
Trichinella spiralis Infection Inhibits the Efficacy of RBD Protein of SARS-CoV-2 Vaccination via Regulating Humoral and Cellular Immunity
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Parasites
2.2. Infection, Vaccination and Treatment
2.3. Enzyme-Linked Immunosorbent Assay (ELISA) of RBD-Specific Antibody Titers
2.4. ELISA for Cytokines
2.5. Detection of Antibody Neutralization Activity
2.6. Flow Cytometry
2.7. Statistical Analysis
3. Results
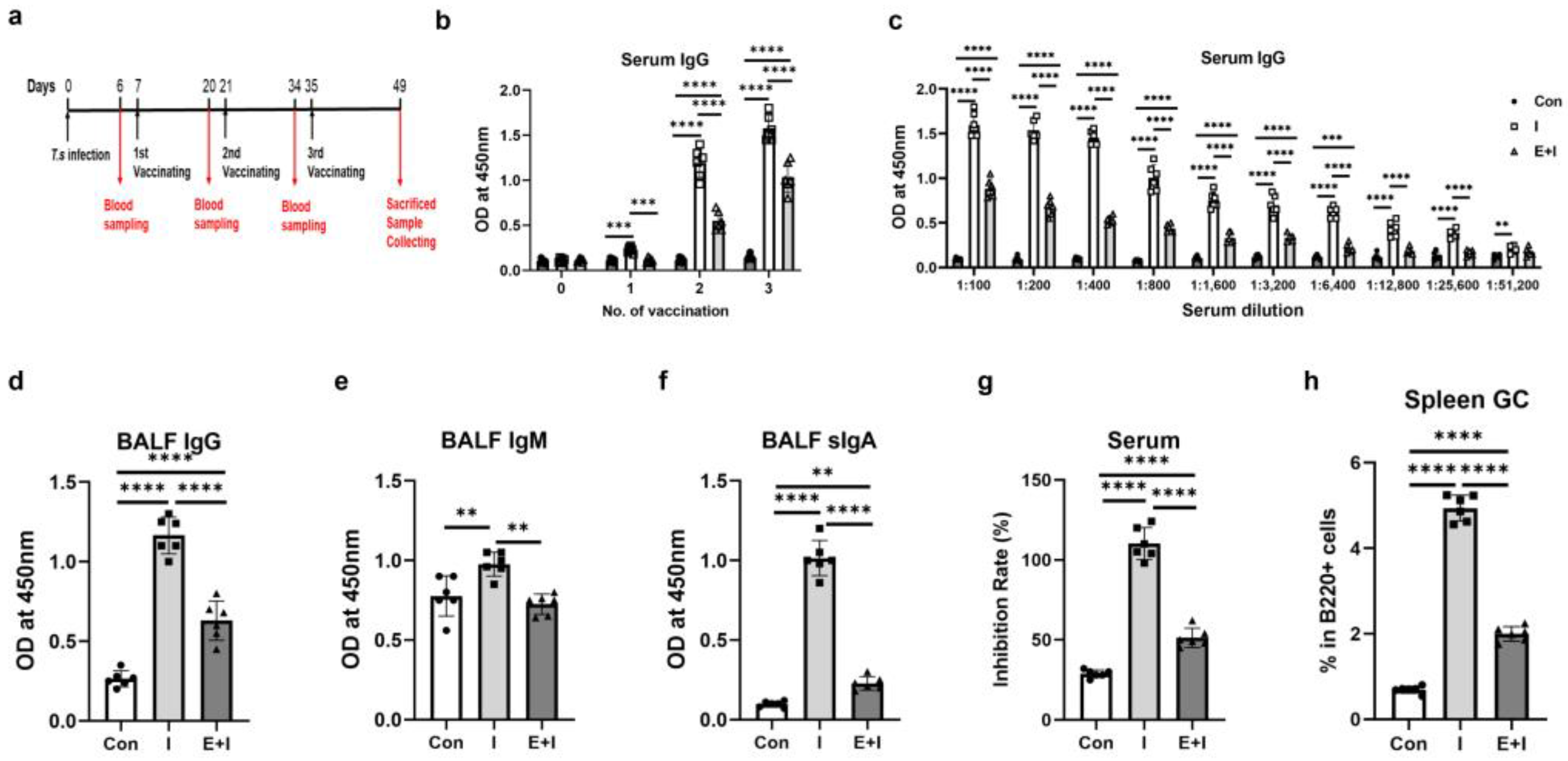
3.1. T. spiralis Infection Inhibits the Production of Anti-RBD-Specific Antibodies in Immunized Mice
3.2. Pre-Exposure to the Enteric Phase of Trichinosis Promotes Th2 and Treg Immune Responses in Immunized Mice
3.3. Pre-Exposure to the Muscular Phase of Trichinosis Promotes Th2 Immune Responses in Immunized Mice
3.4. ALB Treatment Partially Rescues the Inhibition of T. spiralis Infection on the Production of Anti-RBD-Specific Antibodies in Immunized Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Coronavirus Disease (COVID-19). Available online: https://www.who.int/news-room/fact-sheets/detail/coronavirus-disease-(covid-19) (accessed on 25 May 2024).
