
Production of Foot-and-Mouth Disease Type O and A Vaccine Antigens on a Pilot Scale and Determination of Optimal Amount of Antigen for Monovalent Vaccines
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Pilot-Scale Production of FMD Vaccine Antigens
2.3. Virus Titration
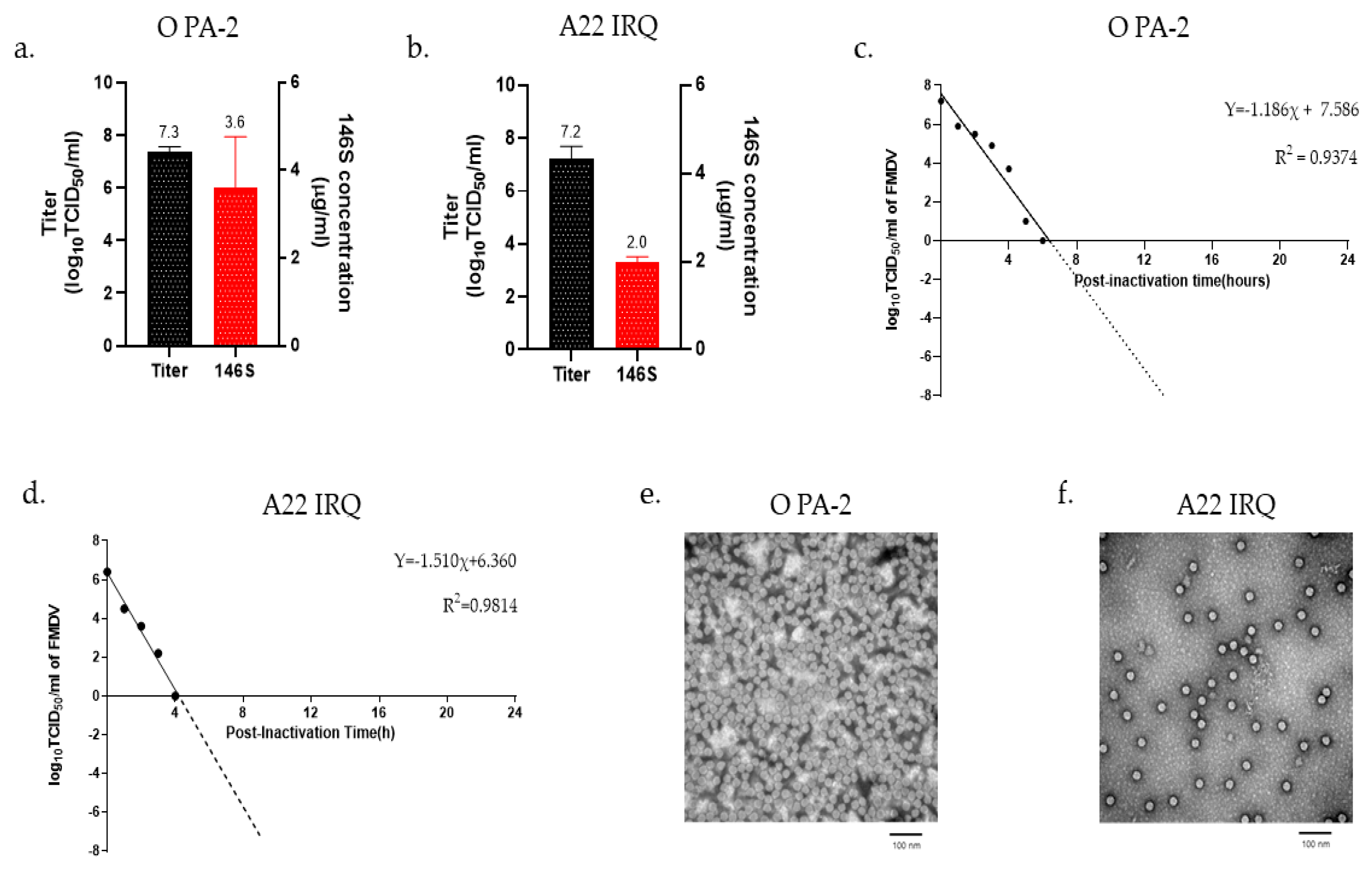
2.4. Quantification of FMDV Particles
2.5. Inactivation Kinetics
2.6. Transmission Electron Microscopy
2.7. Protective Efficacy of Monovalent Vaccines in Pigs
2.8. Virus Neutralization Test
2.9. Statistical Analysis
3. Results
3.1. Scaling up Production of FMD Vaccine Antigen
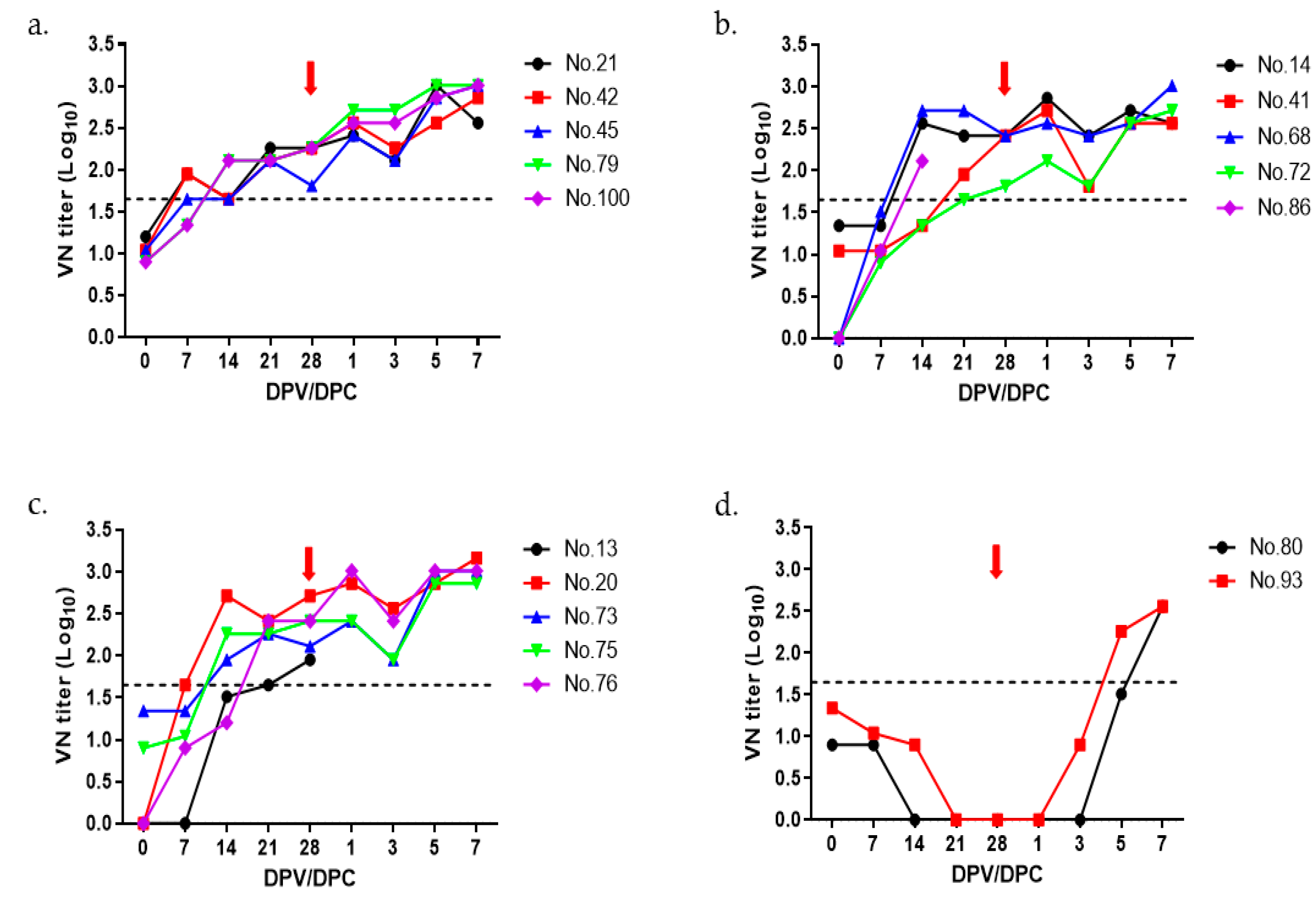
3.2. Immunogenicity and Protective Efficacy of O PA-2 Vaccine in Pigs
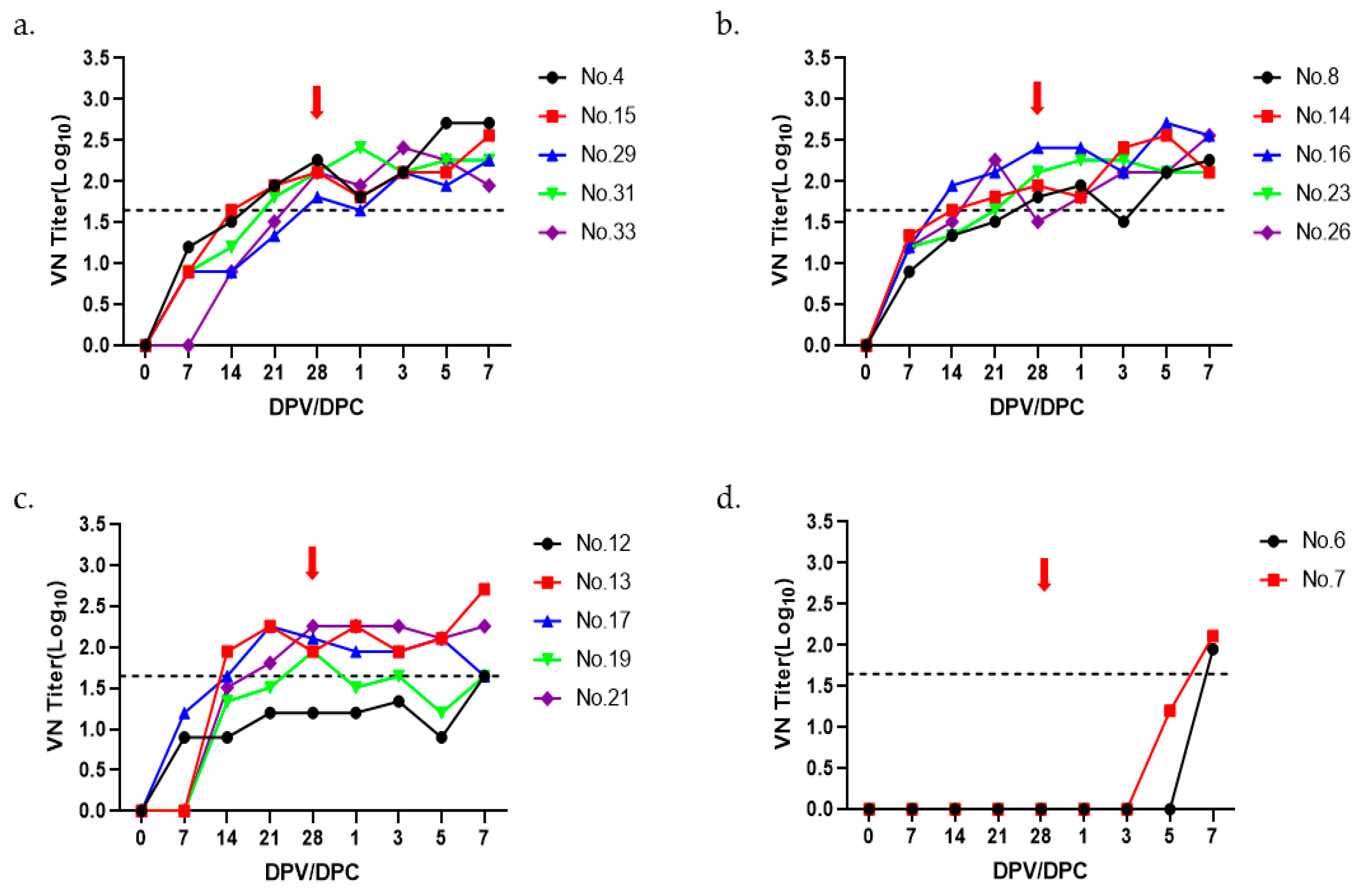
3.3. Immunogenicity and Protective Efficacy of A22 IRQ Vaccine in Pigs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knight-Jones, T.J.; Rushton, J. The economic impacts of foot and mouth disease—What are they, how big are they