
Effects of Influenza Vaccine on the Immune Responses to SARS-CoV-2 Vaccination
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cohort Description and Sampling
2.2. Ethics Statement
2.3. Serum Samples
2.4. Hemagglutination Inhibition Assay
2.5. Influenza Virus Microneutralization Assay
2.6. SARS-CoV-2 IgG Immunoassay Testing
2.7. SARS-CoV-2 Virus Neutralization Assay
2.8. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
3.2. Subjects Who Received Influenza and COVID-19 Vaccinations Are More Responsive to the SARS-CoV-2 Vaccine
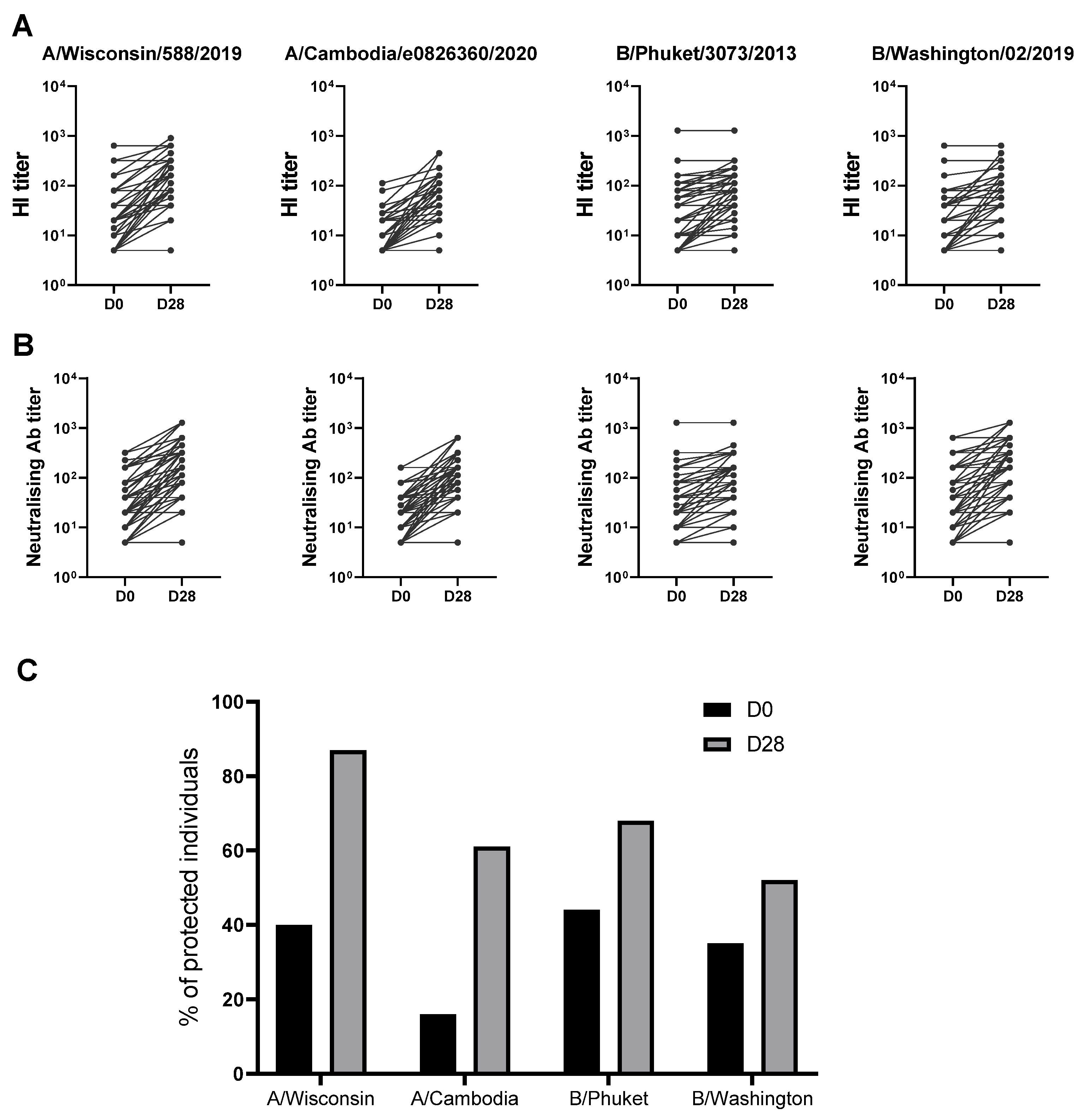
3.3. Quantitative and Qualitative Analysis of the Immune Responses to Seasonal 2021/2022 Influenza Vaccine
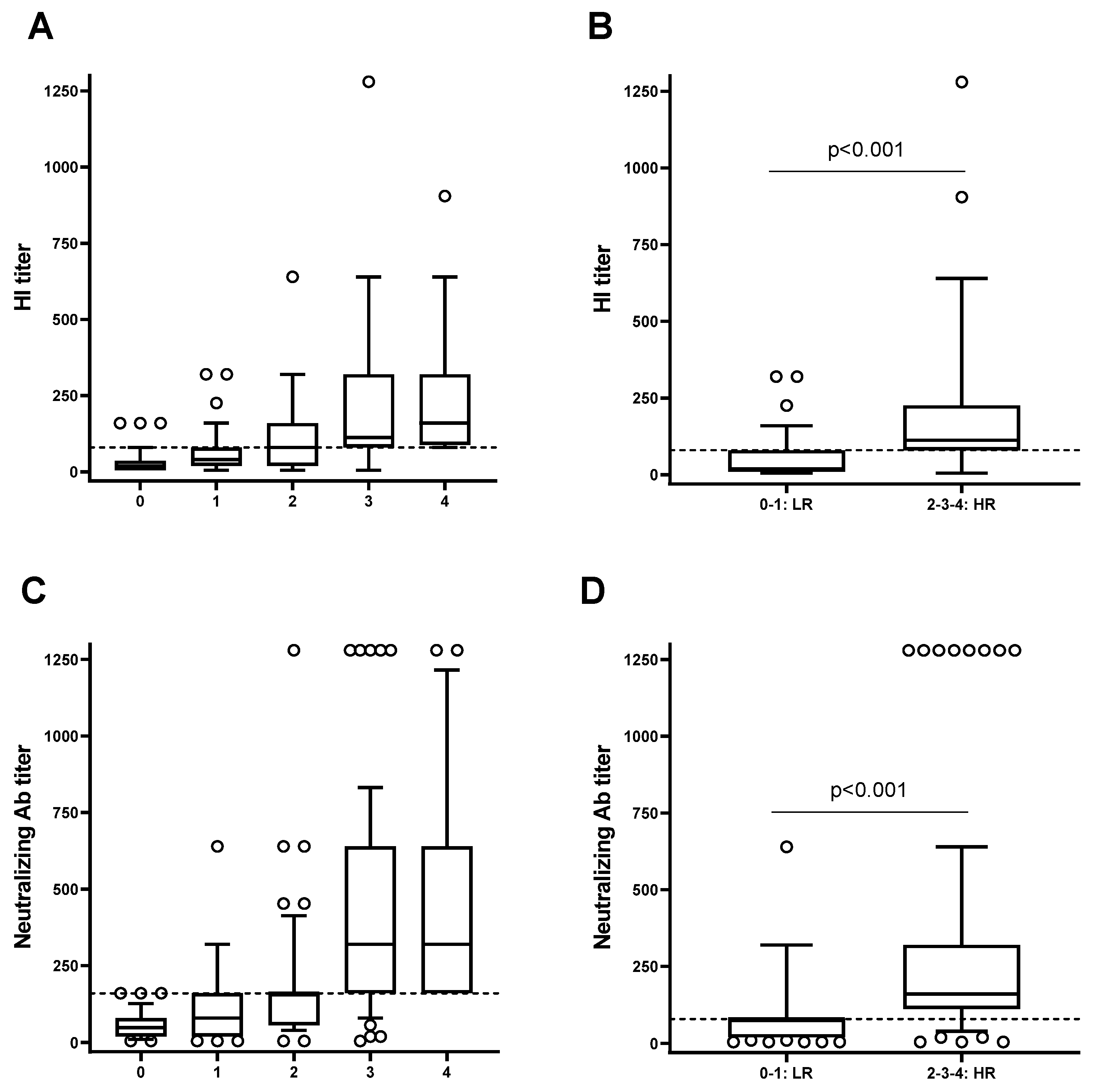
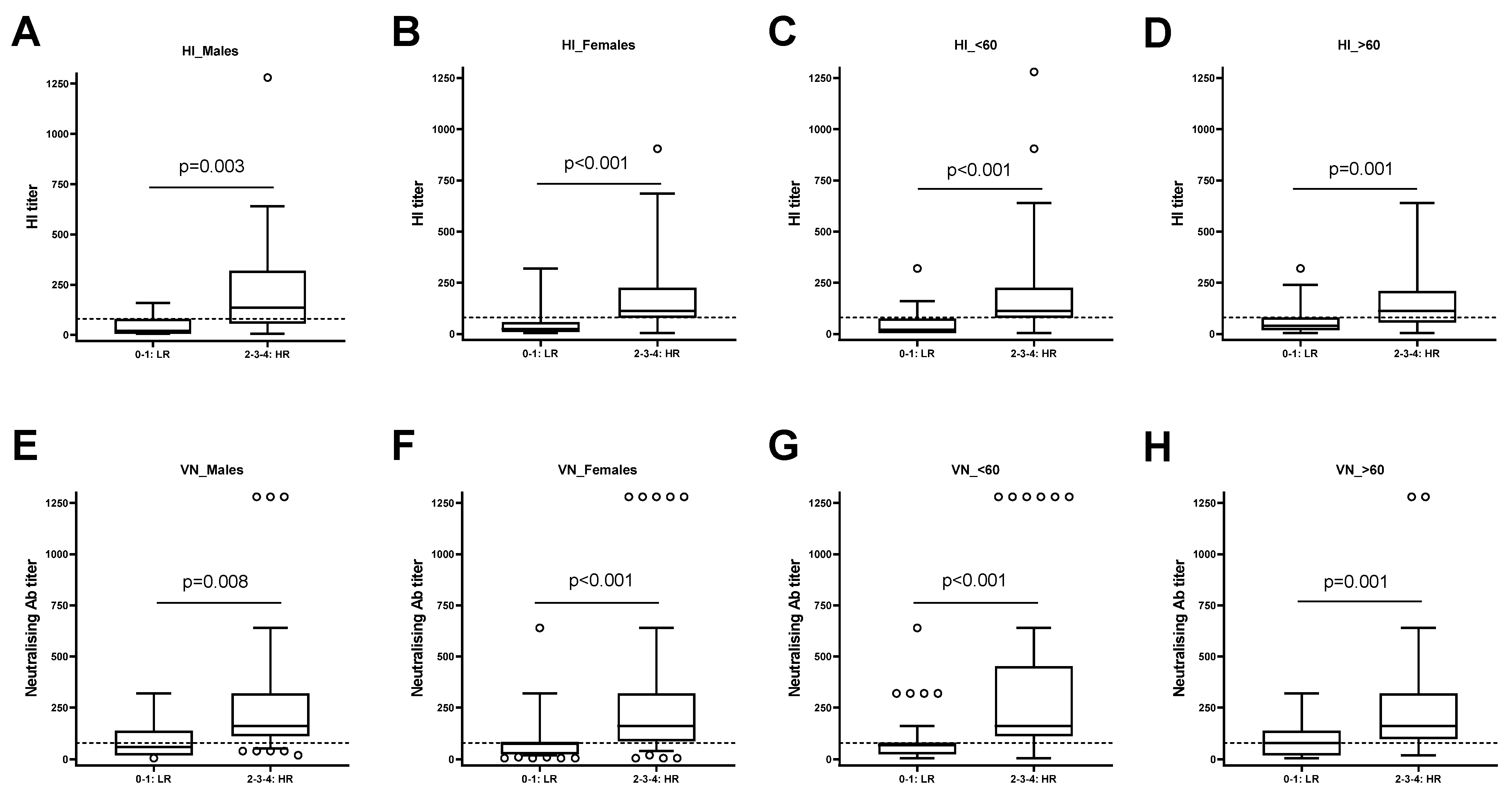
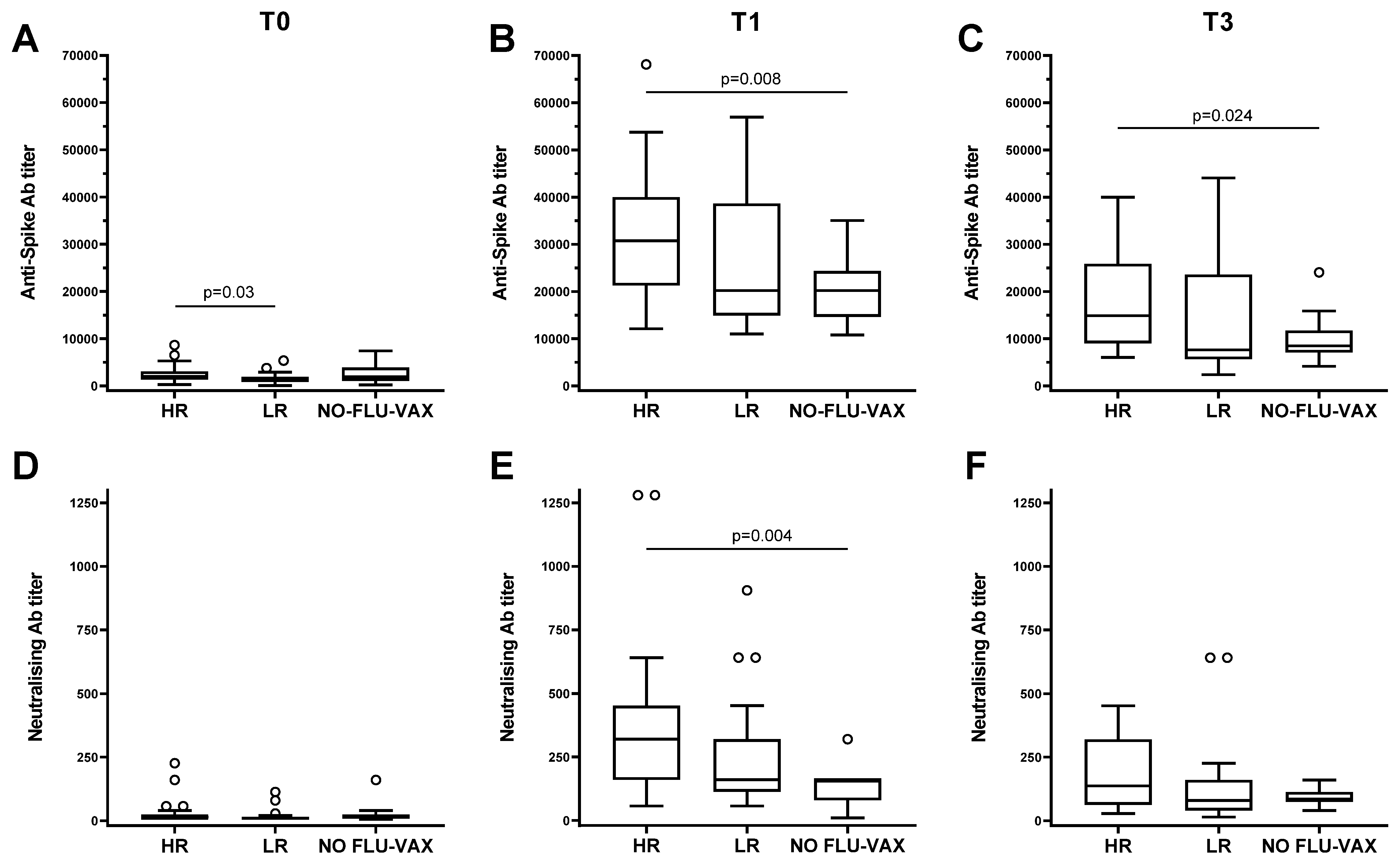
3.4. The High Responders to Influenza Vaccine Compared to the Low Responders Show a Strong Capacity to Respond to the SARS-CoV-2 Vaccine
3.5. Analysis of the Effects of the Flu-Specific Antibody Response on Anti-Spike Antibody Levels in Double-Vaccinated Individuals