- Li, J.X.; Li, Z.P.; Zhu, Y.W.; Li, Z.; Zhu, F. Evidence of the efficacy and the effectiveness of first generation COVID-19 vaccines in clinical trials and real-world studies. Zoonoses 2022, 2, 30. [Google Scholar] [CrossRef]
- Li, M.; Wang, H.; Tian, L.; Pang, Z.; Yang, Q.; Huang, T.; Fan, J.; Song, L.; Tong, Y.; Fan, H. COVID-19 vaccine development: Milestones, lessons and prospects. Signal Transduct. Target. Ther. 2022, 7, 146. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, K.; Rzymski, P.; Islam, M.S.; Makuku, R.; Mushtaq, A.; Khan, A.; Ivanovska, M.; Makka, S.A.; Hashem, F.; Marquez, L.; et al. Cabral-Marques, and N. Rezaei. COVID-19 vaccinations: The unknowns, challenges, and hopes. J. Med. Virol. 2022, 94, 1336–1349. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, W.; Chen, Z.; Lu, S.; Yang, F.; Bi, Z.; Bao, L.; Mo, F.; Li, X.; Huang, Y.; et al. A vaccine targeting the RBD of the S protein of SARS-CoV-2 induces protective immunity. Nature 2020, 586, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Castro Dopico, X.; Ols, S.; Loré, K.; Karlsson Hedestam, G.B. Immunity to SARS-CoV-2 induced by infection or vaccination. J. Intern. Med. 2022, 291, 32–50. [Google Scholar] [CrossRef]
- Kabagenyi, J.; Natukunda, A.; Nassuuna, J.; Sanya, R.E.; Nampijja, M.; Webb, E.L.; Elliott, A.M.; Nkurunungi, G. Urban-rural differences in immune responses to mycobacterial and tetanus vaccine antigens in a tropical setting: A role for helminths? Parasitol. Int. 2020, 78, 102132. [Google Scholar] [CrossRef]
- Nono, J.K.; Kamdem, S.D.; Musaigwa, F.; Nnaji, C.A.; Brombacher, F. Influence of schistosomiasis on host vaccine responses. Trends Parasitol. 2022, 38, 67–79. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Factors That Influence the Immune Response to Vaccination. Clin. Microbiol. Rev. 2019, 32, e00084-18. [Google Scholar] [CrossRef]
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef]
- Maizels, R.M.; McSorley, H.J. Regulation of the host immune system by helminth parasites. J. Allergy Clin. Immunol. 2016, 138, 666–675. [Google Scholar] [CrossRef]
- Elias, D.; Akuffo, H.; Pawlowski, A.; Haile, M.; Schön, T.; Britton, S. Schistosoma mansoni infection reduces the protective efficacy of BCG vaccination against virulent Mycobacterium tuberculosis. Vaccine 2005, 23, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Hou, X.; Nie, G.; Xiao, Y.; Zhang, Q.; Liu, W.Q.; Li, Y.L.; Lei, J.H. Effect of Trichinella spiralis infection on the immune response to HBV vaccine in a mouse model. Foodborne Pathog. Dis. 2013, 10, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Loffredo-Verde, E.; Bhattacharjee, S.; Malo, A.; Festag, J.; Kosinska, A.D.; Ringelhan, M.; Rim Sarkar, S.; Steiger, K.; Heikenwaelder, M.; Protzer, U.; et al. Dynamic, Helminth-Induced Immune Modulation Influences the Outcome of Acute and Chronic Hepatitis B Virus Infection. J. Infect. Dis. 2020, 221, 1448–1461. [Google Scholar] [CrossRef] [PubMed]
- Stetter, N.; Hartmann, W.; Brunn, M.L.; Stanelle-Bertram, S.; Gabriel, G.; Breloer, M. A Combination of Deworming and Prime-Boost Vaccination Regimen Restores Efficacy of Vaccination Against Influenza in Helminth-Infected Mice. Front. Immunol. 2021, 12, 784141. [Google Scholar] [CrossRef] [PubMed]
