
Vaccination with a Protective Ipa Protein-Containing Nanoemulsion Differentially Alters the Transcriptomic Profiles of Young and Elderly Mice following Shigella Infection
 , , , ,
, , , ,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Protein Preparation
2.3. Preparation of L-DBF BECC438 or BECC470/ME Formulations
2.4. Mice and Immunizations
2.4.1. Young Mice
2.4.2. Elderly Mice
2.4.3. Mature Mice
2.5. IgG and IgA ELISAs
2.6. Shigella Challenge Studies
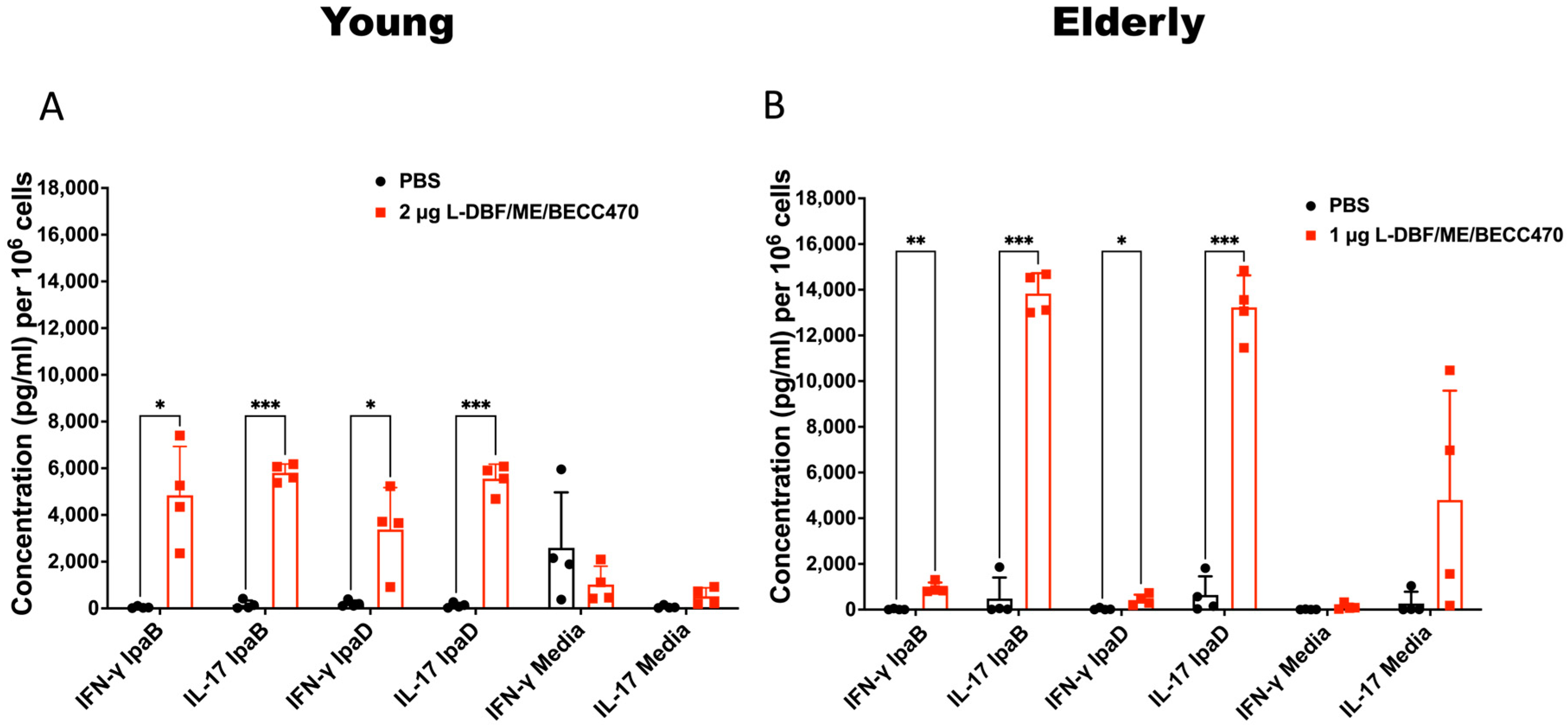
2.7. Cytokine Determinations
2.8. Bulk RNA Sequencing
2.9. RNA Seq Data Preprocessing and Normalization
2.10. Differential Expression Analysis
2.11. Heatmaps
2.12. Volcano Plots
2.13. Gene Ontology (GO) Enrichment Analysis
2.14. Principal Component Analysis (PCA)
2.15. Immune Pathway and Gene Cluster Analyses
2.16. Statistical Analyses
3. Results
3.1. Different Ages of Mice Immunized with Multimeric LDBF + BECC470 Formulated in an Oil-in-Water Emulsion Experience Protection against Shigella Infection
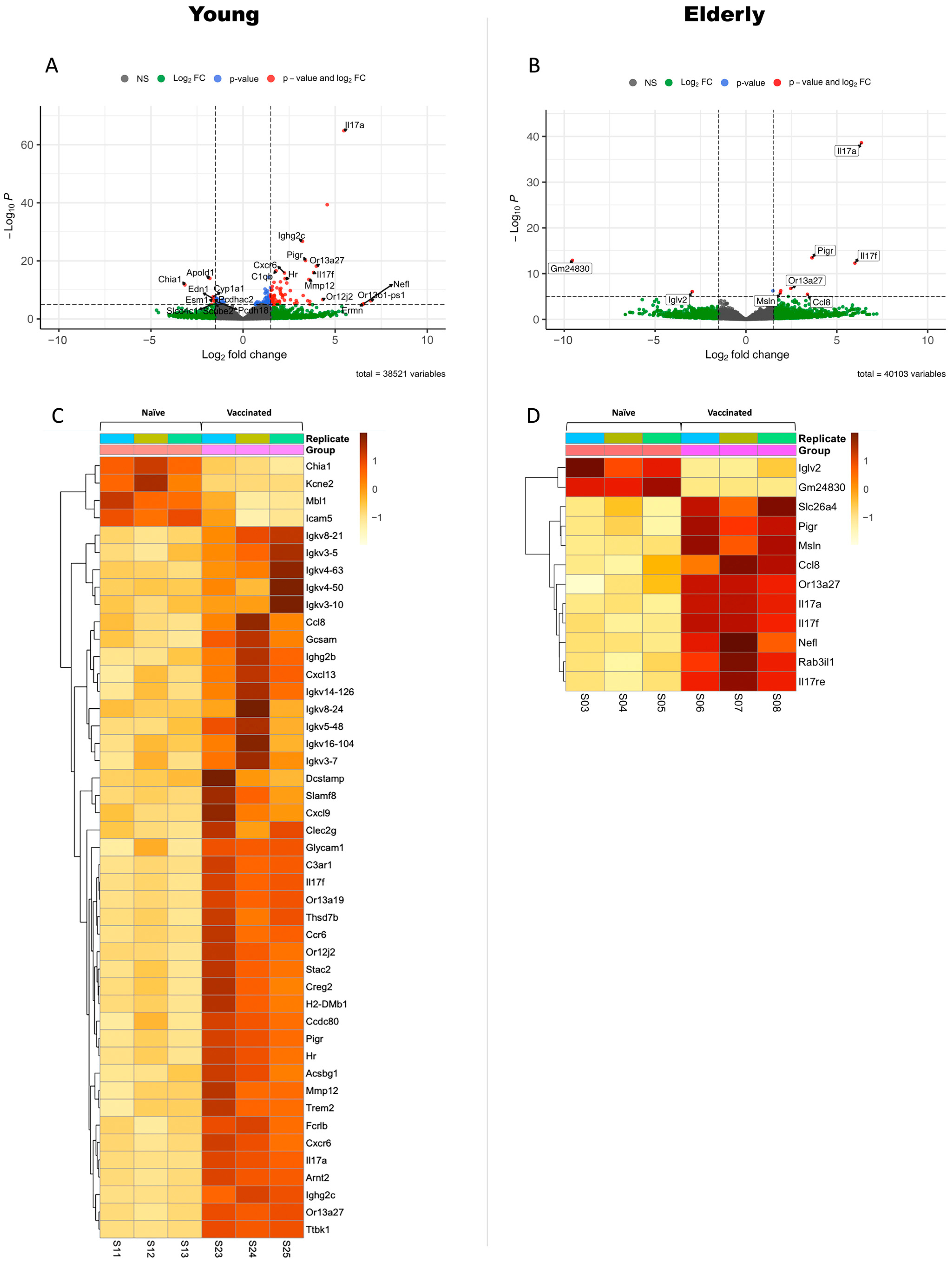
3.2. Differential Gene Expression in Vaccinated Young or Elderly Mice before Infection
3.3. Gene Expression Profiling between Naïve Infected Mice vs. Naïve Mice in Different Age Sets
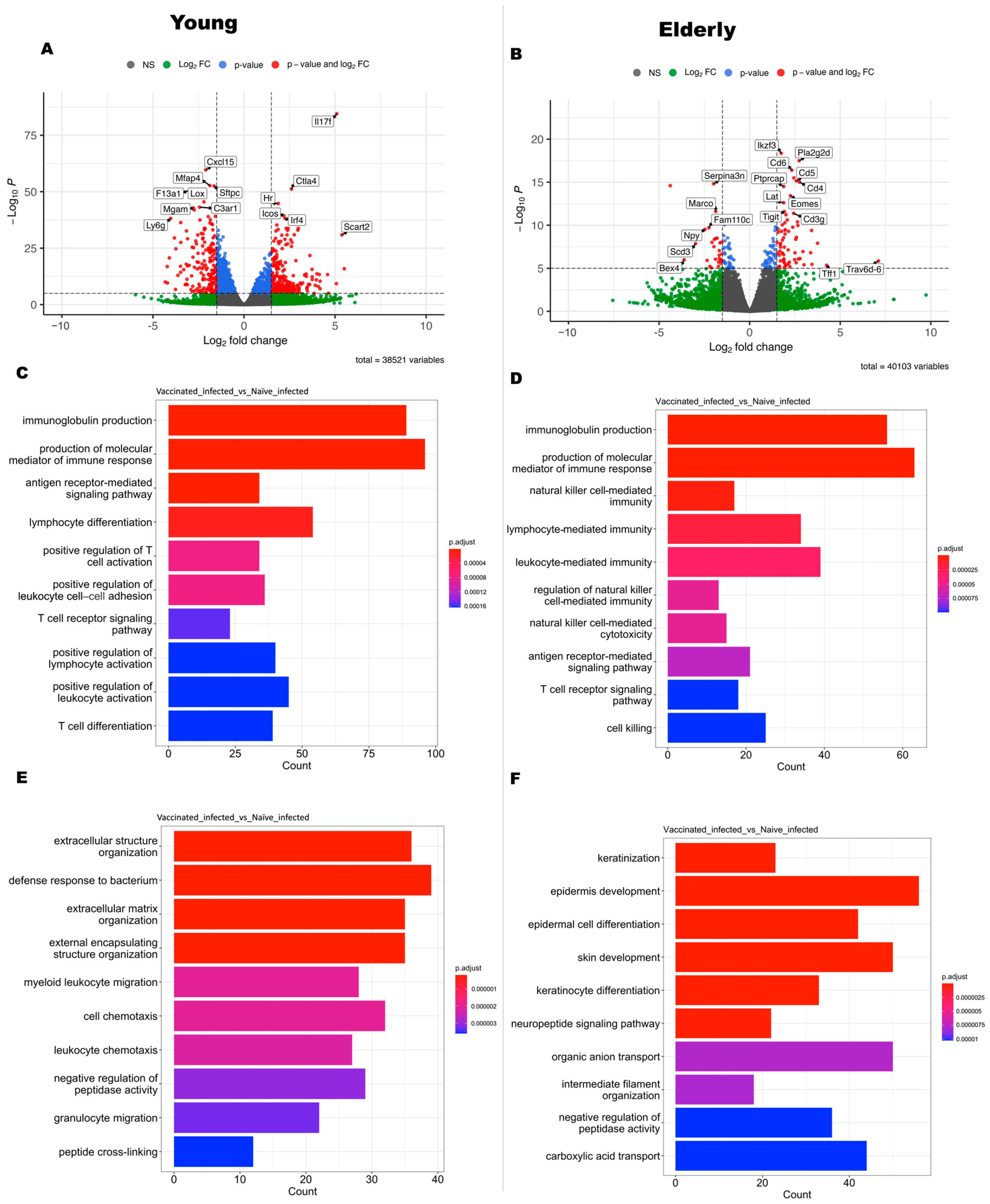
3.4. Gene Expression Profiling between Vaccinated Infected Mice vs. Vaccinated Mice in Different Age Sets
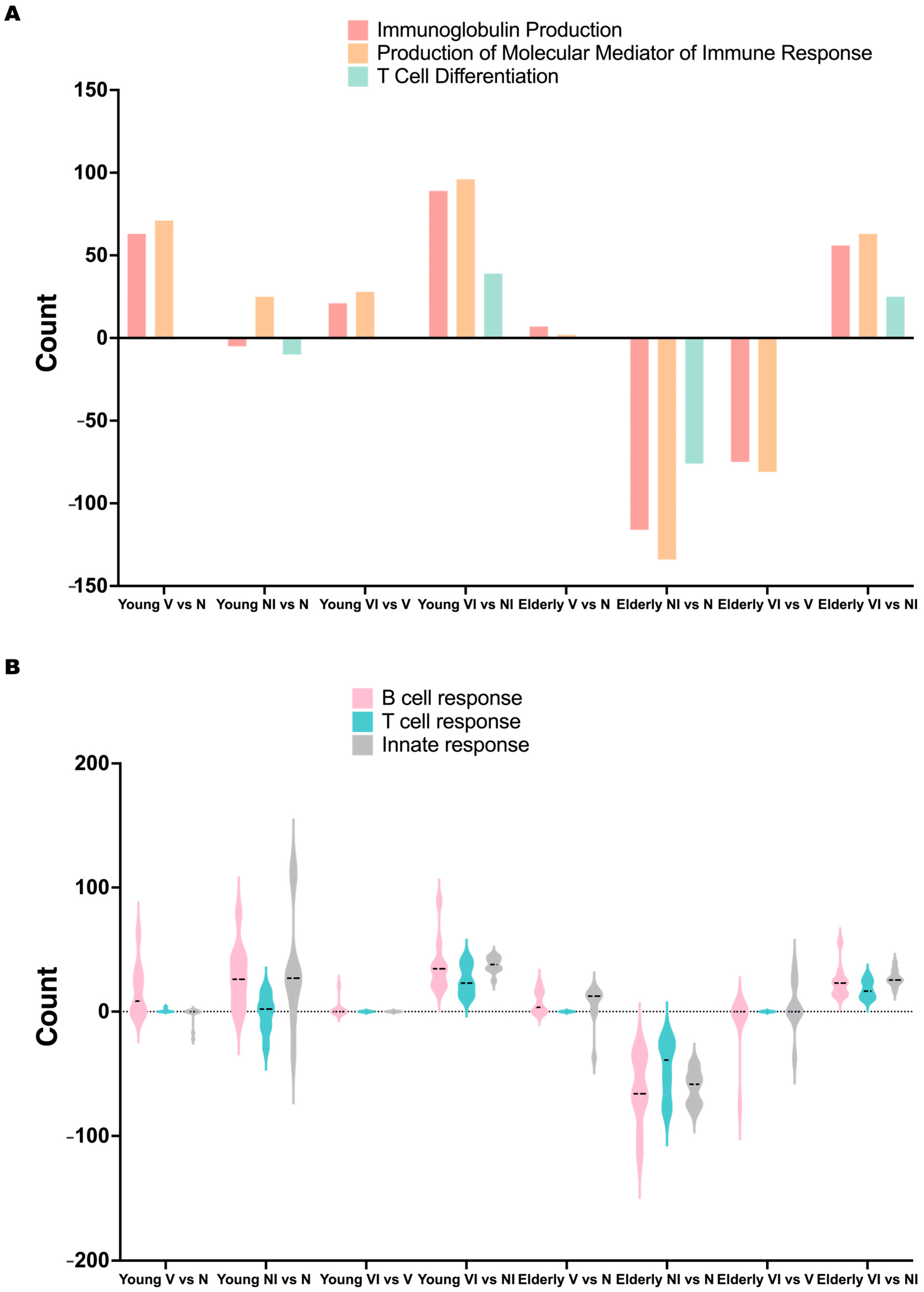
3.5. Age-Dependent Variations in Gene Expression and Pathway Alterations Related to Vaccine-Induced Immune Responses after Infection
3.6. Comparative Analysis of B Cell, T Cell, and Innate Responses in Young and Elderly Mice following Vaccination and Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, T.; Das, S.; Howlader, D.R.; Picking, W.D.; Picking, W.L. Shigella Vaccines: The Continuing Unmet Challenge. Int. J. Mol. Sci. 2024, 25, 4329. [Google Scholar] [CrossRef]
