The Live Attenuated Vaccine Strain “ARRIAH” Completely Protects Goats from a Virulent Lineage IV Field Strain of Peste Des Petits Ruminants Virus

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vaccine Strains
2.2. Challenge Stain
2.3. Experimental Design
2.4. Ethics Statement
2.5. Clinical Monitoring
2.6. Assays
3. Results
- –
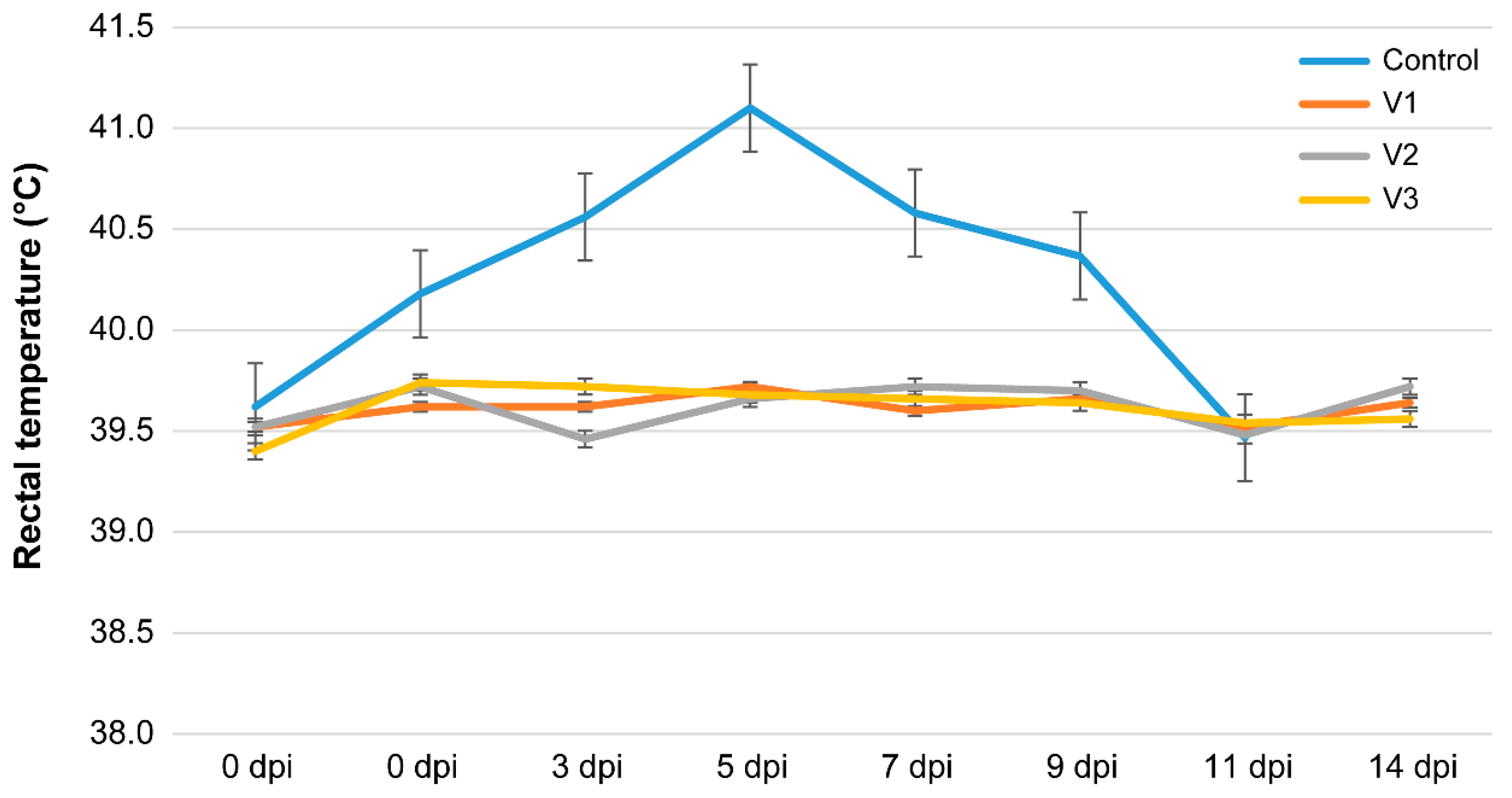
- An increase in rectal temperature to 40–41 °C was observed at 2–4 dpi;
- –
- Apathy and refusal of feed was observed on 3–6 dpi; moreover, the animals lied down a lot and rose reluctantly;
- –
- An increase in the size of lymph nodes was recorded at 5–7 dpi, and serous discharge from the nostrils appeared in some animals;
- –
- At 6–9 dpi, signs of damage to the respiratory system began to appear: serous discharge from the nostrils, hyperemia of the nasal mucosa, wheezing when breathing, coughing, and shortness of breath after running; animals no. three and four lied down and had a cough, shallow breathing, and high body temperature;
- –
- At days 10–12, animals no. three and four were euthanized due to their severe clinical condition (bilateral pneumonia). The remaining animals in the control group showed pronounced clinical signs of respiratory pathology. In addition, diarrhea was recorded in one animal.