- Dzhivhuho, G.A.; Rehrl, S.A.; Ndlovu, H.; Horsnell, W.G.C.; Brombacher, F.; Williamson, A.L.; Chege, G.K. Chronic schistosomiasis suppresses HIV-specific responses to DNA-MVA and MVA-gp140 Env vaccine regimens despite antihelminthic treatment and increases helminth-associated pathology in a mouse model. PLoS Pathog. 2018, 14, e1007182. [Google Scholar] [CrossRef] [PubMed]
- Musaigwa, F.; Kamdem, S.D.; Mpotje, T.; Mosala, P.; Abdel Aziz, N.; Herbert, D.R.; Brombacher, F.; Nono, J.K. Schistosoma mansoni infection induces plasmablast and plasma cell death in the bone marrow and accelerates the decline of host vaccine responses. PLoS Pathog. 2022, 18, e1010327. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, Z.; Sahin, O.S.; Yazar, S.; Bozok Cetintas, V. Genetic and epigenetic factors associated with increased severity of Covid-19. Cell Biol. Int. 2021, 45, 1158–1174. [Google Scholar] [CrossRef] [PubMed]
- Adjobimey, T.; Meyer, J.; Hennenfent, A.; Bara, A.J.; Lagnika, L.; Kocou, B.; Adjagba, M.; Laleye, A.; Hoerauf, A.; Parcina, M. Negative association between Ascaris lumbricoides seropositivity and Covid-19 severity: Insights from a study in Benin. Front. Immunol. 2023, 14, 1233082. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, R.S.; Piedrafita, D.; Greenhill, A.; Mahanty, S. Will helminth co-infection modulate COVID-19 severity in endemic regions? Nat. Rev. Immunol. 2020, 20, 342. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Alkazmi, L.; El-Bouseary, M.M.; Hamad, R.S.; Abdelhamid, M.; Batiha, G.E. The potential nexus between helminths and SARS-CoV-2 infection: A literature review. J. Immunol. Res. 2023, 2023, 5544819. [Google Scholar] [CrossRef]
- Abdoli, A. Helminths and COVID-19 co-infections: A neglected critical challenge. ACS Pharmacol. Transl. Sci. 2020, 3, 1039–1041. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, J.F. Parasites and Parasitology in this SARS-CoV-2, COVID-19 World: An American Society of Parasitologists Presidential Address. J. Parasitol. 2020, 106, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Apiwattanakul, N.; Thomas, P.G.; Iverson, A.R.; McCullers, J.A. Chronic helminth infections impair pneumococcal vaccine responses. Vaccine 2014, 32, 5405–5410. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, W.; Brunn, M.L.; Stetter, N.; Gagliani, N.; Muscate, F.; Stanelle-Bertram, S.; Gabriel, G.; Breloer, M. Helminth Infections Suppress the Efficacy of Vaccination against Seasonal Influenza. Cell Rep. 2019, 29, 2243–2256.e4. [Google Scholar] [CrossRef] [PubMed]
- Breloer, M.; Hartmann, W. Filarial infections compromise influenza vaccination efficacy: Lessons from the mouse. Immunol. Lett. 2023, 255, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Jiang, W.; Bai, Y.; Hou, X.; Jiang, C.; Zhang, C.; Jacques, M.L.; Liu, W.; Lei, J. Purinergic P2X7 Receptor Mediates the Elimination of Trichinella spiralis by Activating NF-κB/NLRP3/IL-1β Pathway in Macrophages. Infect. Immun. 2021, 89, e00683-20. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Cao, H.; Liu, C. SARS-CoV-2 S1 is superior to the RBD as a COVID-19 subunit vaccine antigen. J. Med. Virol. 2021, 93, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Bany, J.; Lach, J.; Golińska, Z. Efficacy of albendazole against Trichinella pseudospiralis and Trichinella spiralis in mice. Wiad. Parazytol. 1992, 38, 143–146. [Google Scholar] [PubMed]