and where do they occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, P.W.; Grubman, M.J.; Baxt, B. Molecular basis of pathogenesis of FMDV. Virus Res. 2003, 91, 9–32. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.; Gibbs, E.; Horzinek, M.; Studdert, M.J. Veterinary Virology; Elsevier: San Diego, CA, USA, 1999; pp. 521–528. [Google Scholar]
- Geale, D.W.; Barnett, P.V.; Clarke, G.W.; Davis, J.; Kasari, T.R. A review of OIE country status recovery using vaccinate-to-live versus vaccinate-to-die foot-and-mouth disease response policies II: Waiting periods after emergency vaccination in FMD free countries. Transbound. Emerg. Dis. 2015, 62, 388–406. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, M.; Upadhyaya, S.; Aviso, S.; Babu, A.; Hutchings, G.; Parida, S. Selection of vaccine strains for serotype O foot-and-mouth disease viruses (2007–2012) circulating in Southeast Asia, East Asia and Far East. Vaccine 2017, 35, 7147–7153. [Google Scholar] [CrossRef] [PubMed]
- Singanallur, N.B.; Dekker, A.; Eblé, P.L.; van Hemert-Kluitenberg, F.; Weerdmeester, K.; Horsington, J.; Vosloo, W.W. Emergency Foot-and-Mouth Disease Vaccines A Malaysia 97 and A(22) Iraq 64 Offer Good Protection against Heterologous Challenge with A Variant Serotype A ASIA/G-IX/SEA-97 Lineage Virus. Vaccines 2020, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Lee, Y.J.; Kim, R.H.; Park, J.N.; Park, M.E.; Ko, M.K.; Choi, J.H.; Chu, J.Q.; Lee, K.N.; Kim, S.M.; et al. Rapid engineering of foot-and-mouth disease vaccine and challenge viruses. J. Virol. 2017, 91, e00155-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, M.K.; Jo, H.E.; Choi, J.H.; You, S.H.; Shin, S.H.; Jo, H.; Lee, M.J.; Kim, S.M.; Kim, B.; Park, J.H. Improved foot-and-mouth disease vaccine with O PanAsia-2 strain protect pigs against O/Jincheon/SKR/2014 originated from South Korea. Vaccine 2020, 38, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakopsychiatr. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Spitteler, M.A.; Romo, A.; Magi, N.; Seo, M.G.; Yun, S.J.; Barroumeres, F.; Régulier, E.G.; Bellinzoni, R. Validation of a high performance liquid chromatography method for quantitation of foot-and-mouth disease virus antigen in vaccines and vaccine manufacturing. Vaccine 2019, 37, 5288–5296. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.P.; Guzylack-Piriou, L.; Juillard, V.; Audonnet, J.C.; Doel, T.; Dawson, H.; Golde, W.T.; Gerber, H.; Peduto, N.; McCullough, K.C.; et al. Innate immune defenses induced by CpG do not promote vaccine-induced protection against foot-and-mouth disease virus in pigs. Clin. Vaccine Immunol. 2009, 16, 1151–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Organisation for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Chapter 3.1.8. Foot and Mouth Disease (Infection with Foot and Mouth Disease Virus). 2022. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 13 January 2023).
