
No Difference in the Prevalence of HIV-1 gag Cytotoxic T-Lymphocyte-Associated Escape Mutations in Viral Sequences from Early and Late Parts of the HIV-1 Subtype C Pandemic in Botswana

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of the Study Samples and Determination of HIV Recent Infection
2.2. Ethical Statement and Consent Process
2.3. HIV-gag Genotyping (Extraction, Amplification and Sequencing)
2.3.1. HIV-1 RNA Extraction and Amplification Used to Generate N1 Sequences
2.3.2. Sequencing of gag
2.4. Hypermutation Screening in BCPP Participants
2.5. Chromatograms and Data Sorting
2.6. Determining Cytotoxic T Lymphocyte Escape Mutations and Statistical Analysis
2.7. Phylogenetic Tree

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Escape Mutation | Significance of Mutations | Reference |
|---|---|---|---|
| P17 | K18R | Elicit lytic response | [21] |
| R20K | [22] | ||
| K26R | Reverting mutation | [23] | |
| H28Q | [24] | ||
| K28R | [24] | ||
| M30R | [25] | ||
| Y79F | [26] | ||
| Y79H | Diminished HLA binding | [27] | |
| A83T | [28] | ||
| E93K | No CTL response detected | [29] | |
| P24 | V128G | Diminish CTL response | [30] |
| A146P | Affect peptide processing | [31] | |
| I147L | Impair replication capacity | [7] | |
| I147M | Escape from epitope | [32] | |
| A163G | Affect presentation by HLA-I | [33] | |
| A165N | Compensatory mutation to A163G | [34] | |
| E173E | Compensatory mutation | [55] | |
| Q177D | [26] | ||
| Q182T | Not recognized by CTL at all | [36] | |
| Q182E | Reduces viral replication capacity | [37] | |
| Q182G | [37,38] | ||
| Q182S | [32,39] | ||
| T186S | [32,39] | ||
| V215L | Loss of viral fitness | [40] | |
| H219Q | Compensate deleterious effect caused by T242N | [41] | |
| I223V | [42] | ||
| M228L | [42] | ||
| T242N | Affect presentation by HLA-I | ||
| I247V | [43] | ||
| G248A | In combination with T242N affect presentation by HLA-l | ||
| G248Q | [44] | ||
| G248T | Reduces viral load on HLA-57/58 positive | [45] | |
| G248D | [45] | ||
| A248G | [46] | ||
| D260E | [47] | ||
| R264K | [44] | ||
| L268M | Affect recognition by TCR | [48] | |
| K302R | [39] | ||
| T310S | [49] | ||
| D312E | An escape to HLA-B 5801 | [50] | |
| T332N | [51] | ||
| G357S | [52] |
3. Results
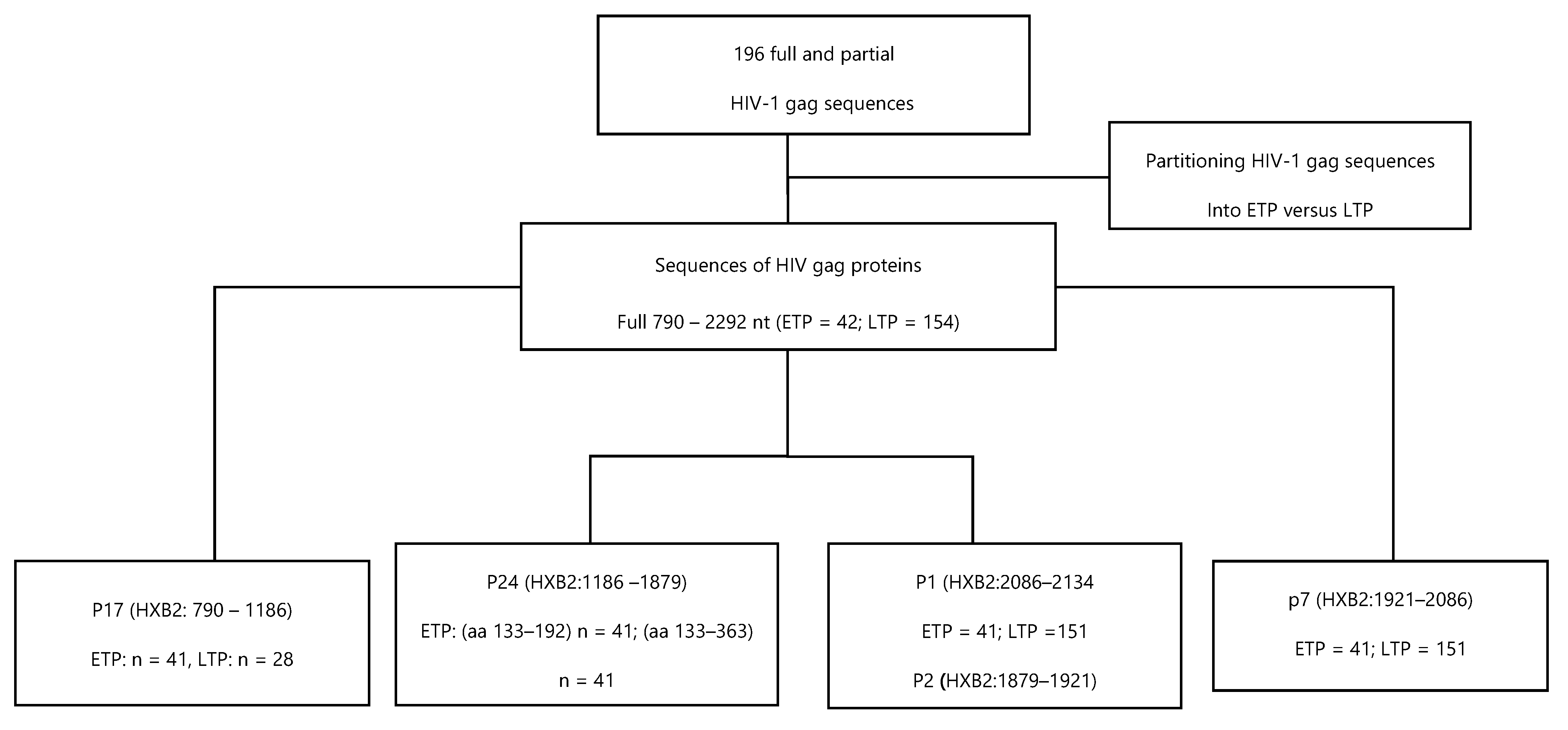
3.1. Characteristics of ETP and LTP HIV-1 gag Isolates
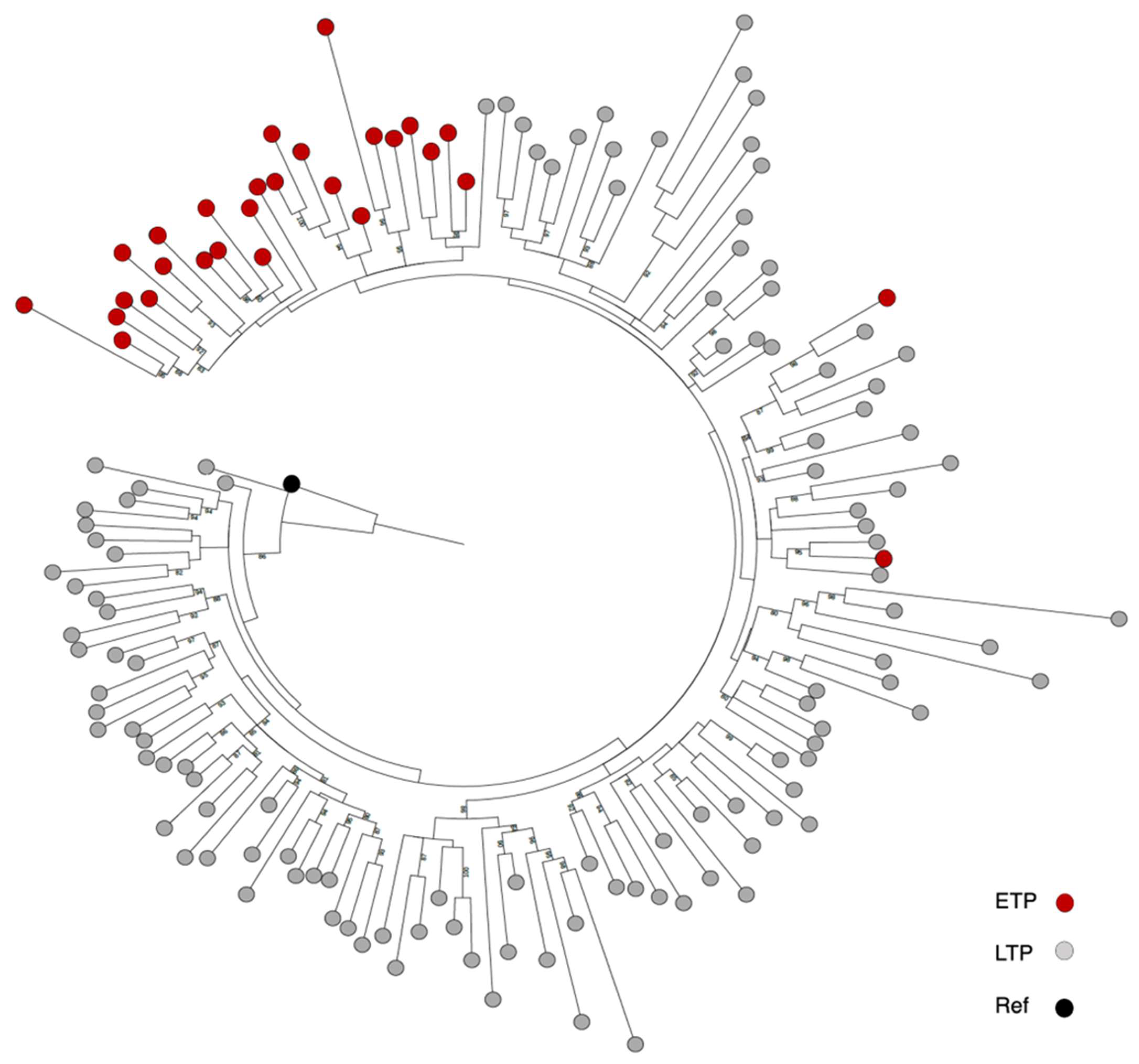
3.2. Comparing the Diversity of Historical and Recent HIV Isolates
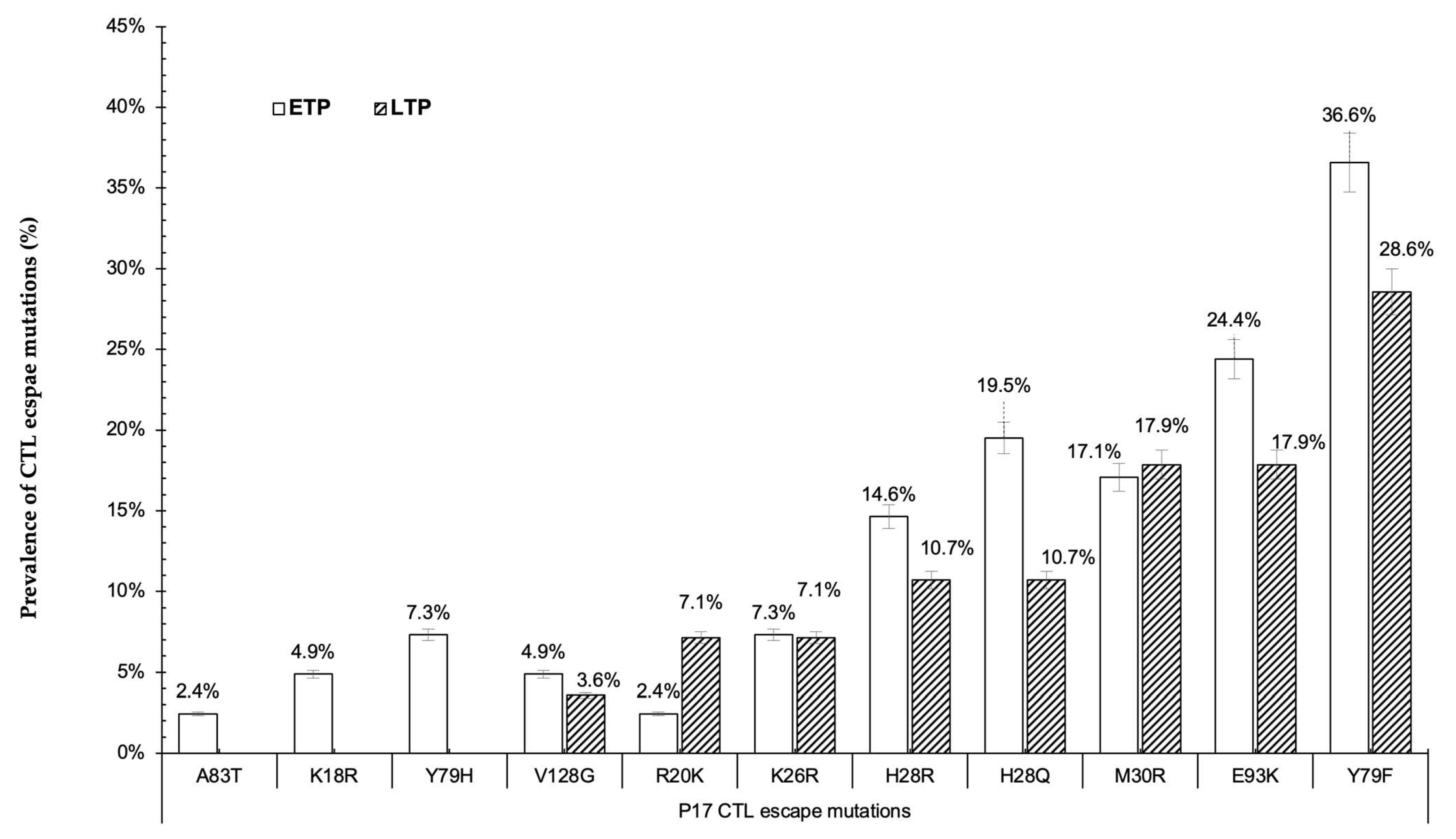
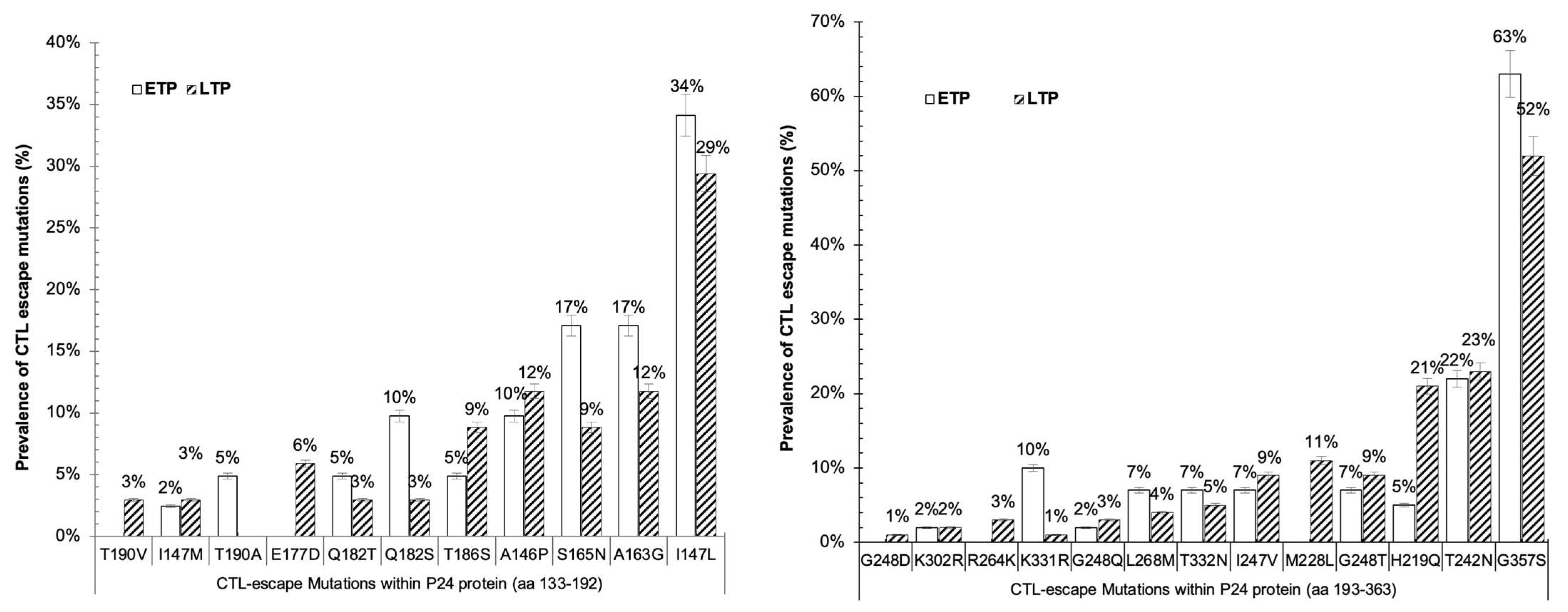
3.3. CTL Escape Mutations in gag Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AIDS. In Danger: UNAIDS Global AIDS Update 2022; United Nations Publications: New York, NY, USA, 2022. [Google Scholar]
- Collins, D.R.; Gaiha, G.D.; Walker, B.D. CD8(+) T cells in HIV control, cure and prevention. Nat. Rev. Immunol. 2020, 20, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Croome, N.; Ahluwalia, M.; Hughes, L.D.; Abas, M. Patient-reported barriers and facilitators to antiretroviral adherence in sub-Saharan Africa. Aids 2017, 31, 995–1007. [Google Scholar] [CrossRef]