- Williams, P.C.M.; Berkley, J.A. Guidelines for the treatment of dysentery (shigellosis): A systematic review of the evidence. Paediatr. Int. Child Health 2018, 38 (Suppl. 1), S50–S65. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Arnold, S.L.M. Target Product Profile and Development Path for Shigellosis Treatment with Antibacterials. ACS Infect. Dis. 2021, 7, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Barry, E.M.; Pasetti, M.F.; Sztein, M.B.; Fasano, A.; Kotloff, K.L.; Levine, M.M. Progress and pitfalls in Shigella vaccine research. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.; Wierzba, T.; Walker, R.I. Status of vaccine research and development for Shigella. Vaccine 2016, 34, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Muthuramalingam, M.; Whittier, S.K.; Picking, W.L.; Picking, W.D. The Shigella Type III Secretion System: An Overview from Top to Bottom. Microorganisms 2021, 9, 451. [Google Scholar] [CrossRef]
- Espina, M.; Olive, A.J.; Kenjale, R.; Moore, D.S.; Ausar, S.F.; Kaminski, R.W.; Oaks, E.V.; Middaugh, C.R.; Picking, W.D.; Picking, W.L. IpaD Localizes to the Tip of the Type III Secretion System Needle of Shigella flexneri. Infect. Immun. 2006, 74, 4391–4400. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Becerra, F.J.; Kissmann, J.M.; Diaz-McNair, J.; Choudhari, S.P.; Quick, A.M.; Mellado-Sanchez, G.; Clements, J.D.; Pasetti, M.F.; Picking, W.L. Broadly protective Shigella vaccine based on type III secretion apparatus proteins. Infect. Immun. 2012, 80, 1222–1231. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lu, T.; Das, S.; Howlader, D.R.; Zheng, Q.; Siva Sai Kumar, R.; Whittier, S.K.; Picking, W.D.; Picking, W.L. L-DBF elicits cross protection against different serotypes of Shigella spp. Front. Trop. Dis. 2021, 2, 729731. [Google Scholar] [CrossRef]