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plotkin, S. History of Vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 12283–12287. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Floyd, K.; Wu, S.; Fang, Z.; Tan, T.K.; Froggatt, H.M.; Powers, J.M.; Leist, S.R.; Gully, K.L.; Hubbard, M.L.; et al. BCG Vaccination Stimulates Integrated Organ Immunity by Feedback of the Adaptive Immune Response to Imprint Prolonged Innate Antiviral Resistance. Nat. Immunol. 2023, 25, 41–53. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Ahmed, S.S.; Curtis, N.; Kollmann, T.R.; Levy, O.; Netea, M.G.; Pollard, A.J.; Van Crevel, R.; Wilson, C.B. Harnessing the Beneficial Heterologous Effects of Vaccination. Nat. Rev. Immunol. 2016, 16, 392–400. [Google Scholar] [CrossRef]
- Nielsen, S.; Sujan, H.M.; Benn, C.S.; Aaby, P.; Hanifi, S.M.A. Oral Polio Vaccine Campaigns May Reduce the Risk of Death from Respiratory Infections. Vaccines 2021, 9, 1133. [Google Scholar] [CrossRef]
- Paolo Matteo, A.; Serena, M.; Claudia Maria, T.; Emma, A. Flu Vaccine Administration in the Period before SARS-CoV-2 Infection and Its Outcomes: An Umbrella Review. Prev. Med. Rep. 2023, 38, 102575. [Google Scholar] [CrossRef]
- Fink, G.; Orlova-Fink, N.; Schindler, T.; Grisi, S.; Ferrer, A.P.S.; Daubenberger, C.; Brentani, A. Inactivated Trivalent Influenza Vaccination Is Associated with Lower Mortality among Patients with COVID-19 in Brazil. BMJ Evid. Based Med. 2021, 26, 192–193. [Google Scholar] [CrossRef]