3.1. Detection of PPRV RNA in Blood Samples and Nasal Swabs
3.2. Detection of Antibodies against PPRV
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gibbs, E.P.; Taylor, W.P.; Lawman, M.J.; Bryant, J. Classification of peste des petits ruminants virus as the fourth member of the genus Morbillivirus. Intervirology 1979, 11, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Banyard, A.C.; Parida, S.; Batten, C.; Oura, C.; Kwiatek, O.; Libeau, G. Global distribution of peste des petits ruminants virus and prospects for improved diagnosis and control. J. Gen. Virol. 2010, 91, 2885–2897. [Google Scholar] [CrossRef] [PubMed]
- Dundon, W.G.; Adombi, C.; Waqas, A.; Otsyina, H.R.; Arthur, C.T.; Silber, R.; Loitsch, A.; Diallo, A. Full genome sequence of a peste des petits ruminants virus (PPRV) from Ghana. Virus Genes 2014, 49, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, M.; Rajput, Z.I.; Arshed, M.J.; Sarwar, G.; Ali, Q. Evidence of peste des petits ruminants virus (PPRV) infection in Sindh Ibex (Capra aegagrus blythi) in Pakistan as confirmed by detection of antigen and antibody. Trop. Anim. Health Prod. 2011, 43, 745–747. [Google Scholar] [CrossRef]
- Shahriari, R.; Khodakaram-Tafti, A.; Mohammadi, A. Molecular characterization of Peste des Petits ruminants virus isolated from four outbreaks occurred in southern Iran. BMC Vet. Res. 2019, 15, 177. [Google Scholar] [CrossRef]
- Parida, S.; Muniraju, M.; Mahapatra, M.; Muthuchelvan, D.; Buczkowski, H.; Banyard, A.C. Peste des petits ruminants. Vet. Microbiol. 2015, 181, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Njeumi, F.; Parida, S.; Benfield, C.T.O. Progress towards eradication of peste des petits ruminants through vaccination. Viruses 2021, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Govindaraj, G.N.; Balamurugan, V.; Reddy, G.B.M.; Yogisharadhya, R.; Reddy, T.S.; Naveenkumar, G.S.; Kumar, K.V.; Chaithra, H.R.; Bi, A.Z.; Parida, S.; et al. Towards eradication of PPR: Disease status, economic cost and perception of veterinarians in Karnataka, India. Animals 2023, 13, 778. [Google Scholar] [CrossRef]
- Njeumi, F.; Bailey, D.; Soula, J.J.; Diop, B.; Tekola, B.G. Eradicating the scourge of peste des petits ruminants from the world. Viruses 2020, 12, 313. [Google Scholar] [CrossRef]
- Aboah, J.; Apolloni, A.; Duboz, R.; Wieland, B.; Kotchofa, P.; Okoth, E.; Dione, M. Ex-ante impact of pest des petits ruminant control on micro and macro socioeconomic indicators in Senegal: A system dynamics modelling approach. PLoS ONE 2023, 18, e0287386. [Google Scholar] [CrossRef]
- Jones, B.A.; Rich, K.M.; Mariner, J.C.; Anderson, J.; Jeggo, M.; Thevasagayam, S.; Cai, Y.; Peters, A.R.; Roeder, P. The economic impact of eradicating peste des petits ruminants: A benefit-cost analysis. PLoS ONE 2016, 11, e0149982. [Google Scholar] [CrossRef]
- Felix, N. Current scenario and control initiatives for PPR at global, regional and country level according to the risk factors and socioeconomic impact. In Proceedings of the Second Regional Conference on Progressive Control of Peste Des Petits Ruminants in South Asia, Kathmandu, Nepal, 19–20 December 2013. [Google Scholar]