- Lu, T.; Das, S.; Howlader, D.R.; Jain, A.; Hu, G.; Dietz, Z.K.; Zheng, Q.; Ratnakaram, S.S.K.; Whittier, S.K.; Varisco, D.J.; et al. Impact of the TLR4 agonist BECC438 on a novel vaccine formulation against Shigella spp. Front. Immunol. 2023, 14, 1194912. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mallett, C.P.; VanDeVerg, L.; Collins, H.H.; Hale, T.L. Evaluation of Shigella vaccine safety and efficacy in an intranasally challenged mouse model. Vaccine 1993, 11, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Roncaioli, J.L.; Turcotte, E.A.; Goers, L.; Chavez, R.A.; Lee, A.Y.; Lesser, C.F.; Rauch, I.; Vance, R.E. NAIP-NLRC4-deficient mice are susceptible to shigellosis. eLife 2020, 9, e59022. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Alphonse, N.; Odendall, C. Animal models of shigellosis: A historical overview. Curr. Opin. Immunol. 2023, 85, 102399. [Google Scholar] [CrossRef] [PubMed]
- Gharpure, R.; Marsh, Z.A.; Tack, D.M.; Collier, S.A.; Strysko, J.; Ray, L.; Payne, D.C.; Garcia-Williams, A.G. Disparities in Incidence and Severity of Shigella Infections among Children—Foodborne Diseases Active Surveillance Network (FoodNet), 2009–2018. J. Pediatr. Infect. Dis. Soc. 2021, 10, 782–788. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Chen, Q.; Wang, Y.; Chu, M.; Hu, Q.Q.; Hannah, M.N.; Rui, J.; Liu, X.C.; Yu, Y.H.; Zhao, F.W.; et al. Relative transmissibility of shigellosis among different age groups: A modeling study in Hubei Province, China. PLoS Negl. Trop. Dis. 2021, 15, e0009501. [Google Scholar] [CrossRef]