- Candelli, M.; Pignataro, G.; Torelli, E.; Gullì, A.; Nista, E.C.; Petrucci, M.; Saviano, A.; Marchesini, D.; Covino, M.; Ojetti, V.; et al. Effect of Influenza Vaccine on COVID-19 Mortality: A Retrospective Study. Intern. Emerg. Med. 2021, 16, 1849–1855. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Liu, M.; Liu, J. The Association between Influenza Vaccination and COVID-19 and Its Outcomes: A Systematic Review and Meta-Analysis of Observational Studies. Vaccines 2021, 9, 529. [Google Scholar] [CrossRef]
- Martinez-Huedo, M.A.; Lopez-De-Andrés, A.; Mora-Zamorano, E.; Hernández-Barrera, V.; Jiménez-Trujillo, I.; Zamorano-Leon, J.J.; Jiménez-García, R. Decreasing Influenza Vaccine Coverage among Adults with High-Risk Chronic Diseases in Spain from 2014 to 2017. Hum. Vaccines Immunother. 2020, 16, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Lewnard, J.A.; Cobey, S. Immune History and Influenza Vaccine Effectiveness. Vaccines 2018, 6, 28. [Google Scholar] [CrossRef]
- Avey, S.; Cheung, F.; Fermin, D.; Frelinger, J.; Gaujoux, R.; Gottardo, R.; Khatri, P.; Kleinstein, S.H.; Kotliarov, Y.; Meng, H.; et al. Multicohort Analysis Reveals Baseline Transcriptional Predictors of Influenza Vaccination Responses. Sci. Immunol. 2017, 2, eaal4656. [Google Scholar] [CrossRef]
