
Mtb-Specific HLA-E-Restricted T Cells Are Induced during Mtb Infection but Not after BCG Administration in Non-Human Primates and Humans
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. HLA-E TM Folding and Production
2.2. Human BCG Revaccination Study
2.3. Controlled Human BCG Infection Studies 1 and 2
2.4. Rhesus Macaque Study 1: Ethics, Animal Care, Vaccination and Pathology Assessment
2.5. Rhesus Macaque Study 2: Ethics, Animal Care, Vaccination, and Pathology Assessment
2.6. Cynomolgus Macaque Study: Ethics, Animal Care, Vaccination and Pathology Assessment
2.7. Flow Cytometry Staining, Acquisition and Analysis of NHP PBMC and BAL Samples
3. Results
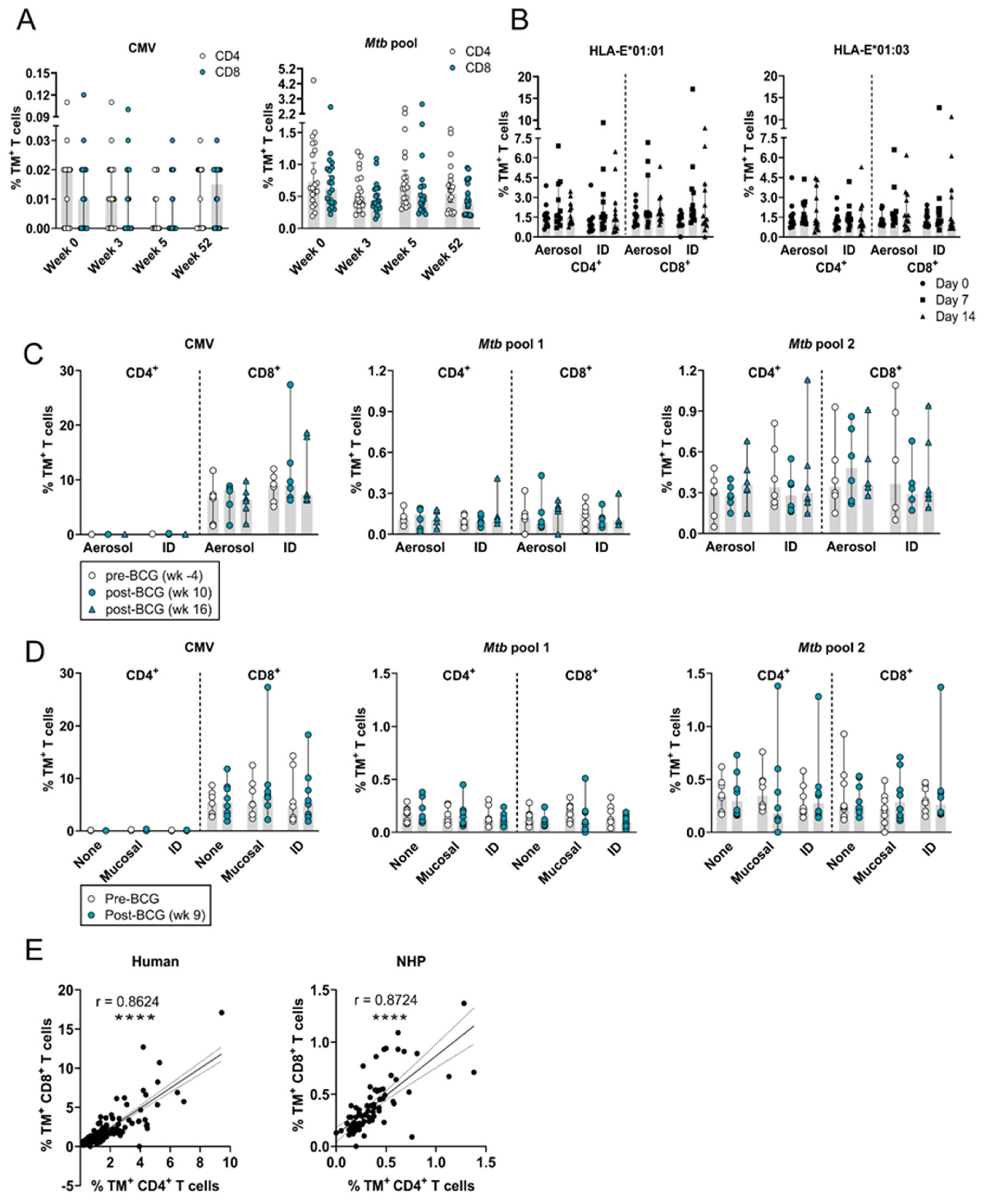
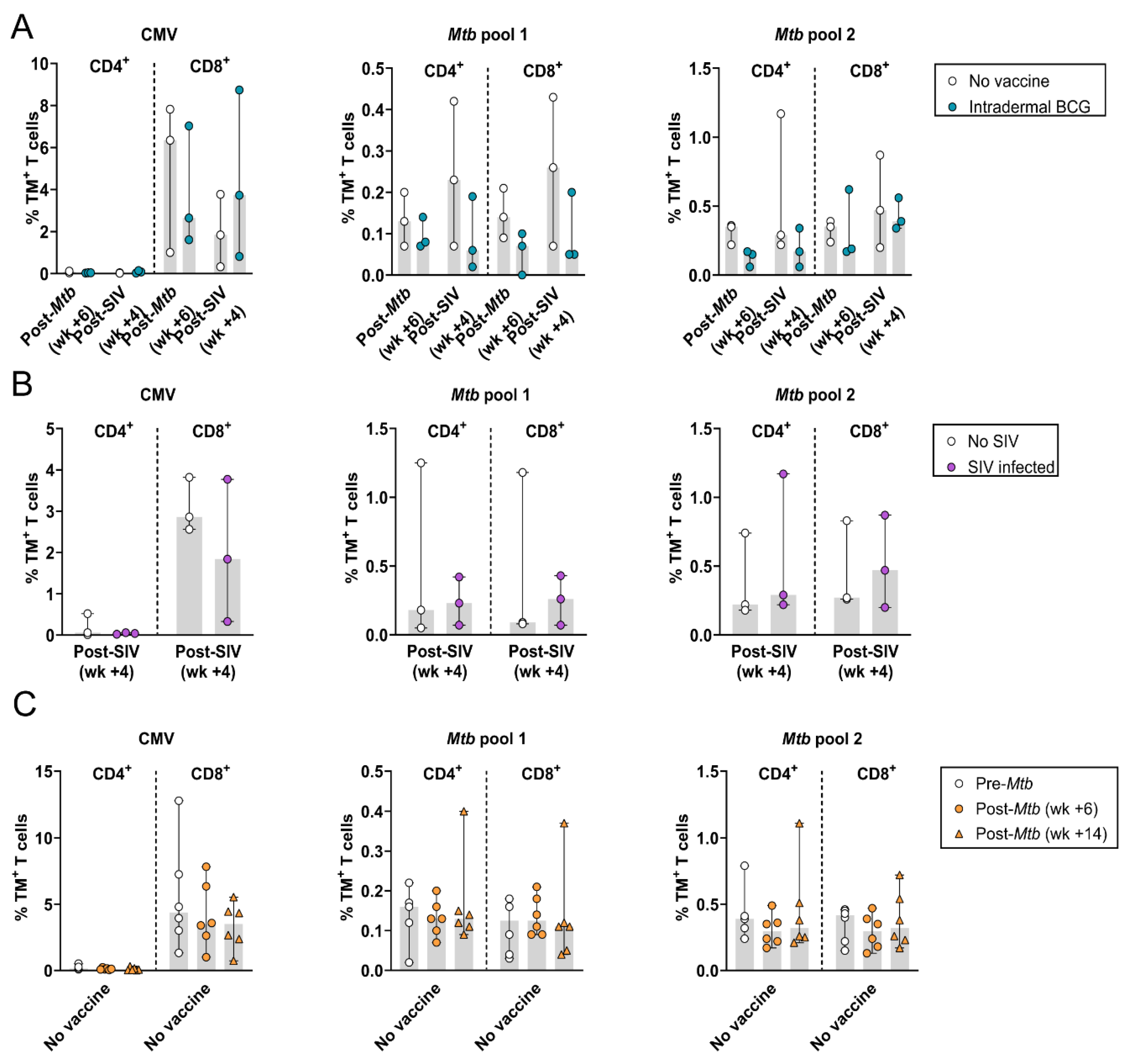
3.1. The Frequency of Circulating HLA-E–Mtb CD4+ and CD8+ T Cells Remains Stable in Humans and RMs after Receiving BCG
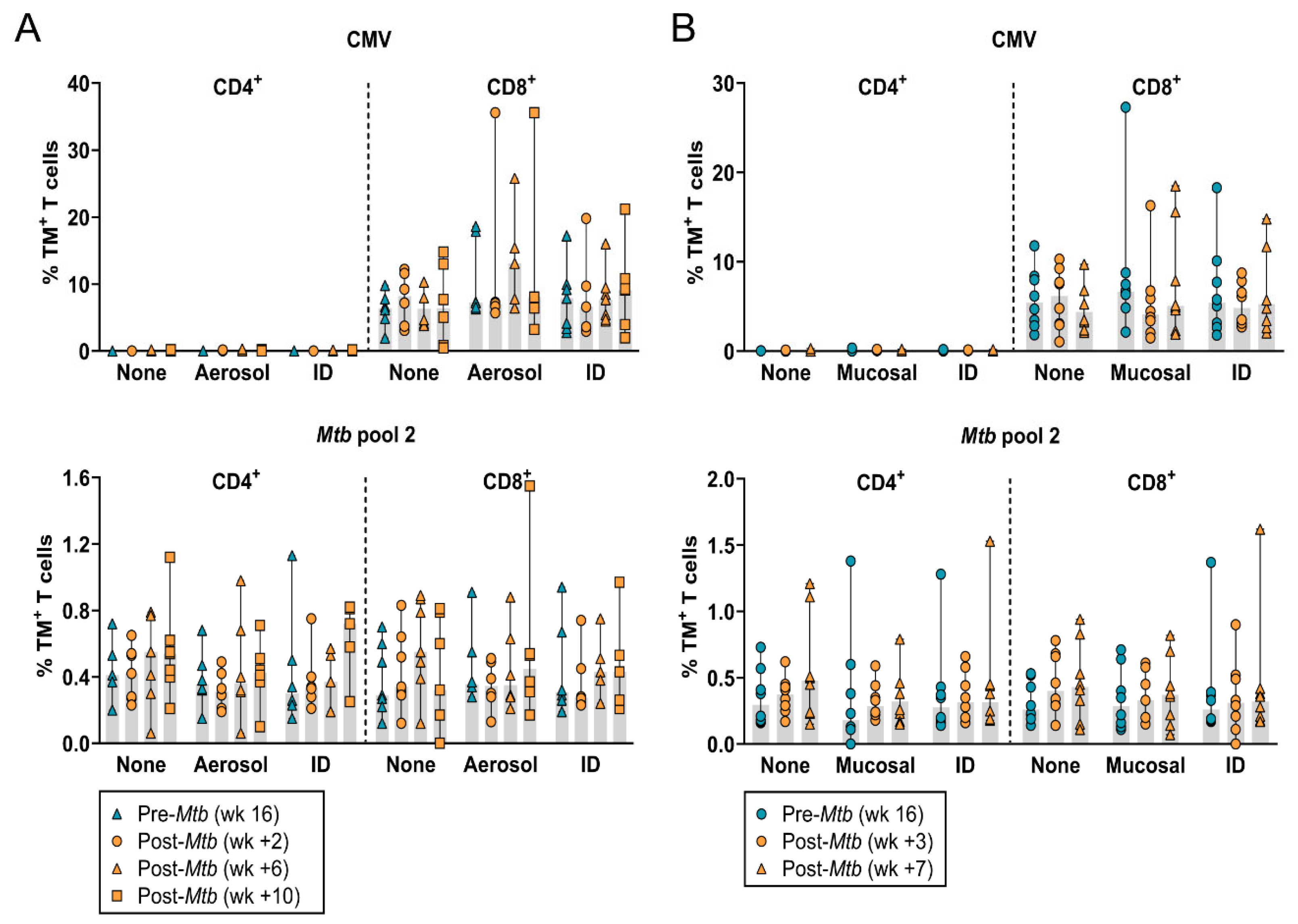
3.2. The Frequency of Circulating HLA-E–Mtb CD4+ and CD8+ T Cells Does Not Change after Mtb Challenge in Unvaccinated and BCG Vaccinated RMs
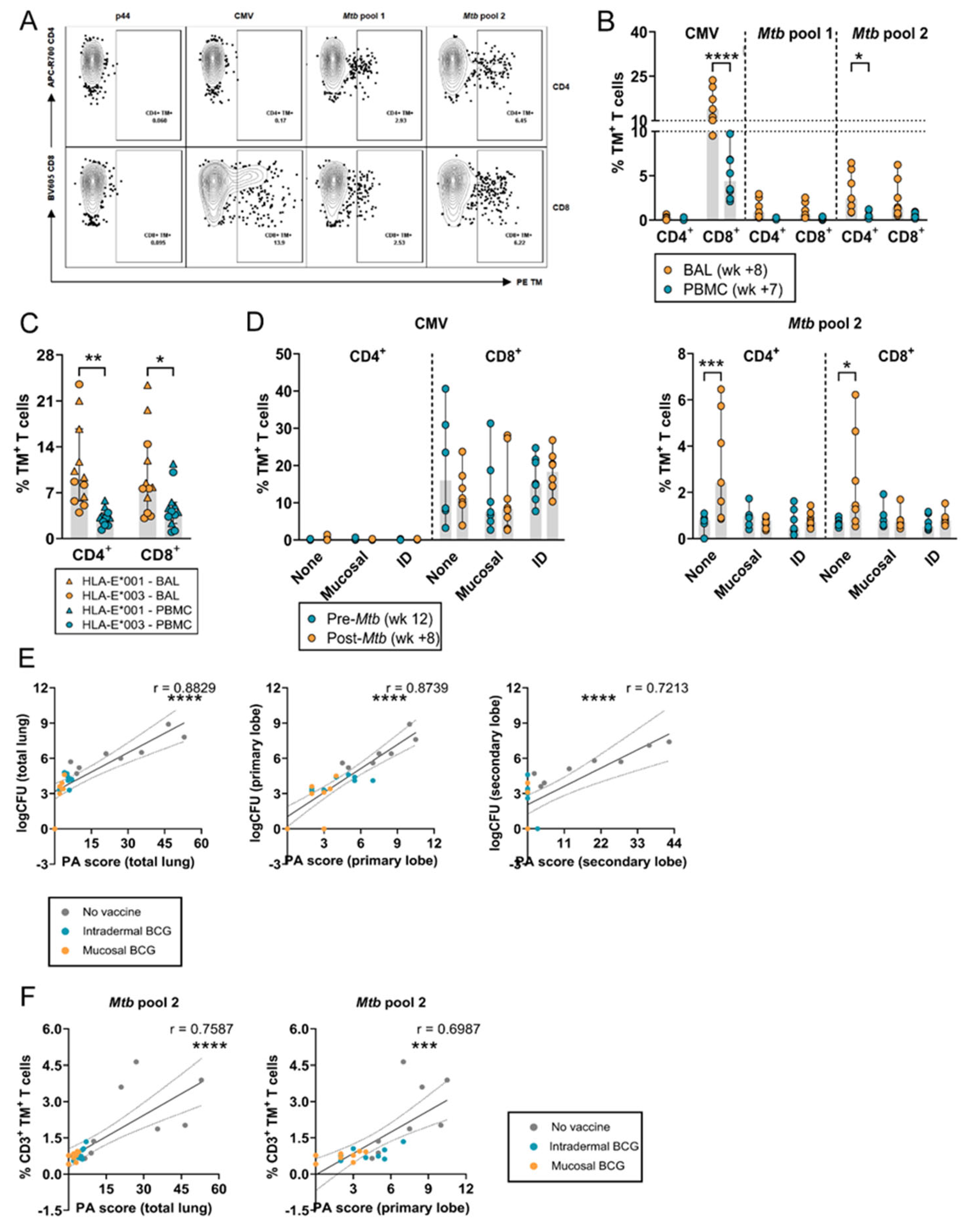
3.3. Increased Frequency of HLA-E–Mtb CD4+ and CD8+ T Cells in the BAL after Mtb Challenge in Unvaccinated RMs
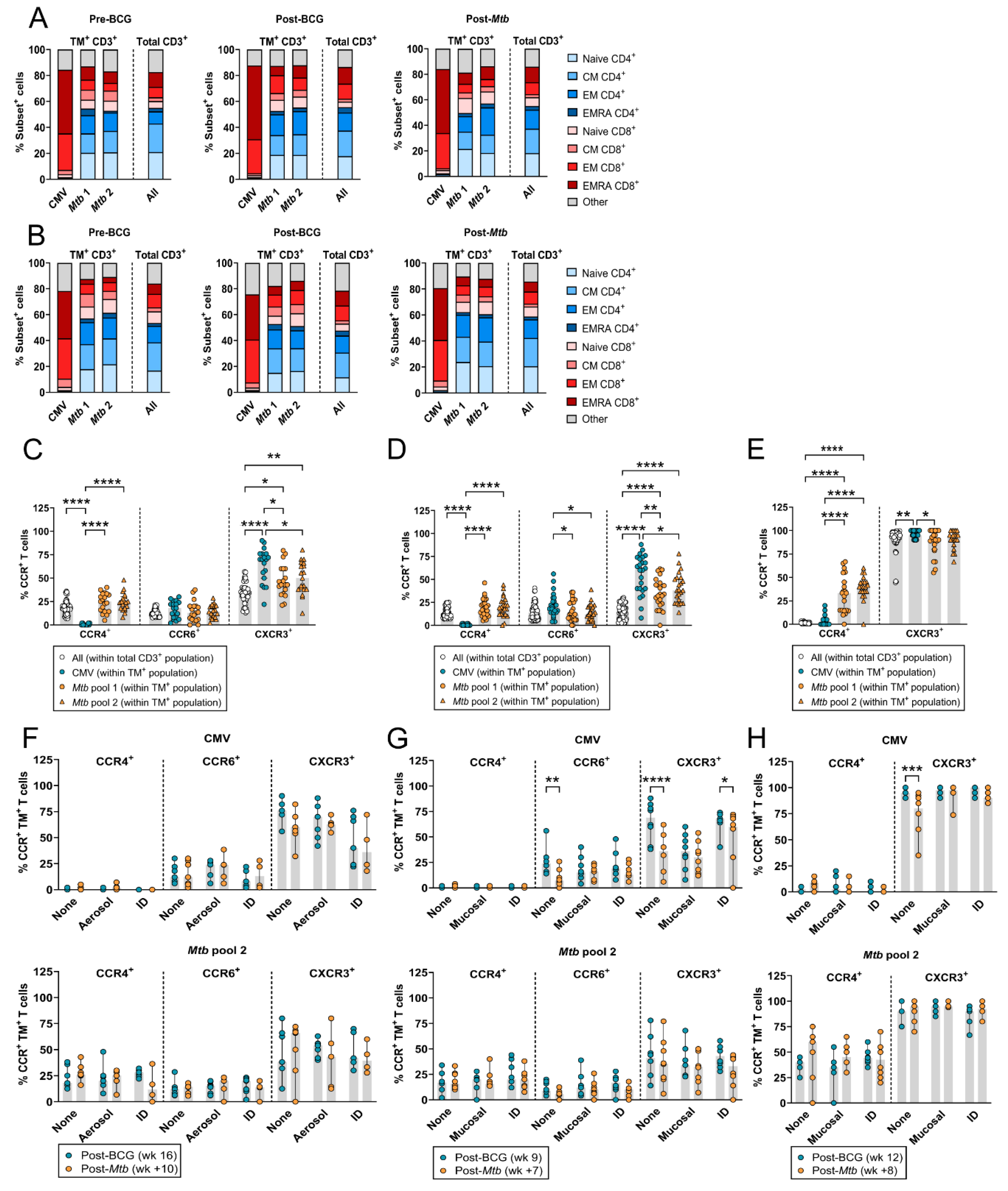
3.4. HLA-E–Mtb CD3+ T Cells Have a Similar Memory Phenotype but Altered Chemokine Receptor Expression Compared to Total T Cells
3.5. Simian Immunodeficiency Virus (SIV) Co-Infection in Mtb-Challenged Cynomolgus Macaques and HLA-E–Mtb CD4+ and CD8+ T Cell Frequencies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2023; World Health Organization: Geneva, Switzerland, 2023. [Google Scholar]