- Grinblat, J.; Weiss, A.; Grosman, B.; Dicker, D.; Beloosesky, Y. Diarrhea in elderly patients due to Clostridium difficile associated with Salmonella and Shigella infection. Arch. Gerontol. Geriatr. 2004, 39, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Ponnappan, S.; Ponnappan, U. Aging and immune function: Molecular mechanisms to interventions. Antioxid. Redox Signal. 2011, 14, 1551–1585. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Frasca, D.; Blomberg, B.B. Aging induces B cell defects and decreased antibody responses to influenza infection and vaccination. Immun. Ageing 2020, 17, 37. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Nikolich-Zugich, J. Ageing and life-long maintenance of T-cell subsets in the face of latent persistent infections. Nat. Rev. Immunol. 2008, 8, 512–522. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yu, Y.; Lu, C.; Yu, W.; Lei, Y.; Sun, S.; Liu, P.; Bai, F.; Chen, Y.; Chen, J. B Cells Dynamic in Aging and the Implications of Nutritional Regulation. Nutrients 2024, 16, 487. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee, K.A.; Flores, R.R.; Jang, I.H.; Saathoff, A.; Robbins, P.D. Immune Senescence, Immunosenescence and Aging. Front. Aging 2022, 3, 900028. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Haupt, R.E.; Harberts, E.M.; Kitz, R.J.; Strohmeier, S.; Krammer, F.; Ernst, R.K.; Frieman, M.B. Novel TLR4 adjuvant elicits protection against homologous and heterologous Influenza A infection. Vaccine 2021, 39, 5205–5213. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee, K.S.; Rader, N.A.; Miller-Stump, O.A.; Cooper, M.; Wong, T.Y.; Shahrier Amin, M.; Barbier, M.; Bevere, J.R.; Ernst, R.K.; Heath Damron, F. Intranasal VLP-RBD vaccine adjuvanted with BECC470 confers immunity against Delta SARS-CoV-2 challenge in K18-hACE2-mice. Vaccine 2023, 41, 5003–5017. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Martinez-Becerra, F.J.; Chen, X.; Dickenson, N.E.; Choudhari, S.P.; Harrison, K.; Clements, J.D.; Picking, W.D.; Van De Verg, L.L.; Walker, R.I.; Picking, W.L. Characterization of a novel fusion protein from IpaB and IpaD of Shigella spp. and its potential as a pan-Shigella vaccine. Infect. Immun. 2013, 81, 4470–4477. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Iyer, V.; Cayatte, C.; Guzman, B.; Schneider-Ohrum, K.; Matuszak, R.; Snell, A.; Rajani, G.M.; McCarthy, M.P.; Muralidhara, B. Impact of formulation and particle size on stability and immunogenicity of oil-in-water emulsion adjuvants. Hum. Vaccines Immunother. 2015, 11, 1853–1864. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Howlader, D.R.; Das, S.; Lu, T.; Hu, G.; Varisco, D.J.; Dietz, Z.K.; Walton, S.P.; Ratnakaram, S.S.K.; Gardner, F.M.; Ernst, R.K.; et al. Effect of Two Unique Nanoparticle Formulations on the Efficacy of a Broadly Protective Vaccine against Pseudomonas aeruginosa. Front. Pharmacol. 2021, 12, 706157. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- R Core Team. R: A Language and Environment for Statistical Computing; (Version: 4.3.0); R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 10 April 2023).
- Kolde, R. Pheatmap: Pretty Heatmaps. [R Package]; Version 1.0.12; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 10 April 2023).
- Blighe, K.; Rana, S.; Lewis, M. EnhancedVolcano: Publication-Ready Volcano Plots with Enhanced Colouring and Labeling [R Package]; Version 1.20.0; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://bioconductor.org/packages/EnhancedVolcano (accessed on 10 April 2023).