- Wallis, C.L.; Godfrey, C.; Fitzgibbon, J.E.; Mellors, J.W. Key Factors Influencing the Emergence of Human Immunodeficiency Virus Drug Resistance in Low- and Middle-Income Countries. J. Infect. Dis. 2017, 216, S851–S856. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Hosoya, N.; Brumme, Z.L.; Brockman, M.A.; Kikuchi, T.; Koga, M.; Nakamura, H.; Koibuchi, T.; Fujii, T.; Carlson, J.M.; et al. Significant reductions in Gag-protease-mediated HIV-1 replication capacity during the course of the epidemic in Japan. J. Virol. 2013, 87, 1465–1476. [Google Scholar] [CrossRef] [PubMed]
- Clavel, F.; Hance, A.J. HIV drug resistance. N. Engl. J. Med. 2004, 350, 1023–1035. [Google Scholar] [CrossRef]
- Miura, T.; Brockman, M.A.; Schneidewind, A.; Lobritz, M.; Pereyra, F.; Rathod, A.; Block, B.L.; Brumme, Z.L.; Brumme, C.J.; Baker, B.; et al. HLA-B57/B*5801 human immunodeficiency virus type 1 elite controllers select for rare gag variants associated with reduced viral replication capacity and strong cytotoxic T-lymphocyte [corrected] recognition. J. Virol. 2009, 83, 2743–2755. [Google Scholar] [CrossRef]
- Kaseke, C.; Tano-Menka, R.; Senjobe, F.; Gaiha, G.D. The Emerging Role for CTL Epitope Specificity in HIV Cure Efforts. J. Infect. Dis. 2021, 223 (Suppl. S1), 32–37. [Google Scholar] [CrossRef]
- Lemey, P.; Rambaut, A.; Pybus, O.G. HIV evolutionary dynamics within and among hosts. AIDS Rev. 2006, 8, 125–140. [Google Scholar]
- Kawashima, Y.; Pfafferott, K.; Frater, J.; Matthews, P.; Payne, R.; Addo, M.; Gatanaga, H.; Fujiwara, M.; Hachiya, A.; Koizumi, H.; et al. Adaptation of HIV-1 to human leukocyte antigen class I. Nature 2009, 458, 641–645. [Google Scholar] [CrossRef]
- Moore, C.B.; John, M.; James, I.R.; Christiansen, F.T.; Witt, C.S.; Mallal, S.A. Evidence of HIV-1 adaptation to HLA-restricted immune responses at a population level. Science 2002, 296, 1439–1443. [Google Scholar] [CrossRef]
- Novitsky, V.; Flores-Villanueva, P.O.; Chigwedere, P.; Gaolekwe, S.; Bussman, H.; Sebetso, G.; Marlink, R.; Yunis, E.J.; Essex, M. Identification of most frequent HLA class I antigen specificities in Botswana: Relevance for HIV vaccine design. Hum. Immunol. 2001, 62, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Ndung’u, T.; Gaseitsiwe, S.; Sepako, E.; Doualla-Bell, F.; Peter, T.; Kim, S.; Thior, I.; Novitsky, V.A.; Essex, M. Major histocompatibility complex class II (HLA-DRB and -DQB) allele frequencies in Botswana: Association with human immunodeficiency virus type 1 infection. Clin. Diagn. Lab. Immunol. 2005, 12, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Schellens, I.M.; Navis, M.; van Deutekom, H.W.; Boeser-Nunnink, B.; Berkhout, B.; Kootstra, N.; Miedema, F.; Kesmir, C.; Schuitemaker, H.; van Baarle, D.; et al. Loss of HIV-1-derived cytotoxic T lymphocyte epitopes restricted by protective HLA-B alleles during the HIV-1 epidemic. AIDS 2011, 25, 1691–1700. [Google Scholar] [CrossRef]