- Hunter, R.L.; Actor, J.K.; Hwang, S.A.; Khan, A.; Urbanowski, M.E.; Kaushal, D.; Jagannath, C. Pathogenesis and Animal Models of Post-Primary (Bronchogenic) Tuberculosis, A Review. Pathogens 2018, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Cardona, P.J.; Williams, A. Experimental animal modelling for TB vaccine development. Int. J. Infect. Dis. 2017, 56, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Van Rhijn, I.; Moody, D.B. Donor Unrestricted T Cells: A Shared Human T Cell Response. J. Immunol. 2015, 195, 1927–1932. [Google Scholar] [CrossRef] [PubMed]
- Kanevskiy, L.; Erokhina, S.; Kobyzeva, P.; Streltsova, M.; Sapozhnikov, A.; Kovalenko, E. Dimorphism of HLA-E and its Disease Association. Int. J. Mol. Sci. 2019, 20, 5496. [Google Scholar] [CrossRef] [PubMed]
- Strong, R.K.; Holmes, M.A.; Li, P.; Braun, L.; Lee, N.; Geraghty, D.E. HLA-E allelic variants. Correlating differential expression, peptide affinities, crystal structures, and thermal stabilities. J. Biol. Chem. 2003, 278, 5082–5090. [Google Scholar] [CrossRef]
- Lee, N.; Llano, M.; Carretero, M.; Ishitani, A.; Navarro, F.; Lopez-Botet, M.; Geraghty, D.E. HLA-E is a major ligand for the natural killer inhibitory receptor CD94/NKG2A. Proc. Natl. Acad. Sci. USA 1998, 95, 5199–5204. [Google Scholar] [CrossRef]
- Braud, V.M.; Allan, D.S.; O’Callaghan, C.A.; Soderstrom, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef]
- Voogd, L.; Ruibal, P.; Ottenhoff, T.H.M.; Joosten, S.A. Antigen presentation by MHC-E: A putative target for vaccination? Trends Immunol. 2022, 43, 355–365. [Google Scholar] [CrossRef]
- Voogd, L.; Drittij, A.; Dingenouts, C.K.E.; Franken, K.; Unen, V.V.; van Meijgaarden, K.E.; Ruibal, P.; Hagedoorn, R.S.; Leitner, J.A.; Steinberger, P.; et al. Mtb HLA-E-tetramer-sorted CD8(+) T cells have a diverse TCR repertoire. iScience 2024, 27, 109233. [Google Scholar] [CrossRef]