- Tsang, J.S.; Schwartzberg, P.L.; Kotliarov, Y.; Biancotto, A.; Xie, Z.; Germain, R.N.; Wang, E.; Olnes, M.J.; Narayanan, M.; Golding, H.; et al. Global Analyses of Human Immune Variation Reveal Baseline Predictors of Postvaccination Responses. Cell 2014, 157, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Hejblum, B.P.; Simon, N.; Jojic, V.; Dekker, C.L.; Thiébaut, R.; Tibshirani, R.J.; Davis, M.M. Systems Analysis of Sex Differences Reveals an Immunosuppressive Role for Testosterone in the Response to Influenza Vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Belongia, E.A.; Skowronski, D.M.; McLean, H.Q.; Chambers, C.; Sundaram, M.E.; De Serres, G. Repeated Annual Influenza Vaccination and Vaccine Effectiveness: Review of Evidence. Expert Rev. Vaccines 2017, 16, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Nienen, M.; Stervbo, U.; Mölder, F.; Kaliszczyk, S.; Kuchenbecker, L.; Gayova, L.; Schweiger, B.; Jürchott, K.; Hecht, J.; Neumann, A.U.; et al. The Role of Pre-Existing Cross-Reactive Central Memory CD4 T-Cells in Vaccination with Previously Unseen Influenza Strains. Front. Immunol. 2019, 10, 593. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Recommended Composition of Influenza Virus Vaccines for Use in the 2021–2022 Northern Hemisphere Influenza Season. Available online: https://www.who.int/publications/m/item/recommended-composition-of-influenza-virus-vaccines-for-use-in-the-2021-2022-northern-hemisphere-influenza-season (accessed on 26 January 2024).
- Manenti, A.; Maggetti, M.; Casa, E.; Martinuzzi, D.; Torelli, A.; Trombetta, C.M.; Marchi, S.; Montomoli, E. Evaluation of SARS-CoV-2 Neutralizing Antibodies Using a CPE-Based Colorimetric Live Virus Micro-Neutralization Assay in Human Serum Samples. J. Med. Virol. 2020, 92, 2096–2104. [Google Scholar] [CrossRef] [PubMed]
- Tartof, S.Y.; Slezak, J.M.; Fischer, H.; Hong, V.; Ackerson, B.K.; Ranasinghe, O.N.; Frankland, T.B.; Ogun, O.A.; Zamparo, J.M.; Gray, S.; et al. Effectiveness of MRNA BNT162b2 COVID-19 Vaccine up to 6 Months in a Large Integrated Health System in the USA: A Retrospective Cohort Study. Lancet 2021, 398, 1407–1416. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control Seasonal Influenza—Annual Epidemiological Report for 2021–2022. Available online: https://www.ecdc.europa.eu/en/publications-data/seasonal-influenza-annual-epidemiological-report-2021-2022 (accessed on 21 February 2023).