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tang, Y. Data Visualization Tools for Statistical Analysis Results [R Package]; Version 0.4.16; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://cran.r-project.org/web/packages/ggfortify/vignettes/plot_pca.html (accessed on 10 April 2023).
- Sellge, G.; Magalhaes, J.G.; Konradt, C.; Fritz, J.H.; Salgado-Pabon, W.; Eberl, G.; Bandeira, A.; Di Santo, J.P.; Sansonetti, P.J.; Phalipon, A. Th17 cells are the dominant T cell subtype primed by Shigella flexneri mediating protective immunity. J. Immunol. 2010, 184, 2076–2085. [Google Scholar] [CrossRef] [PubMed]
- Aslam, A.; Okafor, C.N. Shigella; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Frasca, D.; Blomberg, B.B. B Cell-Specific Biomarkers for Optimal Antibody Responses to Influenza Vaccination and Molecular Pathways That Reduce B Cell Function with Aging. Crit. Rev. Immunol. 2016, 36, 523–537. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hou, Y.; Chen, M.; Bian, Y.; Hu, Y.; Chuan, J.; Zhong, L.; Zhu, Y.; Tong, R. Insights into vaccines for elderly individuals: From the impacts of immunosenescence to delivery strategies. NPJ Vaccines 2024, 9, 77. [Google Scholar] [CrossRef]
- Song, X.; He, X.; Li, X.; Qian, Y. The roles and functional mechanisms of interleukin-17 family cytokines in mucosal immunity. Cell. Mol. Immunol. 2016, 13, 418–431. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hosangadi, D.; Smith, P.G.; Giersing, B.K. Considerations for using ETEC and Shigella disease burden estimates to guide vaccine development strategy. Vaccine 2019, 37, 7372–7380. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Institute of Medicine; Board on Global Health; Forum on Microbial Threats. Global Climate Change and Extreme Weather Events: Understanding the Contributions to Infectious Disease Emergence: Workshop Summary; The National Academies Collection: Reports Funded by National Institutes of Health; National Academies Press: Washington, DC, USA, 2008. [Google Scholar]
- Giersing, B.K.; Isbrucker, R.; Kaslow, D.C.; Cavaleri, M.; Baylor, N.; Maiga, D.; Pavlinac, P.B.; Riddle, M.S.; Kang, G.; MacLennan, C.A. Clinical and regulatory development strategies for Shigella vaccines intended for children younger than 5 years in low-income and middle-income countries. Lancet Glob. Health 2023, 11, e1819–e1826. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hoffmann, J.P.; Srivastava, A.; Yang, H.; Iwanaga, N.; Remcho, T.P.; Hewes, J.L.; Sharoff, R.; Song, K.; Norton, E.B.; Kolls, J.K.; et al. Vaccine-elicited IL-1R signaling results in Th17 TRM-mediated immunity. Commun. Biol. 2024, 7, 433. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Garnica, M.; Aiello, A.; Ligotti, M.E.; Accardi, G.; Arasanz, H.; Bocanegra, A.; Blanco, E.; Calabrò, A.; Chocarro, L.; Echaide, M.; et al. How Can We Improve the Vaccination Response in Older People? Part II: Targeting Immunosenescence of Adaptive Immunity Cells. Int. J. Mol. Sci. 2022, 23, 9797. [Google Scholar] [CrossRef]
- Dubois, V.; Chatagnon, J.; Thiriard, A.; Bauderlique-Le Roy, H.; Debrie, A.S.; Coutte, L.; Locht, C. Suppression of mucosal Th17 memory responses by acellular pertussis vaccines enhances nasal Bordetella pertussis carriage. NPJ Vaccines 2021, 6, 6. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Brinkhoff, A.; Sieberichs, A.; Engler, H.; Dolff, S.; Benson, S.; Korth, J.; Schedlowski, M.; Kribben, A.; Witzke, O.; Wilde, B. Pro-Inflammatory Th1 and Th17 Cells Are Suppressed During Human Experimental Endotoxemia Whereas Anti-Inflammatory IL-10 Producing T-Cells Are Unaffected. Front. Immunol. 