- Ruibal, P.; Franken, K.; van Meijgaarden, K.E.; van Wolfswinkel, M.; Derksen, I.; Scheeren, F.A.; Janssen, G.M.C.; van Veelen, P.A.; Sarfas, C.; White, A.D.; et al. Identification of HLA-E Binding Mycobacterium tuberculosis-Derived Epitopes through Improved Prediction Models. J. Immunol. 2022, 209, 1555–1565. [Google Scholar] [CrossRef]
- van Meijgaarden, K.E.; Haks, M.C.; Caccamo, N.; Dieli, F.; Ottenhoff, T.H.; Joosten, S.A. Human CD8+ T-cells recognizing peptides from Mycobacterium tuberculosis (Mtb) presented by HLA-E have an unorthodox Th2-like, multifunctional, Mtb inhibitory phenotype and represent a novel human T-cell subset. PLoS Pathog. 2015, 11, e1004671. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Sun, H.; Brackenridge, S.; Zhuang, X.; Wing, P.A.C.; Quastel, M.; Walters, L.; Garner, L.; Wang, B.; Yao, X.; et al. HLA-E-restricted SARS-CoV-2-specific T cells from convalescent COVID-19 patients suppress virus replication despite HLA class Ia down-regulation. Sci. Immunol. 2023, 8, eabl8881. [Google Scholar] [CrossRef] [PubMed]
- Salerno-Goncalves, R.; Fernandez-Vina, M.; Lewinsohn, D.M.; Sztein, M.B. Identification of a human HLA-E-restricted CD8+ T cell subset in volunteers immunized with Salmonella enterica serovar Typhi strain Ty21a typhoid vaccine. J. Immunol. 2004, 173, 5852–5862. [Google Scholar] [CrossRef] [PubMed]
- Vietzen, H.; Furlano, P.L.; Cornelissen, J.J.; Bohmig, G.A.; Jaksch, P.; Puchhammer-Stockl, E. HLA-E-restricted immune responses are crucial for the control of EBV infections and the prevention of PTLD. Blood 2023, 141, 1560–1573. [Google Scholar] [CrossRef] [PubMed]
- Doorduijn, E.M.; Sluijter, M.; Marijt, K.A.; Querido, B.J.; van der Burg, S.H.; van Hall, T. T cells specific for a TAP-independent self-peptide remain naive in tumor-bearing mice and are fully exploitable for therapy. Oncoimmunology 2018, 7, e1382793. [Google Scholar] [CrossRef]