- Wimmers, F.; Donato, M.; Kuo, A.; Ashuach, T.; Gupta, S.; Li, C.; Dvorak, M.; Foecke, M.H.; Chang, S.E.; Hagan, T.; et al. The Single-Cell Epigenomic and Transcriptional Landscape of Immunity to Influenza Vaccination. Cell 2021, 184, 3915–3935.e21. [Google Scholar] [CrossRef]
- Geckin, B.; Konstantin Föhse, F.; Domínguez-Andrés, J.; Netea, M.G. Trained Immunity: Implications for Vaccination. Curr. Opin. Immunol. 2022, 77, 102190. [Google Scholar] [CrossRef]
- Rivas, M.N.; Ebinger, J.E.; Wu, M.; Sun, N.; Braun, J.; Sobhani, K.; van Eyk, J.E.; Cheng, S.; Arditi, M. BCG Vaccination History Associates with Decreased SARS-CoV-2 Seroprevalence across a Diverse Cohort of Health Care Workers. J. Clin. Investig. 2021, 131, e145157. [Google Scholar] [CrossRef]
- Ten Doesschate, T.; Moorlag, S.J.C.F.M.; Van Der Vaart, T.W.; Taks, E.; Debisarun, P.; Ten Oever, J.; Bleeker-Rovers, C.P.; Verhagen, P.B.; Lalmohamed, A.; Ter Heine, R.; et al. Two Randomized Controlled Trials of Bacillus Calmette-Guérin Vaccination to Reduce Absenteeism among Health Care Workers and Hospital Admission by Elderly Persons during the COVID-19 Pandemic: A Structured Summary of the Study Protocols for Two Randomised Controlled Trials. Trials 2020, 21, 481. [Google Scholar] [CrossRef] [PubMed]
- Ragni, P.; Marino, M.; Formisano, D.; Bisaccia, E.; Scaltriti, S.; Bedeschi, E.; Grilli, R. Association between Exposure to Influenza Vaccination and COVID-19 Diagnosis and Outcomes. Vaccines 2020, 8, 675. [Google Scholar] [CrossRef]
- Yang, M.J.; Rooks, B.J.; Le, T.T.T.; Santiago, I.O.; Diamond, J.; Dorsey, N.L.; Mainous, A.G. Influenza Vaccination and Hospitalizations among COVID-19 Infected Adults. J. Am. Board. Fam. Med. 2021, 34, S179–S182. [Google Scholar] [CrossRef]
- Wilcox, C.R.; Islam, N.; Dambha-Miller, H. Association between Influenza Vaccination and Hospitalisation or All-Cause Mortality in People with COVID-19: A Retrospective Cohort Study. BMJ Open Respir. Res. 2021, 8, e000857. [Google Scholar] [CrossRef]
- Visalli, G.; Laganà, A.; Lo Giudice, D.; Calimeri, S.; Caccamo, D.; Trainito, A.; Di Pietro, A.; Facciolà, A. Towards a Future of Personalized Vaccinology: Study on Individual Variables Influencing the Antibody Response to the COVID-19 Vaccine. Vaccines 2023, 11, 217. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. Factors That Influence the Immune Response to Vaccination. Clin. Microbiol. Rev. 2019, 32, e00084-18. [Google Scholar] [CrossRef]
- Meireles, L.C.; Marinho, R.T.; Van Damme, P. Three Decades of Hepatitis B Control with Vaccination. World J. Hepatol. 2015, 7, 2127–2132. [Google Scholar] [CrossRef] [PubMed]