2018, 9, 1133. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kak, G.; Raza, M.; Tiwari, B.K. Interferon-gamma (IFN-γ): Exploring its implications in infectious diseases. Biomol. Concepts 2018, 9, 64–79. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B. IFNgamma: Signalling, epigenetics and roles in immunity, metabolism, disease and cancer immunotherapy. Nat. Rev. Immunol. 2018, 18, 545–558. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, T.; Raju, M.; Howlader, D.R.; Dietz, Z.K.; Whittier, S.K.; Varisco, D.J.; Ernst, R.K.; Coghill, L.M.; Picking, W.D.; Picking, W.L. Vaccination with a Protective Ipa Protein-Containing Nanoemulsion Differentially Alters the Transcriptomic Profiles of Young and Elderly Mice following Shigella Infection. Vaccines 2024, 12, 618. https://doi.org/10.3390/vaccines12060618
Lu T, Raju M, Howlader DR, Dietz ZK, Whittier SK, Varisco DJ, Ernst RK, Coghill LM, Picking WD, Picking WL. Vaccination with a Protective Ipa Protein-Containing Nanoemulsion Differentially Alters the Transcriptomic Profiles of Young and Elderly Mice following Shigella Infection. Vaccines. 2024; 12(6):618. https://doi.org/10.3390/vaccines12060618
Chicago/Turabian StyleLu, Ti, Murugesan Raju, Debaki R. Howlader, Zackary K. Dietz, Sean K. Whittier, David J. Varisco, Robert K. Ernst, Lyndon M. Coghill, William D. Picking, and Wendy L. Picking. 2024. "Vaccination with a Protective Ipa Protein-Containing Nanoemulsion Differentially Alters the Transcriptomic Profiles of Young and Elderly Mice following Shigella Infection" Vaccines 12, no. 6: 618. https://doi.org/10.3390/vaccines12060618