- Doorduijn, E.M.; Sluijter, M.; Querido, B.J.; Seidel, U.J.E.; Oliveira, C.C.; van der Burg, S.H.; van Hall, T. T Cells Engaging the Conserved MHC Class Ib Molecule Qa-1(b) with TAP-Independent Peptides Are Semi-Invariant Lymphocytes. Front Immunol 2018, 9, 60. [Google Scholar] [CrossRef]
- Yang, H.; Rei, M.; Brackenridge, S.; Brenna, E.; Sun, H.; Abdulhaqq, S.; Liu, M.K.P.; Ma, W.; Kurupati, P.; Xu, X.; et al. HLA-E-restricted, Gag-specific CD8(+) T cells can suppress HIV-1 infection, offering vaccine opportunities. Sci. Immunol. 2021, 6, eabg1703. [Google Scholar] [CrossRef]
- Jorgensen, P.B.; Livbjerg, A.H.; Hansen, H.J.; Petersen, T.; Hollsberg, P. Epstein-Barr virus peptide presented by HLA-E is predominantly recognized by CD8(bright) cells in multiple sclerosis patients. PLoS ONE 2012, 7, e46120. [Google Scholar] [CrossRef]
- Hansen, S.G.; Marshall, E.E.; Malouli, D.; Ventura, A.B.; Hughes, C.M.; Ainslie, E.; Ford, J.C.; Morrow, D.; Gilbride, R.M.; Bae, J.Y.; et al. A live-attenuated RhCMV/SIV vaccine shows long-term efficacy against heterologous SIV challenge. Sci. Transl. Med. 2019, 11, eaaw2607. [Google Scholar] [CrossRef]
- Malouli, D.; Gilbride, R.M.; Wu, H.L.; Hwang, J.M.; Maier, N.; Hughes, C.M.; Newhouse, D.; Morrow, D.; Ventura, A.B.; Law, L.; et al. Cytomegalovirus-vaccine-induced unconventional T cell priming and control of SIV replication is conserved between primate species. Cell Host Microbe 2022, 30, 1207–1218.e1207. [Google Scholar] [CrossRef]
- Verweij, M.C.; Hansen, S.G.; Iyer, R.; John, N.; Malouli, D.; Morrow, D.; Scholz, I.; Womack, J.; Abdulhaqq, S.; Gilbride, R.M.; et al. Modulation of MHC-E transport by viral decoy ligands is required for RhCMV/SIV vaccine efficacy. Science 2021, 372, eabe9233. [Google Scholar] [CrossRef] [PubMed]