- Guan, F.; Luo, X.; Liu, J.; Huang, Y.; Liu, Q.; Chang, J.; Fang, G.; Kang, D.; Gu, H.; Luo, L.; et al. GSDMA3 deficiency reprograms cellular metabolism and modulates BCR signaling in murine B cells. iScience 2023, 26, 107341. [Google Scholar] [CrossRef]
- Plotkin, S.A. Updates on immunologic correlates of vaccine-induced protection. Vaccine 2020, 38, 2250–2257. [Google Scholar] [CrossRef]
- Young, C.; Brink, R. The unique biology of germinal center B cells. Immunity 2021, 54, 1652–1664. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Duan, L.; Wang, C.; Zhang, C.; Hou, S.; Wang, H.; Song, J.; Zhang, T.; Li, Z.; Wang, M.; et al. Activation and induction of antigen-specific T follicular helper cells play a critical role in recombinant SARS-CoV-2 RBD vaccine-induced humoral responses. Mol. Biomed. 2023, 4, 34. [Google Scholar]
- Zhang, Z.; Mateus, J.; Coelho, C.H.; Dan, J.M.; Moderbacher, C.R.; Gálvez, R.I.; Cortes, F.H.; Grifoni, A.; Tarke, A.; Chang, J.; et al. Humoral and cellular immune memory to four COVID-19 vaccines. Cell 2022, 185, 2434–2451.e17. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, V.; Benincasa, G.; Moccia, G.; Sansone, A.; Signoriello, G.; Napoli, C. Evaluation of circulating leucocyte populations both in subjects with previous SARS-COV-2 infection and in healthy subjects after vaccination. J. Immunol. Methods 2022, 502, 113230. [Google Scholar] [CrossRef] [PubMed]
- Sikder, S.; Pierce, D.; Sarkar, E.R.; McHugh, C.; Quinlan, K.G.R.; Giacomin, P.; Loukas, A. Regulation of host metabolic health by parasitic helminths. Trends Parasitol. 2024, 40, 386–400. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, T.; Singh, B.; Rizvi, Z.A.; Verma, R.; Goswami, S.; Vishwakarma, P.; Jakhar, K.; Sonar, S.; Mani, S.; Bhattacharyya, S.; et al. Comparative immunomodulatory evaluation of the receptor binding domain of the SARS-CoV-2 spike protein; a potential vaccine candidate which imparts potent humoral and Th1 type immune response in a mouse model. Front. Immunol. 2021, 12, 641447. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Li, S.; Jin, X.; Han, J.B.; Xu, K.; Xu, S.; Han, Y.; Liu, C.; Zheng, T.; Liu, M.; et al. A tandem-repeat dimeric RBD protein-based COVID-19 vaccine ZF2001 protects mice and nonhuman primates. Emerg. Microbes Infect. 2022, 11, 1058–1071. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hardy, L.C.; Kapita, C.M.; Hall, J.A.; Arbeeva, L.; Campbell, E.; Urban, J.F.; Belkaid, Y.; Nagler, C.R.; Iweala, O.I. Intestinal helminth infection impairs oral and parenteric vaccine efficacy. J. Immunol. 2023, 211, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Bobat, S.; Darby, M.; Mrdjen, D.; Cook, C.; Logan, E.; Auret, J.; Jones, E.; Schnoeller, C.; Flores-Langarica, A.; Ross, E.A.; et al. Natural and vaccine-mediated immunity to Salmonella Typhimurium is impaired by the helminth Nippostrongylus brasiliensis. PLoS. Negl. Trop. Dis. 2014, 8, e3341. [Google Scholar] [CrossRef]