- Abraham, G.; Sintayehu, A.; Libeau, G.; Albina, E.; Roger, F.; Laekemariam, Y.; Abayneh, D.; Awoke, K.M. Antibody seroprevalences against peste des petits ruminants (PPR) virus in camels, cattle, goats and sheep in Ethiopia. Prev. Vet. Med. 2005, 70, 51–57. [Google Scholar] [CrossRef]
- Munir, M.; Zohari, S.; Berg, M. Molecular Biology and Pathogenesis of Peste Des Petitis Ruminants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–152. [Google Scholar]
- Dundon, W.G.; Diallo, A.; Cattoli, G. Peste des petits ruminants in Africa: A review of currently available molecular epidemiological data, 2020. Arch. Virol. 2020, 165, 2147–2163. [Google Scholar] [CrossRef] [PubMed]
- Woma, T.Y.; Adombi, C.M.; Yu, D.; Qasim, A.M.; Sabi, A.A.; Maurice, N.A.; Olaiya, O.D.; Loitsch, A.; Bailey, D.; Shamaki, D.; et al. Co-circulation of peste-des-petits-ruminants virus asian lineage IV with Lineage II in Nigeria. Transbound. Emerg. Dis. 2016, 63, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Sprygin, A.; Sainnokhoi, T.; Gombo-Ochir, D.; Tserenchimed, T.; Tsolmon, A.; Ankhanbaatar, U.; Krotova, A.; Shumilova, I.; Shalina, K.; Prutnikov, P.; et al. Outbreak of peste des petits ruminants in sheep in Mongolia, 2021. Transbound. Emerg. Dis. 2022, 69, 1695–1697. [Google Scholar] [CrossRef]
- Kwiatek, O.; Ali, Y.H.; Saeed, I.K.; Khalafalla, A.I.; Mohamed, O.I.; Obeida, A.A.; Abdelrahman, M.B.; Osman, H.M.; Taha, K.M.; Abbas, Z.; et al. Asian lineage of peste des petits ruminants virus, Africa. Emerg. Infect. Dis. 2011, 17, 1223–1231. [Google Scholar] [CrossRef] [PubMed]
- Benfield, C.T.O.; Legnardi, M.; Mayen, F.; Almajali, A.; Cinardi, G.; Wisser, D.; Chaka, H.; Njeumi, F. Peste des petits ruminants in the Middle East: Epidemiological situation and status of control and eradication activities after the first phase of the PPR global eradication program (2017-2021). Animals 2023, 13, 1196. [Google Scholar] [CrossRef]
- Diallo, A.; Taylor, W.P.; Lefèvre, P.C.; Provost, A. Attenuation of a strain of pest of small ruminants virus: Candidate for a live homologous vaccine. Rev. Elev. Med. Vet. Pays. Trop. 1989, 42, 311–319. [Google Scholar] [CrossRef]
- Sen, A.; Saravanan, P.; Balamurugan, V.; Rajak, K.K.; Sudhakar, S.B.; Bhanuprakash, V.; Parida, S.; Singh, R.K. Vaccines against peste des petits ruminants virus. Expert. Rev. Vaccines 2010, 9, 785–796. [Google Scholar] [CrossRef]
- Singh, R.P.; Bandyopadhyay, S.K. Peste des petits ruminants vaccine and vaccination in India: Sharing experience with disease endemic countries. Virus Disease 2015, 26, 215–224. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Haider, N.; Gurley, E.S.; Ahmed, S.; Osmani, M.G.; Hossain, M.B.; Islam, A.; Khan, S.A.; Hossain, M.E.; Epstein, J.H.; et al. Epidemiology and genetic characterization of peste des petits ruminants virus in Bangladesh. Vet. Med. Sci. 2018, 4, 161–171. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Animal Health and Welfare (AHAW). Scientific opinion on peste des petits ruminants. EFSA J. 2015, 13, 94. [Google Scholar]
- Scherbinin, S.V.; Karaulov, A.K.; Zakharov, V.M. Risk analysis of peste des petits ruminants introduction into the territory of the Russian Federation. Vet. Sci. Today 2017, 4, 17–22. (In Russian) [Google Scholar]
- Kwiatek, O.; Libeau, G.; Guendouz, S.; Corbanini, C.; Gogin, A.; Lunitsin, A.; Sindryakova, I.; Koblasov, D.; Bataille, A. Genomic characterization of peste des petits ruminants vaccine seed “45G37/35-k”, Russia. Vet. Res. 2022, 53, 79. [Google Scholar] [CrossRef] [PubMed]