- Nattermann, J.; Nischalke, H.D.; Hofmeister, V.; Kupfer, B.; Ahlenstiel, G.; Feldmann, G.; Rockstroh, J.; Weiss, E.H.; Sauerbruch, T.; Spengler, U. HIV-1 infection leads to increased HLA-E expression resulting in impaired function of natural killer cells. Antivir. Ther. 2005, 10, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Gela, A.; Murphy, M.; Rodo, M.; Hadley, K.; Hanekom, W.A.; Boom, W.H.; Johnson, J.L.; Hoft, D.F.; Joosten, S.A.; Ottenhoff, T.H.M.; et al. Effects of BCG vaccination on donor unrestricted T cells in two prospective cohort studies. EBioMedicine 2022, 76, 103839. [Google Scholar] [CrossRef] [PubMed]
- Hatherill, M.; Geldenhuys, H.; Pienaar, B.; Suliman, S.; Chheng, P.; Debanne, S.M.; Hoft, D.F.; Boom, W.H.; Hanekom, W.A.; Johnson, J.L. Safety and reactogenicity of BCG revaccination with isoniazid pretreatment in TST positive adults. Vaccine 2014, 32, 3982–3988. [Google Scholar] [CrossRef]
- Satti, I.; Marshall, J.L.; Harris, S.A.; Wittenberg, R.; Tanner, R.; Lopez Ramon, R.; Wilkie, M.; Ramos Lopez, F.; Riste, M.; Wright, D.; et al. Safety of a controlled human infection model of tuberculosis with aerosolised, live-attenuated Mycobacterium bovis BCG versus intradermal BCG in BCG-naive adults in the UK: A dose-escalation, randomised, controlled, phase 1 trial. Lancet Infect. Dis. 2024, 24, 909–921. [Google Scholar] [CrossRef]
- White, A.D.; Sarfas, C.; Sibley, L.S.; Gullick, J.; Clark, S.; Rayner, E.; Gleeson, F.; Catala, M.; Nogueira, I.; Cardona, P.J.; et al. Protective Efficacy of Inhaled BCG Vaccination Against Ultra-Low Dose Aerosol M. tuberculosis Challenge in Rhesus Macaques. Pharmaceutics 2020, 12, 394. [Google Scholar] [CrossRef]