- Moyo, S.; Gaseitsiwe, S.; Mohammed, T.; Pretorius Holme, M.; Wang, R.; Kotokwe, K.P.; Boleo, C.; Mupfumi, L.; Yankinda, E.K.; Chakalisa, U.; et al. Cross-sectional estimates revealed high HIV incidence in Botswana rural communities in the era of successful ART scale-up in 2013–2015. PLoS ONE 2018, 13, e0204840. [Google Scholar] [CrossRef] [PubMed]
- Novitsky, V.; Wang, R.; Margolin, L.; Baca, J.; Moyo, S.; Musonda, R.; Essex, M. Dynamics and timing of in vivo mutations at Gag residue 242 during primary HIV-1 subtype C infection. Virology 2010, 403, 37–46. [Google Scholar] [CrossRef]
- Rowley, C.F.; MacLeod, I.J.; Maruapula, D.; Lekoko, B.; Gaseitsiwe, S.; Mine, M.; Essex, M. Sharp increase in rates of HIV transmitted drug resistance at antenatal clinics in Botswana demonstrates the need for routine surveillance. J. Antimicrob. Chemother 2016, 71, 1361–1366. [Google Scholar] [CrossRef]
- Novitsky, V.; Wang, R.; Margolin, L.; Baca, J.; Kebaabetswe, L.; Rossenkhan, R.; Bonney, C.; Herzig, M.; Nkwe, D.; Moyo, S.; et al. Timing constraints of in vivo gag mutations during primary HIV-1 subtype C infection. PLoS ONE 2009, 4, e7727. [Google Scholar] [CrossRef] [PubMed]
- Gaolathe, T.; Wirth, K.E.; Holme, M.P.; Makhema, J.; Moyo, S.; Chakalisa, U.; Yankinda, E.K.; Lei, Q.; Mmalane, M.; Novitsky, V.; et al. Botswana’s progress toward achieving the 2020 UNAIDS 90-90-90 antiretroviral therapy and virological suppression goals: A population-based survey. Lancet HIV 2016, 3, e221–e230. [Google Scholar] [CrossRef]
- Wilson, C.C.; Brown, R.C.; Korber, B.T.; Wilkes, B.M.; Ruhl, D.J.; Sakamoto, D.; Kunstman, K.; Luzuriaga, K.; Hanson, I.C.; Widmayer, S.M.; et al. Frequent detection of escape from cytotoxic T-lymphocyte recognition in perinatal human immunodeficiency virus (HIV) type 1 transmission: The ariel project for the prevention of transmission of HIV from mother to infant. J. Virol. 1999, 73, 3975–3985. [Google Scholar] [CrossRef]
- Harrer, T.; Harrer, E.; Kalams, S.A.; Barbosa, P.; Trocha, A.; Johnson, R.P.; Elbeik, T.; Feinberg, M.B.; Buchbinder, S.P.; Walker, B.D. Cytotoxic T lymphocytes in asymptomatic long-term nonprogressing HIV-1 infection. Breadth and specificity of the response and relation to in vivo viral quasispecies in a person with prolonged infection and low viral load. J. Immunol. 1996, 156, 2616–2623. [Google Scholar] [CrossRef]
- Vollbrecht, T.; Brackmann, H.; Henrich, N.; Roeling, J.; Seybold, U.; Bogner, J.R.; Goebel, F.D.; Draenert, R. Impact of changes in antigen level on CD38/PD-1 co-expression on HIV-specific CD8 T cells in chronic, untreated HIV-1 infection. J. Med. Virol. 2010, 82, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Chopera, D.R.; Mlotshwa, M.; Woodman, Z.; Mlisana, K.; de Assis Rosa, D.; Martin, D.P.; Abdool Karim, S.; Gray, C.M.; Williamson, C. Virological and immunological factors associated with HIV-1 differential disease progression in HLA-B 58:01-positive individuals. J. Virol. 2011, 85, 7070–7080. [Google Scholar] [CrossRef] [PubMed]