- Pope, R.A.; Parida, S.; Bailey, D.; Brownlie, J.; Barrett, T.; Banyard, A.C. Early events following experimental infection with peste-des-petits ruminants virus suggest immune cell targeting. PLoS ONE 2013, 8, e55830. [Google Scholar] [CrossRef] [PubMed]
- Cartn-Rojas, A. Transboundary animal diseases and international trade. Int. Trade Econ. Policy Perspect. 2012, 143, 166. [Google Scholar]
- De Haan, L.; Kimani, N.; Rushton, J.; Lubroth, J. Why is small ruminant health important—Peste des petits ruminants and its impact on poverty and economics? In Peste Des Petits Ruminants Virus; Springer: Berlin/Heidelberg, Germany, 2015; pp. 195–226. [Google Scholar]
- Kihu, S.M.; Gitao, G.C.; Bebora, L.C.; Njenga, M.J.; Wairire, G.G.; Maingi, N.; Wahome, R.G. Economic losses associated with peste des petits ruminants in Turkana County Kenya. Pastoralism 2015, 5, 9. [Google Scholar] [CrossRef]
- OIE and FAO. FAO/OIE Guidelines for the Control and Prevention of Peste Des Petits Ruminants (ppr) in Wildlife Populations; OIE and FAO: Rome, Italy, 2021. [Google Scholar]
- Hodgson, S.; Moffat, K.; Hill, H.; Flannery, J.T.; Graham, S.P.; Baron, M.D.; Darpel, K.E. Comparison of the immunogenicities and cross-lineage efficacies of live attenuated peste des petits ruminants virus vaccines PPRV/Nigeria/75/1 and PPRV/Sungri/96. J. Virol. 2018, 92, e01471-18. [Google Scholar] [CrossRef]
- Diev, V.I.; Kapuskin, E.V.; Kukushkina, M.S. Study of antigenic and immunogenic properties of an experimental associated vaccine against sheep pox, goat pox and small ruminant plague. In Proceedings of the Federal Center for Animal Health; The Federal Center for Animal Health: Vladimir, Rusia, 2008; Volume 6, pp. 266–272. [Google Scholar]
- Konstantinov, A.V.; Starov, S.K.; Diev, V.I.; Moroz, N.V.; Kurnenkova, E.V.; Basova, D.K.; Kononov, A.V.; Fedoseyev, K.Y. Antigenic and protective properties of experimental associated virus vaccine against sheep pox and goat pox. Veterinariâ. Segodnâ. 2018, 2, 42–46. [Google Scholar] [CrossRef]
- Legnardi, M.; Raizman, E.; Beltran-Alcrudo, D.; Cinardi, G.; Robinson, T.; Falzon, L.C.; Djomgang, H.K.; Okori, E.; Parida, S.; Njeumi, F.; et al. Peste des petits ruminants in central and Eastern Asia/West Eurasia: Epidemiological situation and status of control and eradication activities after the first phase of the PPR global eradication programme (2017–2021). Animals 2022, 12, 2030. [Google Scholar] [CrossRef]
- Wasee Ullah, R.; Bin Zahur, A.; Latif, A.; Iqbal Dasti, J.; Irshad, H.; Afzal, M.; Rasheed, T.; Rashid Malik, A.; Qureshi, Z.U. Detection of peste des petits ruminants viral RNA in fecal samples of goats after an outbreak in Punjab Province of Pakistan: A longitudinal study. BioMed Res. Int. 2016, 2016, 1486824. [Google Scholar] [CrossRef] [PubMed]
- Ezeibe, M.C.O.; Okoroafor, O.N.; Ngene, A.A.; Eze, J.I.; Eze, I.C.; Ugonabo, J.A.C. Persistent detection of peste de petits ruminants antigen in the faeces of recovered goats. Trop. Anim. Health Prod. 2008, 40, 517–519. [Google Scholar] [CrossRef] [PubMed]