- Wammes, L.J.; Hamid, F.; Wiria, A.E.; de Gier, B.; Sartono, E.; Maizels, R.M.; Luty, A.J.; Fillié, Y.; Brice, G.T.; Supali, T.; et al. Regulatory T cells in human geohelminth infection suppress immune responses to BCG and Plasmodium falciparum. Eur. J. Immunol. 2010, 40, 437–442. [Google Scholar] [CrossRef]
- Grencis, R.K.; Hu¨ltner, L.; Else, K.J. Host protective immunity to Trichinella spiralis in mice: Activation of Th cell subsets and lymphokine secretion in mice expressing different response phenotypes. Immunology 1991, 74, 329–332. [Google Scholar] [PubMed]
- Lei, H.; Alu, A.; Yang, J.; Ren, W.; He, C.; Lan, T.; He, X.; Yang, L.; Li, J.; Wang, Z.; et al. Intranasal administration of a recombinant RBD vaccine induces long-term immunity against Omicron-included SARS-CoV-2 variants. Signal Transduct. Target. Ther. 2022, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Lederer, K.; Castaño, D.; Gómez Atria, D.; Oguin, T.H., 3rd; Wang, S.; Manzoni, T.B.; Muramatsu, H.; Hogan, M.J.; Amanat, F.; Cherubin, P.; et al. SARS-CoV-2 mRNA vaccines foster potent antigen-specific germinal center responses associated with neutralizing antibody generation. Immunity 2020, 53, 1281–1295. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Hong, K.J.; Kim, H.; Nam, J.H. Influenza vaccines: Past, present, and future. Rev. Med. Virol. 2022, 32, e2243. [Google Scholar] [CrossRef] [PubMed]
- van Riet, E.; Hartgers, F.C.; Yazdanbakhsh, M. Chronic helminth infections induce immunomodulation: Consequences and mechanisms. Immunobiology 2007, 212, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Egwang, T.G.; Owalla, T.J.; Kemigisha, M. COVID-19 vaccine trials must include helminth-infected cohorts. Nat. Immunol. 2022, 23, 148. [Google Scholar] [CrossRef]
- Adjobimey, T.; Meyer, J.; Terkeš, V.; Parcina, M.; Hoerauf, A. Helminth antigens differentially modulate the activation of CD4+ and CD8+ T lymphocytes of convalescent COVID-19 patients in vitro. BMC Med. 2022, 20, 241. [Google Scholar] [CrossRef] [PubMed]
- Elias, D.; Wolday, D.; Akuffo, H.; Petros, B.; Bronner, U.; Britton, S. Effect of deworming on human T cell responses to mycobacterial antigens in helminth-exposed individuals before and after bacille Calmette-Guérin (BCG) vaccination. Clin. Exp. Immunol. 2001, 123, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Elias, D.; Britton, S.; Aseffa, A.; Engers, H.; Akuffo, H. Poor immunogenicity of BCG in helminth infected population is associated with increased in vitro TGF-beta production. Vaccine 2008, 26, 3897–3902. [Google Scholar] [CrossRef]
- Cooper, P.J.; Chico, M.E.; Losonsky, G.; Sandoval, C.; Espinel, I.; Sridhara, R.; Aguilar, M.; Guevara, A.; Guderian, R.H.; Levine, M.M.; et al. Albendazole treatment of children with ascariasis enhances the vibriocidal antibody response to the live attenuated oral cholera vaccine CVD 103-HgR. J. Infect. Dis. 2000, 182, 1199–1206. [Google Scholar] [CrossRef]
- Van Dorst, M.M.A.R.; Pyuza, J.J.; Nkurunungi, G.; Kullaya, V.I.; Smits, H.H.; Hogendoorn, P.C.W.; Wammes, L.J.; Everts, B.; Elliott, A.M.; Jochems, S.P.; et al. Immunological factors linked to geographical variation in vaccine responses. Nat. Rev. Immunol. 2024, 24, 250–263. [Google Scholar] [CrossRef]
