
Influenza A Vaccine Candidates Based on Virus-like Particles Formed by Coat Proteins of Single-Stranded RNA Phages Beihai32 and PQ465

 , , , , , ,
, , , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
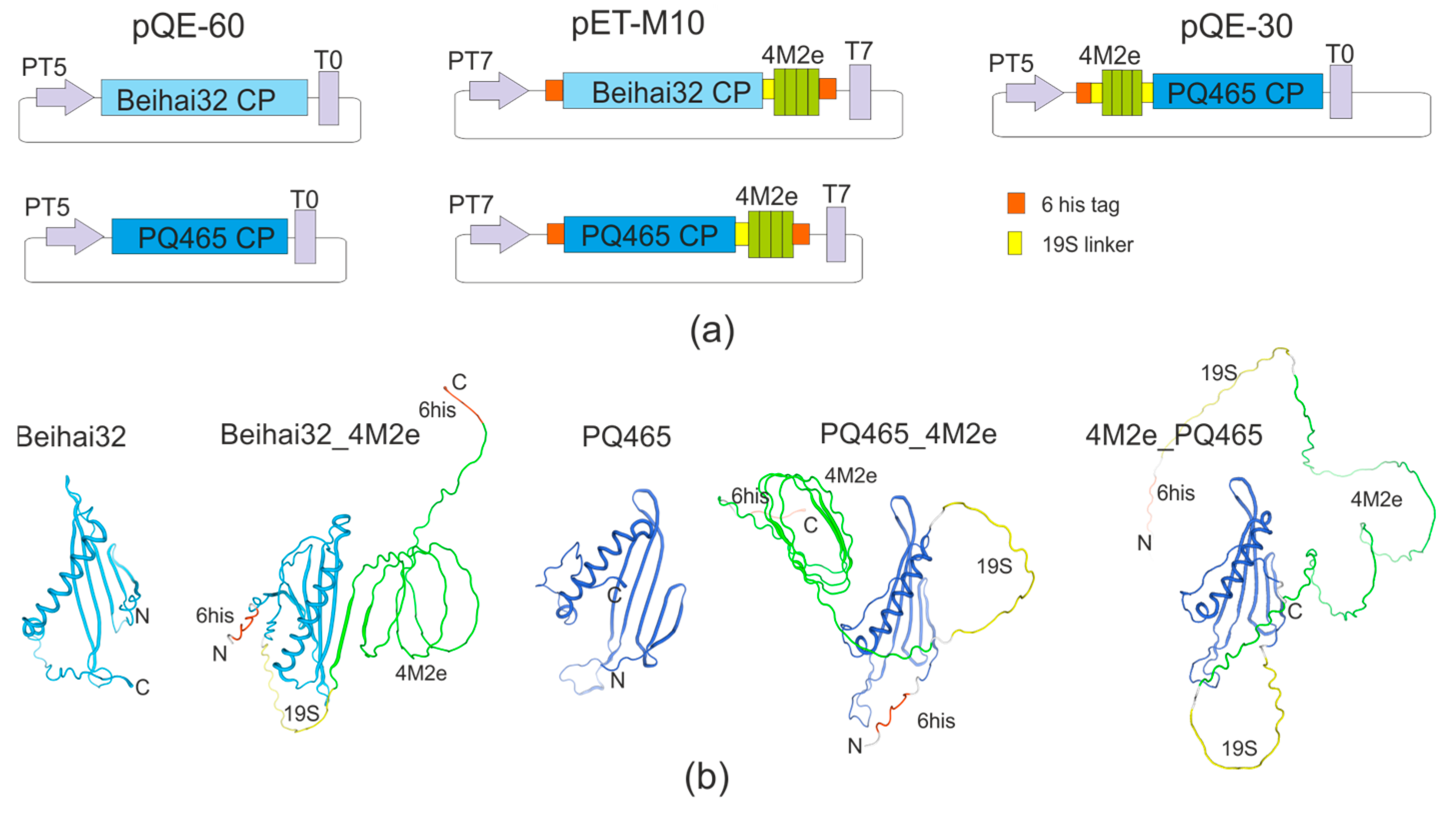
2.1. Construction of Expression Vectors
2.2. Expression of Recombinant Proteins
2.3. Purification of VLPs
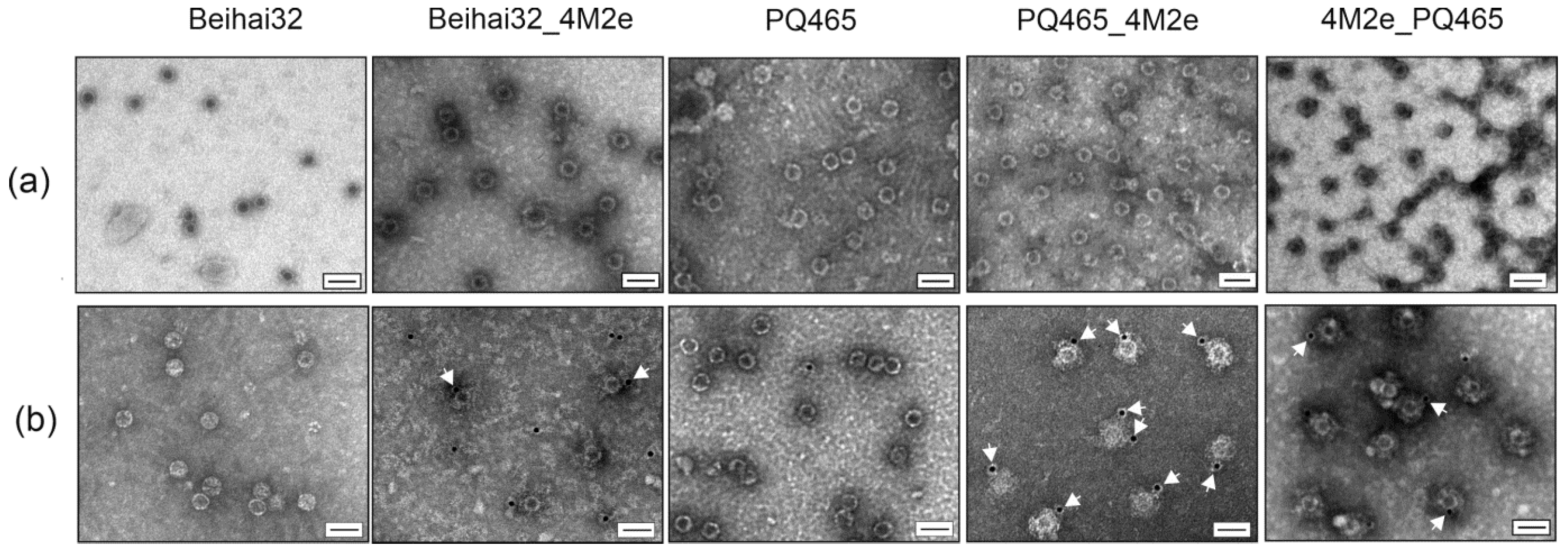
2.4. Electron Microscopy
2.5. Immunogold Transmission Electron Microscopy
2.6. Analysis of Antigenic Properties of VLPs by ELISA
2.7. Immunization of Mice
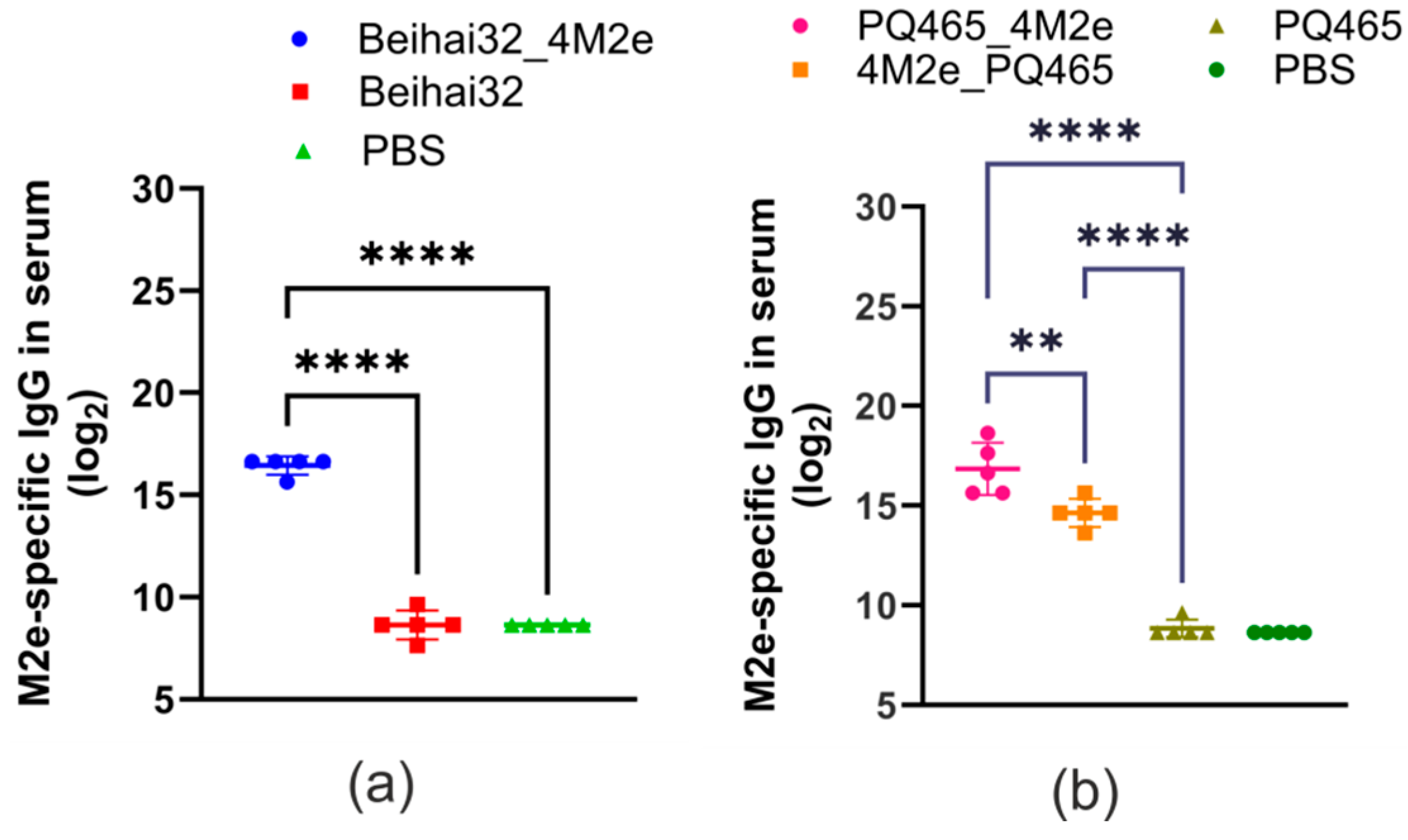
2.8. Analysis of Anti-M2e Antibody Titers by ELISA
2.9. Influenza Virus Challenge of Mice
2.10. Statistical Analysis
2.11. Ethics Statement
3. Results
3.1. Chimeric VLPs Carrying the M2e Peptide of the Influenza Virus: Design, Expression, and Purification
3.2. Exposure of the 4M2e Peptide on the Chimeric VLPs
3.3. Immunogenicity and Protective Activity of Chimeric VLPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pushko, P.; Tretyakova, I. Influenza Virus Like Particles (VLPs): Opportunities for H7N9 Vaccine Development. Viruses 2020, 12, 518. [Google Scholar] [CrossRef] [PubMed]