- Bataille, A.; Kwiatek, O.; Belfkhi, S.; Mounier, L.; Parida, S.; Mahapatra, M.; Caron, A.; Chubwa, C.C.; Keyyu, J.; Kock, R.; et al. Optimization and evaluation of a non-invasive tool for peste des petits ruminants surveillance and control. Sci. Rep. 2019, 9, 4742. [Google Scholar] [CrossRef] [PubMed]
- Minor, P.D. Live attenuated vaccines: Historical successes and current challenges. Virology 2015, 479–480, 379–392. [Google Scholar] [CrossRef]
- Hanley, K.A. The double-edged sword: How evolution can make or break a live-attenuated virus vaccine. Evolution 2011, 4, 635–643. [Google Scholar] [CrossRef]
- Bull, J.J.; Smithson, M.W.; Nuismer, S.L. Transmissible viral vaccines. Trends Microbiol. 2018, 26, 6–15. [Google Scholar] [CrossRef]
- Eloiflin, R.J.; Boyer, M.; Kwiatek, O.; Guendouz, S.; Loire, E.; Servan de Almeida, R.; Libeau, G.; Bataille, A. Evolution of attenuation and risk of reversal in peste des petits ruminants vaccine strain Nigeria 75/1. Viruses 2019, 11, 724. [Google Scholar] [CrossRef]
- Mahapatra, M.; Selvaraj, M.; Parida, S. Comparison of immunogenicity and protective efficacy of PPR live attenuated vaccines (Nigeria 75/1 and Sungri 96) administered by intranasal and subcutaneous routes. Vaccines 2020, 8, 168. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symptom | Score | |
|---|---|---|
| Body temperature | >40 °C (every 24 h) | 0.5 |
| >41 °C (every 24 h) | 1 | |
| Vaccine injection site | Swelling over 3 cm in diameter; redness and pain for 12 h | 1 |
| Pathological changes in the skin or underneath | 2 | |
| Overall well-being | Apathy, weakness, and loss of appetite (every 24 h) | 0.5 |
| Reluctance to get up (every 24 h) | 2 | |
| Death | 6 | |
| Changes in the mucus epithelium of the mouth | Hyperemia | 1 |
| Erosive lesions; hypersalivation | 2 | |
| Necrosis | 3 | |
| Changes in the mucus epithelium of the nose | Hyperemia; nasal discharge | 1 |
| Erosive lesions | 2 | |
| Changes in the conjunctiva (conjunctivitis and serous discharge) | 2 | |
| Enlargement of lymph nodes | 1 | |
| Gross pathology in the respiratory tract | 3 | |
| Gross pathology in the gastrointestinal tract | 3 | |
| Group | Control | V1 | V2 | V3 |
|---|---|---|---|---|
| Average score after vaccination | 0 | 1.0 | 0.9 | 1.1 |
| Average score after challenge | 15.2 | 0.1 | 0.2 | 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byadovskaya, O.; Shalina, K.; Prutnikov, P.; Shumilova, I.; Tenitilov, N.; Konstantinov, A.; Moroz, N.; Chvala, I.; Sprygin, A. The Live Attenuated Vaccine Strain “ARRIAH” Completely Protects Goats from a Virulent Lineage IV Field Strain of Peste Des Petits Ruminants Virus. Vaccines 2024, 12, 110. https://doi.org/10.3390/vaccines12020110
Byadovskaya O, Shalina K, Prutnikov P, Shumilova I, Tenitilov N, Konstantinov A, Moroz N, Chvala I, Sprygin A. The Live Attenuated Vaccine Strain “ARRIAH” Completely Protects Goats from a Virulent Lineage IV Field Strain of Peste Des Petits Ruminants Virus. Vaccines. 2024; 12(2):110. https://doi.org/10.3390/vaccines12020110
Chicago/Turabian StyleByadovskaya, Olga, Kseniya Shalina, Pavel Prutnikov, Irina Shumilova, Nikita Tenitilov, Alexei Konstantinov, Nataliya Moroz, Ilya Chvala, and Alexander Sprygin. 2024. "The Live Attenuated Vaccine Strain “ARRIAH” Completely Protects Goats from a Virulent Lineage IV Field Strain of Peste Des Petits Ruminants Virus" Vaccines 12, no. 2: 110. https://doi.org/10.3390/vaccines12020110