- Kloverpris, H.N.; Stryhn, A.; Harndahl, M.; van der Stok, M.; Payne, R.P.; Matthews, P.C.; Chen, F.; Riddell, L.; Walker, B.D.; Ndung’u, T.; et al. HLA-B*57 Micropolymorphism shapes HLA allele-specific epitope immunogenicity, selection pressure, and HIV immune control. J. Virol. 2012, 86, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Ntale, R.S.; Chopera, D.R.; Ngandu, N.K.; Assis de Rosa, D.; Zembe, L.; Gamieldien, H.; Mlotshwa, M.; Werner, L.; Woodman, Z.; Mlisana, K.; et al. Temporal association of HLA-B*81:01- and HLA-B*39:10-mediated HIV-1 p24 sequence evolution with disease progression. J. Virol. 2012, 86, 12013–12024. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, B.D.; Yang, O.O.; Hultin, L.; Hausner, M.A.; Hultin, P.; Matud, J.; Kunstman, K.; Killian, S.; Altman, J.; Kommander, K.; et al. Epitope escape mutation and decay of human immunodeficiency virus type 1-specific CTL responses. J. Immunol. 2003, 171, 5372–5379. [Google Scholar] [CrossRef] [PubMed]
- Iversen, A.K.; Stewart-Jones, G.; Learn, G.H.; Christie, N.; Sylvester-Hviid, C.; Armitage, A.E.; Kaul, R.; Beattie, T.; Lee, J.K.; Li, Y.; et al. Conflicting selective forces affect T cell receptor contacts in an immunodominant human immunodeficiency virus epitope. Nat. Immunol. 2006, 7, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, A.C.; Iversen, A.K.; Chapman, J.M.; de Oliviera, T.; Spotts, G.; McMichael, A.J.; Davenport, M.P.; Hecht, F.M.; Nixon, D.F. Sequential broadening of CTL responses in early HIV-1 infection is associated with viral escape. PLoS ONE 2007, 2, e225. [Google Scholar] [CrossRef]
- Jones, N.A.; Wei, X.; Flower, D.R.; Wong, M.; Michor, F.; Saag, M.S.; Hahn, B.H.; Nowak, M.A.; Shaw, G.M.; Borrow, P. Determinants of human immunodeficiency virus type 1 escape from the primary CD8+ cytotoxic T lymphocyte response. J. Exp. Med. 2004, 200, 1243–1256. [Google Scholar] [CrossRef]
- Carlson, J.M.; Listgarten, J.; Pfeifer, N.; Tan, V.; Kadie, C.; Walker, B.D.; Ndung’u, T.; Shapiro, R.; Frater, J.; Brumme, Z.L.; et al. Widespread impact of HLA restriction on immune control and escape pathways of HIV-1. J. Virol. 2012, 86, 5230–5243. [Google Scholar] [CrossRef]
- Aidoo, M.; Sawadogo, S.; Bile, E.C.; Yang, C.; Nkengasong, J.N.; McNicholl, J.M. Viral, HLA and T cell elements in cross-reactive immune responses to HIV-1 subtype A, CRF01_AE and CRF02_AG vaccine sequence in Ivorian blood donors. Vaccine 2008, 26, 4830–4839. [Google Scholar] [CrossRef]
- Leslie, A.; Price, D.A.; Mkhize, P.; Bishop, K.; Rathod, A.; Day, C.; Crawford, H.; Honeyborne, I.; Asher, T.E.; Luzzi, G.; et al. Differential selection pressure exerted on HIV by CTL targeting identical epitopes but restricted by distinct HLA alleles from the same HLA supertype. J. Immunol. 2006, 177, 4699–4708. [Google Scholar] [CrossRef] [PubMed]
- Kløverpris, H.N.; Leslie, A.; Goulder, P. Role of HLA Adaptation in HIV Evolution. Front. Immunol. 2015, 6, 665. [Google Scholar] [CrossRef] [PubMed]