- Li, X.R.; Yang, Y.K.; Wang, R.B.; An, F.L.; Zhang, Y.D.; Nie, J.Q.; Ahamada, H.; Liu, X.X.; Liu, C.L.; Deng, Y.; et al. A scale-down model of 4000-L cell culture process for inactivated foot-and-mouth disease vaccine production. Vaccine 2019, 37, 6380–6389. [Google Scholar] [CrossRef] [PubMed]
- Corbic Ramljak, I.; Stanger, J.; Real-Hohn, A.; Dreier, D.; Wimmer, L.; Redlberger-Fritz, M.; Fischl, W.; Klingel, K.; Mihovilovic, M.D.; Blaas, D.; et al. Cellular N-myristoyltransferases play a crucial picornavirus genus-specific role in viral assembly, virion maturation, and infectivity. PLoS Pathog. 2018, 14, e1007203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.; Rodríguez, Y.Y.; Hershey, B.J.; Tadassa, Y.; Dodd, K.A.; Jia, W. Validation of a binary ethylenimine (BEI) inactivation procedure for biosafety treatment of foot-and-mouth disease viruses (FMDV), vesicular stomatitis viruses (VSV), and swine vesicular disease virus (SVDV). Vet. Microbiol. 2021, 252, 108928. [Google Scholar] [CrossRef] [PubMed]
- Aarthi, D.; Ananda Rao, K.; Robinson, R.; Srinivasan, V.A. Validation of binary ethyleneimine (BEI) used as an inactivant for foot and mouth disease tissue culture vaccine. Biologicals 2004, 32, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Rweyemamu, M.M.; Unehara, O.; Giorgi, W.; Medeiros, R.; Lucca, D.; Baltazar, M. Effect of formaldehyde and binary ethyleneimine (BEI) on the integrity of foot and mouth disease virus capsid. Rev. Sci. Tech. 1989, 8, 747–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barteling, S.J.; Meloen, R.H. A simple method for the quantification of 140-s particles of foot-and-mouth disease virus (FMDV). Arch. Gesamte Virusforsch. 1974, 45, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Doel, T.R. FMD vaccines. Virus Res. 2003, 91, 19. [Google Scholar] [CrossRef] [PubMed]
- Park, J.N.; Lee, S.Y.; Chu, J.Q.; Lee, Y.J.; Kim, R.H.; Lee, K.N.; Kim, S.M.; Tark, D.S.; Kim, B.; Park, J.H. Protection to homologous and heterologous challenge in pigs immunized with vaccine against foot-and-mouth disease type O caused an epidemic in East Asia during 2010/2011. Vaccine 2014, 32, 1882–1889. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Group | Pig ID | Virus-Neutralizing Titer | Clinical Score | Protection | ||||
|---|---|---|---|---|---|---|---|---|
| 0 dpv | 7 dpv | 14 dpv | 21 dpv | 28 dpv | ||||
| 20 µg | #21 | 1.20 | 1.95 | 1.65 | 2.26 | 2.26 | 0 | Yes |
| #42 | 1.04 | 1.95 | 1.65 | 2.11 | 2.26 | 0 | Yes | |
| #45 | 1.04 | 1.65 | 1.65 | 2.11 | 1.81 | 0 | Yes | |
| #79 | 0.9 | 1.34 | 2.11 | 2.11 | 2.26 | 0 | Yes | |
| #100 | 0.9 | 1.34 | 2.11 | 2.11 | 2.26 | 0 | Yes | |
| 15 µg | #14 | 1.34 | 1.34 | 2.56 | 2.41 | 2.41 | 0 | Yes |
| #41 | 1.04 | 1.04 | 1.34 | 1.95 | 2.41 | 0 | Yes | |
| #68 | <0.9 | 1.51 | 2.71 | 2.71 | 2.41 | 0 | Yes | |
| #72 | <0.9 | 0.9 | 1.34 | 1.65 | 1.81 | 0 | Yes | |
| #86 | <0.9 | 1.04 | 2.11 | - | - | - | - | |
| 10 µg | #13 | <0.9 | <0.9 | 1.51 | 1.65 | 1.95 | - | - |
