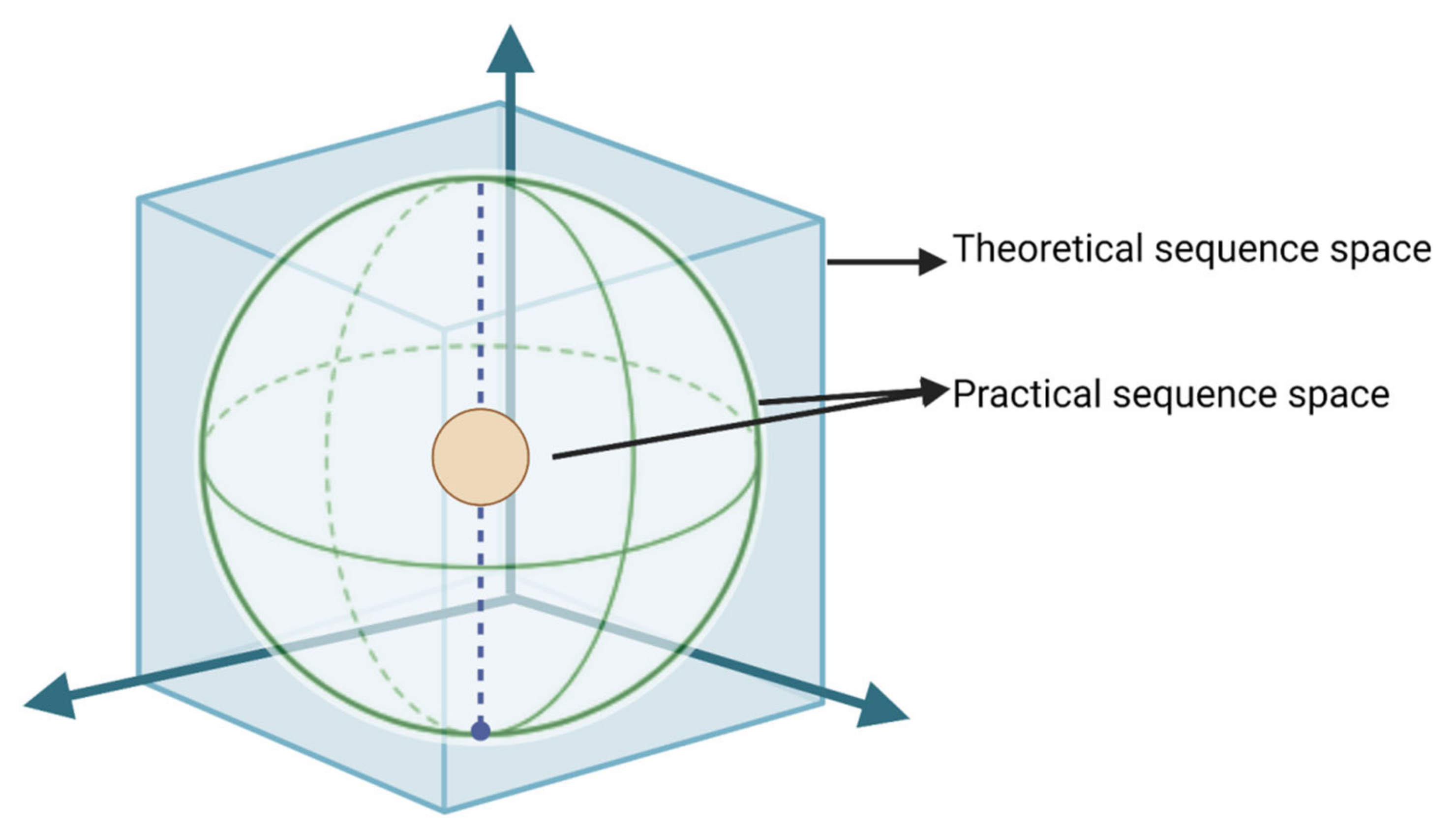


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Family | Mutagenic Base and Nucleotide Analog |
|---|---|---|
| Polio virus | Picornaviridae | Ribavirin, 5-nitrocytidine |
| Dengue virus | Flavivirus | 5-Fluorouracil, ribavirin |
| Zika virus | Flavivirus | Ribavirin, favipiravir |
| Hepatitis C virus | Flaviviridae | Ribavirin, favipiravir |
| Hepatitis E virus | Hepeviridae | Ribavirin |
| SARS-CoV-2 | Coronaviridae | Favipiravir |
| Tobacco mosaic virus | Virgaviridae | 5-fluorouracil |
| Influenza A virus | Orthomyxoviridae | 5-fluorouracil, Ribavirin 5-azacytidine |
| Vesicular stomatitis virus | Rhabdoviridae | 5-fluorouracil |
| Hantaan virus | Hantaviridae | Ribavirin |
| Rift valley fever virus | Phenuiviridae | Favipiravir |
| Ebola virus | Filoviridae | 5-fluorouracil |
| Marburg virus | Filovirade | Favipiravir |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, K.; Mehta, D.; Dumka, S.; Chauhan, A.S.; Kumar, S. Quasispecies Nature of RNA Viruses: Lessons from the Past. Vaccines 2023, 11, 308. https://doi.org/10.3390/vaccines11020308
Singh K, Mehta D, Dumka S, Chauhan AS, Kumar S. Quasispecies Nature of RNA Viruses: Lessons from the Past. Vaccines. 2023; 11(2):308. https://doi.org/10.3390/vaccines11020308
Chicago/Turabian StyleSingh, Kiran, Deepa Mehta, Shaurya Dumka, Aditya Singh Chauhan, and Sachin Kumar. 2023. "Quasispecies Nature of RNA Viruses: Lessons from the Past" Vaccines 11, no. 2: 308. https://doi.org/10.3390/vaccines11020308
APA StyleSingh, K., Mehta, D., Dumka, S., Chauhan, A. S., & Kumar, S. (2023). Quasispecies Nature of RNA Viruses: Lessons from the Past. Vaccines, 11(2), 308. https://doi.org/10.3390/vaccines11020308