- Geldmacher, C.; Metzler, I.S.; Tovanabutra, S.; Asher, T.E.; Gostick, E.; Ambrozak, D.R.; Petrovas, C.; Schuetz, A.; Ngwenyama, N.; Kijak, G.; et al. Minor viral and host genetic polymorphisms can dramatically impact the biologic outcome of an epitope-specific CD8 T-cell response. Blood 2009, 114, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Leitman, E.M.; Thobakgale, C.F.; Adland, E.; Ansari, M.A.; Raghwani, J.; Prendergast, A.J.; Tudor-Williams, G.; Kiepiela, P.; Hemelaar, J.; Brener, J.; et al. Role of HIV-specific CD8(+) T cells in pediatric HIV cure strategies after widespread early viral escape. J. Exp. Med. 2017, 214, 3239–3261. [Google Scholar] [CrossRef]
- Troyer, R.M.; McNevin, J.; Liu, Y.; Zhang, S.C.; Krizan, R.W.; Abraha, A.; Tebit, D.M.; Zhao, H.; Avila, S.; Lobritz, M.A.; et al. Variable fitness impact of HIV-1 escape mutations to cytotoxic T lymphocyte (CTL) response. PLoS Pathog. 2009, 5, e1000365. [Google Scholar] [CrossRef]
- Hoof, I.; Pérez, C.L.; Buggert, M.; Gustafsson, R.K.; Nielsen, M.; Lund, O.; Karlsson, A.C. Interdisciplinary analysis of HIV-specific CD8+ T cell responses against variant epitopes reveals restricted TCR promiscuity. J. Immunol. 2010, 184, 5383–5391. [Google Scholar] [CrossRef]
- Tang, Y.; Huang, S.; Dunkley-Thompson, J.; Steel-Duncan, J.C.; Ryland, E.G.; St John, M.A.; Hazra, R.; Christie, C.D.; Feeney, M.E. Correlates of spontaneous viral control among long-term survivors of perinatal HIV-1 infection expressing human leukocyte antigen-B57. AIDS 2010, 24, 1425–1435. [Google Scholar] [CrossRef]
- Kløverpris, H.N.; Stryhn, A.; Harndahl, M.; Carlson, J.M.; Leslie, A.J.; Chen, F.; Riddell, L.; Mulenga, J.; Walker, B.D.; Ndung’u, T.; et al. HLA-A*68:02-restricted Gag-specific cytotoxic T lymphocyte responses can drive selection pressure on HIV but are subdominant and ineffective. AIDS 2013, 27, 1717. [Google Scholar] [CrossRef]
- Turnbull, E.L.; Lopes, A.R.; Jones, N.A.; Cornforth, D.; Newton, P.; Aldam, D.; Pellegrino, P.; Turner, J.; Williams, I.; Wilson, C.M.; et al. HIV-1 epitope-specific CD8+ T cell responses strongly associated with delayed disease progression cross-recognize epitope variants efficiently. J. Immunol. 2006, 176, 6130–6146. [Google Scholar] [CrossRef]
- Martinez-Picado, J.; Prado, J.G.; Fry, E.E.; Pfafferott, K.; Leslie, A.; Chetty, S.; Thobakgale, C.; Honeyborne, I.; Crawford, H.; Matthews, P.; et al. Fitness cost of escape mutations in p24 Gag in association with control of human immunodeficiency virus type 1. J. Virol. 2006, 80, 3617–3623. [Google Scholar] [CrossRef]
- Tenzer, S.; Crawford, H.; Pymm, P.; Gifford, R.; Sreenu, V.B.; Weimershaus, M.; de Oliveira, T.; Burgevin, A.; Gerstoft, J.; Akkad, N.; et al. HIV-1 adaptation to antigen processing results in population-level immune evasion and affects subtype diversification. Cell Rep. 2014, 7, 448–463. [Google Scholar] [CrossRef] [PubMed]
- Matthews, P.C.; Koyanagi, M.; Kløverpris, H.N.; Harndahl, M.; Stryhn, A.; Akahoshi, T.; Gatanaga, H.; Oka, S.; Juarez Molina, C.; Valenzuela Ponce, H.; et al. Differential clade-specific HLA-B*3501 association with HIV-1 disease outcome is linked to immunogenicity of a single Gag epitope. J. Virol. 2012, 86, 12643–12654. [Google Scholar] [CrossRef] [PubMed]