- Zhu, F.; Liu, W.; Liu, T.; Shi, L.; Zheng, W.; Guan, F.; Lei, J. A new role for old friends: Effects of helminth infections on vaccine efficacy. Pathogens 2022, 11, 1163. [Google Scholar] [CrossRef]
- Cooper, P.J.; Chico, M.E.; Sandoval, C.; Espinel, I.; Guevara, A.; Levine, M.M.; Griffin, G.E.; Nutman, T.B. Human infection with Ascaris lumbricoides is associated with suppression of the interleukin-2 response to recombinant cholera toxin B subunit following vaccination with the live oral cholera vaccine CVD 103-HgR. Infect. Immun. 2001, 69, 1574–1580. [Google Scholar] [CrossRef] [PubMed]
- Brückner, S.; Agnandji, S.T.; Berberich, S.; Bache, E.; Fernandes, J.F.; Schweiger, B.; Massinga Loembe, M.; Engleitner, T.; Lell, B.; Mordmüller, B.; et al. Effect of Antihelminthic Treatment on Vaccine Immunogenicity to a Seasonal Influenza Vaccine in Primary School Children in Gabon: A Randomized Placebo-Controlled Trial. PLoS Negl. Trop. Dis. 2015, 9, e0003768. [Google Scholar] [CrossRef]
- Brückner, S.; Agnandji, S.T.; Elias, J.; Berberich, S.; Bache, E.; Fernandes, J.; Loembe, M.M.; Hass, J.; Lell, B.; Mordmüller, B.; et al. A single-dose antihelminthic treatment does not influence immunogenicity of a meningococcal and a cholera vaccine in Gabonese school children. Vaccine 2016, 34, 5384–5390. [Google Scholar] [CrossRef] [PubMed]
- Wait, L.F.; Dobson, A.P.; Graham, A.L. Do parasite infections interfere with immunisation? A review and meta-analysis. Vaccine 2020, 38, 5582–5590. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.; Tang, X.; Xiao, T.; Qiao, Y.; Tian, X.; Zhu, B.; Chen, J.; Chen, C.; Li, Y.; Lin, X.; et al. Enhancing antitumor efficacy of oncolytic virus M1 via albendazole-sustained CD8+ T cell activation. Oncology 2024, 32, 200813. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, S.K.; Singh, A.; Gupta, K.K.; Khatoon, J.; Prasad, A.; Rai, R.P.; Gupta, R.K.; Tripathi, M.; Husain, N. Immune response to Taenia solium cysticerci after anti-parasitic therapy. Int. J. Parasitol. 2015, 45, 749–759. [Google Scholar] [CrossRef]
- Mamykova, O.I. Experimental Study of Albendazole Immunotropic Activity in Low Doses. Russ. Agric. Sci. 2021, 47, 88–92. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, F.; Zheng, W.; Gong, Y.; Zhang, J.; Yu, Y.; Zhang, J.; Liu, M.; Guan, F.; Lei, J. Trichinella spiralis Infection Inhibits the Efficacy of RBD Protein of SARS-CoV-2 Vaccination via Regulating Humoral and Cellular Immunity. Vaccines 2024, 12, 729. https://doi.org/10.3390/vaccines12070729
Zhu F, Zheng W, Gong Y, Zhang J, Yu Y, Zhang J, Liu M, Guan F, Lei J. Trichinella spiralis Infection Inhibits the Efficacy of RBD Protein of SARS-CoV-2 Vaccination via Regulating Humoral and Cellular Immunity. Vaccines. 2024; 12(7):729. https://doi.org/10.3390/vaccines12070729
Chicago/Turabian StyleZhu, Feifan, Wenwen Zheng, Yiyan Gong, Jinyuan Zhang, Yihan Yu, Jixian Zhang, Mengjun Liu, Fei Guan, and Jiahui Lei. 2024. "Trichinella spiralis Infection Inhibits the Efficacy of RBD Protein of SARS-CoV-2 Vaccination via Regulating Humoral and Cellular Immunity" Vaccines 12, no. 7: 729. https://doi.org/10.3390/vaccines12070729