- Camous, X.; Visan, L.; Ying, C.T.T.; Abel, B.; Nyunt, M.S.Z.; Narang, V.; Poidinger, M.; Carre, C.; Sesay, S.; Bosco, N.; et al. Healthy Elderly Singaporeans Show No Age-Related Humoral Hyporesponsiveness nor Diminished Plasmablast Generation in Response to Influenza Vaccine. Immun. Ageing 2018, 15, 28. [Google Scholar] [CrossRef]
- Carre, C.; Wong, G.; Narang, V.; Tan, C.; Chong, J.; Chin, H.X.; Xu, W.; Lu, Y.; Chua, M.; Poidinger, M.; et al. Endoplasmic Reticulum Stress Response and Bile Acid Signatures Associate with Multi-Strain Seroresponsiveness during Elderly Influenza Vaccination. iScience 2021, 24, 102970. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Kistner, O.; Montomoli, E.; Viviani, S.; Marchi, S. Influenza Viruses and Vaccines: The Role of Vaccine Effectiveness Studies for Evaluation of the Benefits of Influenza Vaccines. Vaccines 2022, 10, 714. [Google Scholar] [CrossRef]
- Waldock, J.; Remarque, E.J.; Zheng, L.; Ho, S.; Hoschler, K.; Neumann, B.; Sediri-Schön, H.; Trombetta, C.M.; Montomoli, E.; Marchi, S.; et al. Haemagglutination Inhibition and Virus Microneutralisation Serology Assays: Use of Harmonised Protocols and Biological Standards in Seasonal Influenza Serology Testing and Their Impact on Inter-Laboratory Variation and Assay Correlation: A FLUCOP Collaborative Study. Front. Immunol. 2023, 14, 1155552. [Google Scholar] [PubMed]
- Ng, T.; Flores-Malavet, V.; Mansoor, M.A.M.; Arvelo, A.C.; Dhume, K.; Prokop, E.; McKinstry, K.K.; Strutt, T.M. Intermediate Levels of Pre-Existing Protective Antibody Allow Priming of Protective T Cell Immunity against Influenza. J. Immunol. 2023, 210, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Kavian, N.; Hachim, A.; Cowling, B.J.; Valkenburg, S.A. Repeated Influenza Vaccination Provides Cumulative Protection from Distinct H3N2 Viruses. Clin. Transl. Immunol. 2021, 10, e1297. [Google Scholar] [CrossRef]
- Cevirgel, A.; Shetty, S.A.; Vos, M.; Nanlohy, N.M.; Beckers, L.; Bijvank, E.; Rots, N.; van Beek, J.; Buisman, A.; van Baarle, D. Pre-vaccination Immunotypes Reveal Weak and Robust Antibody Responders to Influenza Vaccination. Aging Cell 2023, 23, e14048. [Google Scholar] [CrossRef]
- Künzel, W.; Glathe, H.; Engelmann, H.; Van Hoecke, C. Kinetics of to Trivalent Humoral Antibody Response Inactivated Split. Influenza Vaccine in Subjects Previously Vaccinated or Vaccinated for the First Time. Vaccine 1996, 14, 1108–1110. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Flannery, B.; Foppa, I.M.; Chung, J.R.; Nowalk, M.P.; Zimmerman, R.K.; Gaglani, M.; Monto, A.S.; Martin, E.T.; Belongia, E.A.; et al. Effects of Prior Season Vaccination on Current Season Vaccine Effectiveness in the United States Flu Vaccine Effectiveness Network, 2012–2013 Through 2017–2018. Clin. Infect. Dis. 2021, 73, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Sugishita, Y.; Nakayama, T.; Sugawara, T.; Ohkusa, Y. Negative Effect on Immune Response of Repeated Influenza Vaccination and Waning Effectiveness in Interseason for Elderly People. Vaccine 2020, 38, 3759–3765. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Fukushima, W.; Morikawa, S.; Fujioka, M.; Matsushita, T.; Kubota, M.; Yagi, Y.; Takasaki, Y.; Shindo, S.; Yamashita, Y.; et al. Influence of Prior Influenza Vaccination on Current Influenza Vaccine Effectiveness in Children Aged 1 to 5 Years. Vaccines 2021, 9, 1447. [Google Scholar] [CrossRef]
- Kitamura, S.; Matsushita, M.; Komatsu, N.; Yagi, Y.; Takeuchi, S.; Seo, H. Impact of Repeated Yearly Vaccination on Immune Responses to Influenza Vaccine in an Elderly Population. Am. J. Infect. Control 2020, 48, 1422–1425. [Google Scholar] [CrossRef]







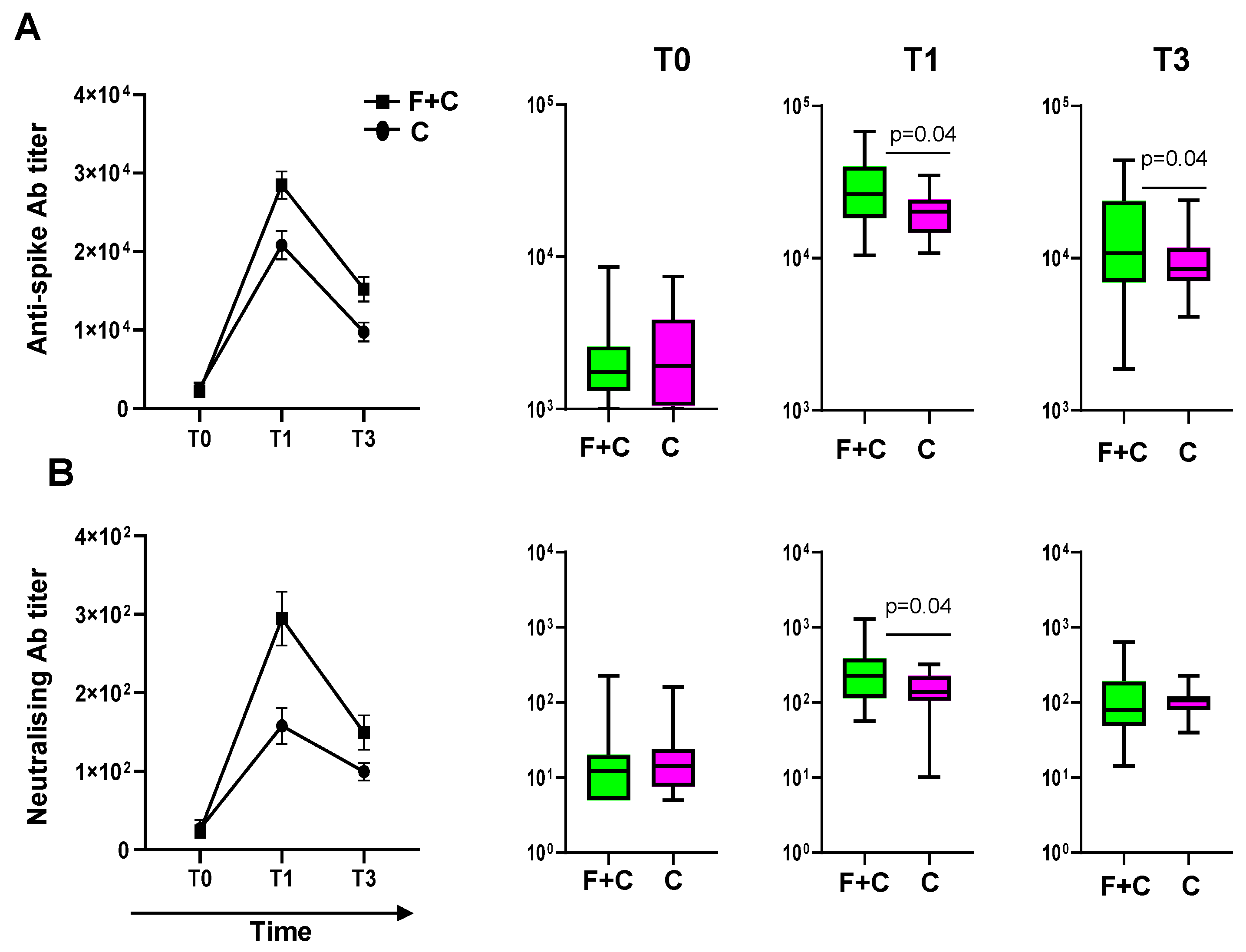{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Two Vaccines (n = 54, 73%) | One Vaccine (n = 20, 27%) | Total (n = 74) | ||||
|---|---|---|---|---|---|---|
| Gender | n. | % | n. | % | n. | % |
| Male | 18 | 33 | 6 | 30 | 24 | 32 |
| Female | 36 | 67 | 14 | 70 | 50 | 68 |
| Age | ||||||
| Median | 57 | 51 | 54 | |||
| IQR; range | (49–60); (35–70) | (49–56); (31–72) | (49–60); (31–72) | |||
| Education | ||||||
| Higher | 41 | 76 | 12 | 60 | 53 | 72 |
| Middle or lower | 13 | 24 | 8 | 40 | 21 | 28 |
| Frailty | ||||||
| Very fit | 8 | 15 | 7 | 35 | 15 | 20 |
| Fit | 39 | 72 | 12 | 60 | 51 | 69 |
| Managing well | 7 | 13 | 1 | 5 | 8 | 11 |
| Body mass index | ||||||
| Underweight | 1 | 2 | 0 | 0 | 1 | 1 |
| Healthy weight | 30 | 55 | 12 | 60 | 42 | 57 |
| Overweight | 20 | 37 | 6 | 30 | 26 | 35 |
| Obesity | 3 | 6 | 2 | 10 | 5 | 7 |
| Smoking | ||||||
| Yes | 7 | 13 | 5 | 25 | 12 | 16 |
| No | 47 | 87 | 15 | 75 | 62 | 84 |
| Medical conditions | ||||||
| Yes | 23 | 43 | 11 | 55 | 34 | 46 |
| No | 31 | 57 | 9 | 45 | 40 | 54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riccomi, A.; Trombetta, C.M.; Dorrucci, M.; Di Placido, D.; Sanarico, N.; Farchi, F.; Giuseppetti, R.; Villano, U.; Marcantonio, C.; Marchi, S.; et al. Effects of Influenza Vaccine on the Immune Responses to SARS-CoV-2 Vaccination. Vaccines 2024, 12, 425. https://doi.org/10.3390/vaccines12040425
Riccomi A, Trombetta CM, Dorrucci M, Di Placido D, Sanarico N, Farchi F, Giuseppetti R, Villano U, Marcantonio C, Marchi S, et al. Effects of Influenza Vaccine on the Immune Responses to SARS-CoV-2 Vaccination. Vaccines. 2024; 12(4):425. https://doi.org/10.3390/vaccines12040425
Chicago/Turabian StyleRiccomi, A., C. M. Trombetta, M. Dorrucci, D. Di Placido, N. Sanarico, F. Farchi, R. Giuseppetti, U. Villano, C. Marcantonio, S. Marchi, and et al. 2024. "Effects of Influenza Vaccine on the Immune Responses to SARS-CoV-2 Vaccination" Vaccines 12, no. 4: 425. https://doi.org/10.3390/vaccines12040425