- Frater, A.J.; Brown, H.; Oxenius, A.; Günthard, H.F.; Hirschel, B.; Robinson, N.; Leslie, A.J.; Payne, R.; Crawford, H.; Prendergast, A.; et al. Effective T-cell responses select human immunodeficiency virus mutants and slow disease progression. J. Virol. 2007, 81, 6742–6751. [Google Scholar] [CrossRef] [PubMed]
- Dorrell, L.; Willcox, B.E.; Jones, E.Y.; Gillespie, G.; Njai, H.; Sabally, S.; Jaye, A.; DeGleria, K.; Rostron, T.; Lepin, E.; et al. Cytotoxic T lymphocytes recognize structurally diverse, clade-specific and cross-reactive peptides in human immunodeficiency virus type-1 gag through HLA-B53. Eur. J. Immunol. 2001, 31, 1747–1756. [Google Scholar] [CrossRef]
- Geels, M.J.; Dubey, S.A.; Anderson, K.; Baan, E.; Bakker, M.; Pollakis, G.; Paxton, W.A.; Shiver, J.W.; Goudsmit, J. Broad cross-clade T-cell responses to gag in individuals infected with human immunodeficiency virus type 1 non-B clades (A to G): Importance of HLA anchor residue conservation. J. Virol. 2005, 79, 11247–11258. [Google Scholar] [CrossRef]
- Yokomaku, Y.; Miura, H.; Tomiyama, H.; Kawana-Tachikawa, A.; Takiguchi, M.; Kojima, A.; Nagai, Y.; Iwamoto, A.; Matsuda, Z.; Ariyoshi, K. Impaired Processing and Presentation of Cytotoxic-T-Lymphocyte (CTL) Epitopes Are Major Escape Mechanisms from CTL Immune Pressure in Human Immunodeficiency Virus Type 1 Infection. J. Virol. 2004, 78, 1324–1332. [Google Scholar] [CrossRef]
- Kulkarni, V.; Rosati, M.; Valentin, A.; Ganneru, B.; Singh, A.K.; Yan, J.; Rolland, M.; Alicea, C.; Beach, R.K.; Zhang, G.M.; et al. HIV-1 p24(gag) derived conserved element DNA vaccine increases the breadth of immune response in mice. PLoS ONE 2013, 8, e60245. [Google Scholar] [CrossRef]
- Posada, D. ModelTest Server: A web-based tool for the statistical selection of models of nucleotide substitution online. Nucleic Acids Res. 2006, 34, W700–W703. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Payne, R.P.; Branch, S.; Kløverpris, H.; Matthews, P.C.; Koofhethile, C.K.; Strong, T.; Adland, E.; Leitman, E.; Frater, J.; Ndung’u, T.; et al. Differential escape patterns within the dominant HLA-B*57:03-restricted HIV Gag epitope reflect distinct clade-specific functional constraints. J. Virol. 2014, 88, 4668–4678. [Google Scholar] [CrossRef]
- Navis, M.; Matas, D.E.; Rachinger, A.; Koning, F.A.; van Swieten, P.; Kootstra, N.A.; Schuitemaker, H. Molecular evolution of human immunodeficiency virus type 1 upon transmission between human leukocyte antigen disparate donor-recipient pairs. PLoS ONE 2008, 3, e2422. [Google Scholar] [CrossRef] [PubMed]
- Kiepiela, P.; Leslie, A.J.; Honeyborne, I.; Ramduth, D.; Thobakgale, C.; Chetty, S.; Rathnavalu, P.; Moore, C.; Pfafferott, K.J.; Hilton, L.; et al. Dominant influence of HLA-B in mediating the potential co-evolution of HIV and HLA. Nature 2004, 432, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Leslie, A.J.; Pfafferott, K.J.; Chetty, P.; Draenert, R.; Addo, M.M.; Feeney, M.; Tang, Y.; Holmes, E.C.; Allen, T.; Prado, J.G.; et al. HIV evolution: CTL escape mutation and reversion after transmission. Nat. Med. 2004, 10, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Berger, C.T.; Carlson, J.M.; Brumme, C.J.; Hartman, K.L.; Brumme, Z.L.; Henry, L.M.; Rosato, P.C.; Piechocka-Trocha, A.; Brockman, M.A.; Harrigan, P.R.; et al. Viral adaptation to immune selection pressure by HLA class I-restricted CTL responses targeting epitopes in HIV frameshift sequences. J. Exp. Med. 2010, 207, 61–75. [Google Scholar] [CrossRef]