| #20 | <0.9 | 1.65 | 2.71 | 2.41 | 2.71 | 0 | Yes | |
| #73 | 1.34 | 1.34 | 1.95 | 2.26 | 2.11 | 0 | Yes | |
| #75 | 0.9 | 1.04 | 2.26 | 2.26 | 2.41 | 0 | Yes | |
| #76 | <0.9 | 0.9 | 1.20 | 2.41 | 2.41 | 0 | Yes | |
| Control | #80 | 0.9 | 0.9 | <0.9 | <0.9 | <0.9 | 7 | No |
| #93 | 1.34 | 1.04 | 0.9 | <0.9 | <0.9 | 5 | No | |
| Group | Pig ID | Virus-Neutralizing Titer | Clinical Score | Protection | ||||
|---|---|---|---|---|---|---|---|---|
| 0 dpv | 7 dpv | 14 dpv | 21 dpv | 28 dpv | ||||
| 15 µg | #4 | <0.9 | 1.20 | 1.51 | 1.95 | 2.26 | 0 | Yes |
| #15 | <0.9 | 0.9 | 1.65 | 1.95 | 2.11 | 0 | Yes | |
| #29 | <0.9 | 0.9 | 0.9 | 1.34 | 1.81 | 0 | Yes | |
| #31 | <0.9 | 0.9 | 1.20 | 1.81 | 2.11 | 0 | Yes | |
| #33 | <0.9 | <0.9 | 0.9 | 1.51 | 2.11 | 0 | Yes | |
| 10 µg | #8 | <0.9 | 0.9 | 1.34 | 1.51 | 1.81 | 0 | Yes |
| #14 | <0.9 | 1.34 | 1.65 | 1.81 | 1.95 | 0 | Yes | |
| #16 | <0.9 | 1.20 | 1.95 | 2.11 | 2.41 | 0 | Yes | |
| #23 | <0.9 | 1.20 | 1.34 | 1.65 | 2.11 | 0 | Yes | |
| #26 | <0.9 | 1.20 | 1.51 | 2.26 | 1.51 | 0 | Yes | |
| 5 µg | #12 | <0.9 | 0.9 | 0.9 | 1.20 | 1.20 | 0 | Yes |
| #13 | <0.9 | <0.9 | 1.95 | 2.26 | 1.95 | 0 | Yes | |
| #17 | <0.9 | 1.20 | 1.65 | 2.26 | 2.11 | 0 | Yes | |
| #19 | <0.9 | <0.9 | 1.34 | 1.51 | 1.95 | 0 | Yes | |
| #21 | <0.9 | <0.9 | 1.51 | 1.81 | 2.26 | 0 | Yes | |
| Control | #6 | <0.9 | <0.9 | <0.9 | <0.9 | <0.9 | 6 | No |
| #7 | <0.9 | <0.9 | <0.9 | <0.9 | <0.9 | 4 | No | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.Y.; Lee, J.-H.; Yang, J.M.; Lee, S.-Y.; Park, S.Y.; Jin, J.S.; Kim, D.; Park, J.-W.; Park, J.-H.; Park, S.H.; et al. Production of Foot-and-Mouth Disease Type O and A Vaccine Antigens on a Pilot Scale and Determination of Optimal Amount of Antigen for Monovalent Vaccines. Vaccines 2023, 11, 1156. https://doi.org/10.3390/vaccines11071156
Kim JY, Lee J-H, Yang JM, Lee S-Y, Park SY, Jin JS, Kim D, Park J-W, Park J-H, Park SH, et al. Production of Foot-and-Mouth Disease Type O and A Vaccine Antigens on a Pilot Scale and Determination of Optimal Amount of Antigen for Monovalent Vaccines. Vaccines. 2023; 11(7):1156. https://doi.org/10.3390/vaccines11071156
Chicago/Turabian StyleKim, Jae Young, Ji-Hye Lee, Jong Min Yang, Seo-Yong Lee, Sun Young Park, Jong Sook Jin, Dohyun Kim, Jung-Won Park, Jong-Hyeon Park, Sang Hyun Park, and et al. 2023. "Production of Foot-and-Mouth Disease Type O and A Vaccine Antigens on a Pilot Scale and Determination of Optimal Amount of Antigen for Monovalent Vaccines" Vaccines 11, no. 7: 1156. https://doi.org/10.3390/vaccines11071156
APA StyleKim, J. Y., Lee, J.-H., Yang, J. M., Lee, S.-Y., Park, S. Y., Jin, J. S., Kim, D., Park, J.-W., Park, J.-H., Park, S. H., & Ko, Y.-J. (2023). Production of Foot-and-Mouth Disease Type O and A Vaccine Antigens on a Pilot Scale and Determination of Optimal Amount of Antigen for Monovalent Vaccines. Vaccines, 11(7), 1156. https://doi.org/10.3390/vaccines11071156