- Dijkman, K.; Sombroek, C.C.; Vervenne, R.A.W.; Hofman, S.O.; Boot, C.; Remarque, E.J.; Kocken, C.H.M.; Ottenhoff, T.H.M.; Kondova, I.; Khayum, M.A.; et al. Prevention of tuberculosis infection and disease by local BCG in repeatedly exposed rhesus macaques. Nat. Med. 2019, 25, 255–262. [Google Scholar] [CrossRef]
- White, A.D.; Sibley, L.; Gullick, J.; Sarfas, C.; Clark, S.; Fagrouch, Z.; Verschoor, E.; Salguero, F.J.; Dennis, M.; Sharpe, S. TB and SIV Coinfection; a Model for Evaluating Vaccine Strategies against TB Reactivation in Asian Origin Cynomolgus Macaques: A Pilot Study Using BCG Vaccination. Vaccines 2021, 9, 945. [Google Scholar] [CrossRef]
- van Wolfswinkel, M.; van Meijgaarden, K.E.; Ottenhoff, T.H.M.; Niewold, P.; Joosten, S.A. Extensive flow cytometric immunophenotyping of human PBMC incorporating detection of chemokine receptors, cytokines and tetramers. Cytom. A 2023, 103, 600–610. [Google Scholar] [CrossRef]
- Walters, L.C.; Harlos, K.; Brackenridge, S.; Rozbesky, D.; Barrett, J.R.; Jain, V.; Walter, T.S.; O’Callaghan, C.A.; Borrow, P.; Toebes, M.; et al. Pathogen-derived HLA-E bound epitopes reveal broad primary anchor pocket tolerability and conformationally malleable peptide binding. Nat. Commun. 2018, 9, 3137. [Google Scholar] [CrossRef]
- Darrah, P.A.; Zeppa, J.J.; Maiello, P.; Hackney, J.A.; Wadsworth, M.H., 2nd; Hughes, T.K.; Pokkali, S.; Swanson, P.A., 2nd; Grant, N.L.; Rodgers, M.A.; et al. Prevention of tuberculosis in macaques after intravenous BCG immunization. Nature 2020, 577, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Prezzemolo, T.; van Meijgaarden, K.E.; Franken, K.; Caccamo, N.; Dieli, F.; Ottenhoff, T.H.M.; Joosten, S.A. Detailed characterization of human Mycobacterium tuberculosis specific HLA-E restricted CD8(+) T cells. Eur. J. Immunol. 2018, 48, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L.C.; Westall, G.P.; Widjaja, J.M.; Mifsud, N.A.; Nguyen, T.H.; Meehan, A.C.; Kotsimbos, T.C.; Brooks, A.G. The Presence of HLA-E-Restricted, CMV-Specific CD8+ T Cells in the Blood of Lung Transplant Recipients Correlates with Chronic Allograft Rejection. PLoS ONE 2015, 10, e0135972. [Google Scholar] [CrossRef] [PubMed]