- Crawford, H.; Lumm, W.; Leslie, A.; Schaefer, M.; Boeras, D.; Prado, J.G.; Tang, J.; Farmer, P.; Ndung’u, T.; Lakhi, S.; et al. Evolution of HLA-B*5703 HIV-1 escape mutations in HLA-B*5703-positive individuals and their transmission recipients. J. Exp. Med. 2009, 206, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.; Muenchhoff, M.; Mann, J.; Roberts, H.E.; Matthews, P.; Adland, E.; Hempenstall, A.; Huang, K.-H.; Brockman, M.; Brumme, Z.; et al. Impact of HLA-driven HIV adaptation on virulence in populations of high HIV seroprevalence. Proc. Natl. Acad. Sci. USA 2014, 111, E5393–E5400. [Google Scholar] [CrossRef]
- Castro-Nallar, E.; Perez-Losada, M.; Burton, G.F.; Crandall, K.A. The evolution of HIV: Inferences using phylogenetics. Mol. Phylogenet. Evol. 2012, 62, 777–792. [Google Scholar] [CrossRef]
- Leitner, T.; Romero-Severson, E. Phylogenetic patterns recover known HIV epidemiological relationships and reveal common transmission of multiple variants. Nat. Microbiol. 2018, 3, 983–988. [Google Scholar] [CrossRef]
- Arien, K.K.; Troyer, R.M.; Gali, Y.; Colebunders, R.L.; Arts, E.J.; Vanham, G. Replicative fitness of historical and recent HIV-1 isolates suggests HIV-1 attenuation over time. AIDS 2005, 19, 1555–1564. [Google Scholar] [CrossRef]
- Song, H.; Pavlicek, J.W.; Cai, F.; Bhattacharya, T.; Li, H.; Iyer, S.S.; Bar, K.J.; Decker, J.M.; Goonetilleke, N.; Liu, M.K.; et al. Impact of immune escape mutations on HIV-1 fitness in the context of the cognate transmitted/founder genome. Retrovirology 2012, 9, 89. [Google Scholar] [CrossRef]
- Yang, Y.; Ganusov, V.V. Kinetics of HIV-Specific CTL Responses Plays a Minimal Role in Determining HIV Escape Dynamics. Front. Immunol. 2018, 9, 140. [Google Scholar] [CrossRef] [PubMed]
- McMichael, A.J.; Phillips, R.E. Escape of human immunodeficiency virus from immune control. Annu. Rev. Immunol. 1997, 15, 271–296. [Google Scholar] [CrossRef] [PubMed]
- Deng, K.; Pertea, M.; Rongvaux, A.; Wang, L.; Durand, C.M.; Ghiaur, G.; Lai, J.; McHugh, H.L.; Hao, H.; Zhang, H.; et al. Broad CTL response is required to clear latent HIV-1 due to dominance of escape mutations. Nature 2015, 517, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Ganusov, V.V.; Barber, D.L.; De Boer, R.J. Killing of targets by CD8 T cells in the mouse spleen follows the law of mass action. PLoS ONE 2011, 6, e15959. [Google Scholar] [CrossRef] [PubMed]
- Ganusov, V.V.; Goonetilleke, N.; Liu, M.K.; Ferrari, G.; Shaw, G.M.; McMichael, A.J.; Borrow, P.; Korber, B.T.; Perelson, A.S. Fitness costs and diversity of the cytotoxic T lymphocyte (CTL) response determine the rate of CTL escape during acute and chronic phases of HIV infection. J. Virol. 2011, 85, 10518–10528. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, T.C.; Dodds, E.J.; Yant, L.J.; Vojnov, L.; Rudersdorf, R.; Cullen, C.; Evans, D.T.; Desrosiers, R.C.; Mothe, B.R.; Sidney, J.; et al. Reversion of CTL escape-variant immunodeficiency viruses in vivo. Nat. Med. 2004, 10, 275–281. [Google Scholar] [CrossRef] [PubMed]




| ETP (N = 42) | LTP (N = 154) | ||
|---|---|---|---|
| Time of sample collection | 2004–2008 | 2012–2018 | |
| Sample used to isolate HIV-1 isolates | Plasma | Buffy coat + plasma | |
| HIV-1 subtype | C (42) | C (154) | |
| Distance matrices | 0.096 | 0.106 | |
| Mean viral load (VL) (copies/mL) (p-value < 0.015) | 31,400 (IQR: 3356–209,500) | 11,152 (IQR: 2475–33,349) | |
| Mean time to seroconversion (days) | Duration of recent infection of approximately 130 days | Duration of recent infection of approximately 130 days | |
| Gender distribution | 9 Males, 33 Females | 28 Males, 126 Females | |
| Age (median; Q1, Q3) years | 27 (25–32) | 26 (22–32) | |
| Method used to measure recency | Timing patients who sero-convert based on Fiebig stage II; IV or V | HIV-1 LAg-Avidity serological assay | |
| Fully covered gag proteins (n) | P17 | 41 | 28 |
| P24 | 41 | 34 | |
| P1P2 | 41 | 151 | |
| P6P7 | 41 | 151 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mokaleng, B.; Choga, W.T.; Bareng, O.T.; Maruapula, D.; Ditshwanelo, D.; Kelentse, N.; Mokgethi, P.; Moraka, N.O.; Motswaledi, M.S.; Tawe, L.; et al. No Difference in the Prevalence of HIV-1 gag Cytotoxic T-Lymphocyte-Associated Escape Mutations in Viral Sequences from Early and Late Parts of the HIV-1 Subtype C Pandemic in Botswana. Vaccines 2023, 11, 1000. https://doi.org/10.3390/vaccines11051000
Mokaleng B, Choga WT, Bareng OT, Maruapula D, Ditshwanelo D, Kelentse N, Mokgethi P, Moraka NO, Motswaledi MS, Tawe L, et al. No Difference in the Prevalence of HIV-1 gag Cytotoxic T-Lymphocyte-Associated Escape Mutations in Viral Sequences from Early and Late Parts of the HIV-1 Subtype C Pandemic in Botswana. Vaccines. 2023; 11(5):1000. https://doi.org/10.3390/vaccines11051000
Chicago/Turabian StyleMokaleng, Baitshepi, Wonderful Tatenda Choga, Ontlametse Thato Bareng, Dorcas Maruapula, Doreen Ditshwanelo, Nametso Kelentse, Patrick Mokgethi, Natasha Onalenna Moraka, Modisa Sekhamo Motswaledi, Leabaneng Tawe, and et al. 2023. "No Difference in the Prevalence of HIV-1 gag Cytotoxic T-Lymphocyte-Associated Escape Mutations in Viral Sequences from Early and Late Parts of the HIV-1 Subtype C Pandemic in Botswana" Vaccines 11, no. 5: 1000. https://doi.org/10.3390/vaccines11051000
APA StyleMokaleng, B., Choga, W. T., Bareng, O. T., Maruapula, D., Ditshwanelo, D., Kelentse, N., Mokgethi, P., Moraka, N. O., Motswaledi, M. S., Tawe, L., Koofhethile, C. K., Moyo, S., Zachariah, M., & Gaseitsiwe, S. (2023). No Difference in the Prevalence of HIV-1 gag Cytotoxic T-Lymphocyte-Associated Escape Mutations in Viral Sequences from Early and Late Parts of the HIV-1 Subtype C Pandemic in Botswana. Vaccines, 11(5), 1000. https://doi.org/10.3390/vaccines11051000