- Pietra, G.; Romagnani, C.; Mazzarino, P.; Falco, M.; Millo, E.; Moretta, A.; Moretta, L.; Mingari, M.C. HLA-E-restricted recognition of cytomegalovirus-derived peptides by human CD8+ cytolytic T lymphocytes. Proc. Natl. Acad. Sci. USA 2003, 100, 10896–10901. [Google Scholar] [CrossRef]
- Mazzarino, P.; Pietra, G.; Vacca, P.; Falco, M.; Colau, D.; Coulie, P.; Moretta, L.; Mingari, M.C. Identification of effector-memory CMV-specific T lymphocytes that kill CMV-infected target cells in an HLA-E-restricted fashion. Eur. J. Immunol. 2005, 35, 3240–3247. [Google Scholar] [CrossRef]
- Hansen, S.G.; Wu, H.L.; Burwitz, B.J.; Hughes, C.M.; Hammond, K.B.; Ventura, A.B.; Reed, J.S.; Gilbride, R.M.; Ainslie, E.; Morrow, D.W.; et al. Broadly targeted CD8(+) T cell responses restricted by major histocompatibility complex E. Science 2016, 351, 714–720. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Name | Sequence | Allele | Fluorochrome |
|---|---|---|---|
| Human BCG infection studies 1 and 2: Mtb pool and CMV | |||
| p34 | VMTTVLATL | HLA-E*01:01 + HLA-E*01:03 separately | PE |
| p55 | VMATRRNVL | HLA-E*01:01 + HLA-E*01:03 separately | PE |
| p62 | RMPPLGHEL | HLA-E*01:01 + HLA-E*01:03 separately | PE |
| p68 | VLRPGGHFL | HLA-E*01:01 + HLA-E*01:03 separately | PE |
| CMV | VLAPRTLLL | HLA-E*01:01 + HLA-E*01:03 separately | APC |
| Human BCG revaccination study: Mtb pool and CMV | |||
| p34 | VMTTVLATL | HLA-E*01:01 + HLA-E*01:03 combined | PE |
| p55 | VMATRRNVL | HLA-E*01:01 + HLA-E*01:03 combined | PE |
| p62 | RMPPLGHEL | HLA-E*01:01 + HLA-E*01:03 combined | PE |
| p68 | VLRPGGHFL | HLA-E*01:01 + HLA-E*01:03 combined | PE |
| CMV | VLAPRTLLL | HLA-E*01:01 + HLA-E*01:03 combined | APC |
| NHP studies: Mtb pool 1 | |||
| p55 | VMATRRNVL | HLA-E*01:03 | PE |
| p62 | RMPPLGHEL | HLA-E*01:03 | PE |
| NHP studies: Mtb pool 2 | |||
| MTBHLAE_31 | VLPAKLILM | HLA-E*01:03 | PE |
| MTBHLAE_34 | LLPIKIPLI | HLA-E*01:03 | PE |
| MTBHLAE_63 | ILAFEAPEL | HLA-E*01:03 | PE |
| MTBHLAE_93 | RLEAVVMLL | HLA-E*01:03 | PE |
| NHP studies: CMV | |||
| CMV | VLAPRTLLL | HLA-E*01:03 | PE |
| NHP studies: p44 | |||
| p44 | RLPAKAPLL | HLA-E*01:03 | PE |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voogd, L.; van Wolfswinkel, M.; Satti, I.; White, A.D.; Dijkman, K.; Gela, A.; van Meijgaarden, K.E.; Franken, K.L.M.C.; Marshall, J.L.; Ottenhoff, T.H.M.; et al. Mtb-Specific HLA-E-Restricted T Cells Are Induced during Mtb Infection but Not after BCG Administration in Non-Human Primates and Humans. Vaccines 2024, 12, 1129. https://doi.org/10.3390/vaccines12101129
Voogd L, van Wolfswinkel M, Satti I, White AD, Dijkman K, Gela A, van Meijgaarden KE, Franken KLMC, Marshall JL, Ottenhoff THM, et al. Mtb-Specific HLA-E-Restricted T Cells Are Induced during Mtb Infection but Not after BCG Administration in Non-Human Primates and Humans. Vaccines. 2024; 12(10):1129. https://doi.org/10.3390/vaccines12101129
Chicago/Turabian StyleVoogd, Linda, Marjolein van Wolfswinkel, Iman Satti, Andrew D. White, Karin Dijkman, Anele Gela, Krista E. van Meijgaarden, Kees L. M. C. Franken, Julia L. Marshall, Tom H. M. Ottenhoff, and et al. 2024. "Mtb-Specific HLA-E-Restricted T Cells Are Induced during Mtb Infection but Not after BCG Administration in Non-Human Primates and Humans" Vaccines 12, no. 10: 1129. https://doi.org/10.3390/vaccines12101129