- Dormitzer, P.R.; Galli, G.; Castellino, F.; Golding, H.; Khurana, S.; Del Giudice, G.; Rappuoli, R. Influenza Vaccine Immunology. Immunol. Rev. 2011, 239, 167–177. [Google Scholar] [CrossRef]
- Shao, W.; Li, X.; Goraya, M.; Wang, S.; Chen, J.-L. Evolution of Influenza A Virus by Mutation and Re-Assortment. Int. J. Mol. Sci. 2017, 18, 1650. [Google Scholar] [CrossRef] [PubMed]
- Fiers, W.; De Filette, M.; El Bakkouri, K.; Schepens, B.; Roose, K.; Schotsaert, M.; Birkett, A.; Saelens, X. M2e-Based Universal Influenza A Vaccine. Vaccine 2009, 27, 6280–6283. [Google Scholar] [CrossRef]
- Tan, M.P.; Tan, W.S.; Mohamed Alitheen, N.B.; Yap, W.B. M2e-Based Influenza Vaccines with Nucleoprotein: A Review. Vaccines 2021, 9, 739. [Google Scholar] [CrossRef]
- Mezhenskaya, D.; Isakova-Sivak, I.; Rudenko, L. M2e-Based Universal Influenza Vaccines: A Historical Overview and New Approaches to Development. J. Biomed. Sci. 2019, 26, 76. [Google Scholar] [CrossRef]
- Neirynck, S.; Deroo, T.; Saelens, X.; Vanlandschoot, P.; Jou, W.M.; Fiers, W. A Universal Influenza A Vaccine Based on the Extracellular Domain of the M2 Protein. Nat. Med. 1999, 5, 1157–1163. [Google Scholar] [CrossRef]
- Kolpe, A.; Schepens, B.; Fiers, W.; Saelens, X. M2-Based Influenza Vaccines: Recent Advances and Clinical Potential. Expert Rev. Vaccines 2017, 16, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Mardanova, E.S.; Ravin, N.V. Plant-Produced Recombinant Influenza A Vaccines Based on the M2e Peptide. Curr. Pharm. Des. 2018, 24, 1317–1324. [Google Scholar] [CrossRef]
- Zykova, A.A.; Blokhina, E.A.; Stepanova, L.A.; Shuklina, M.A.; Tsybalova, L.M.; Kuprianov, V.V.; Ravin, N.V. Nanoparticles based on artificial self-assembling peptide and displaying M2e peptide and stalk HA epitopes of influenza A virus induce potent humoral and T-cell responses and protect against the viral infection. Nanomedicine 2022, 39, 102463. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine Delivery: A Matter of Size, Geometry, Kinetics and Molecular Patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Zepeda-Cervantes, J.; Ramírez-Jarquín, J.O.; Vaca, L. Interaction Between Virus-Like Particles (VLPs) and Pattern Recognition Receptors (PRRs) From Dendritic Cells (DCs): Toward Better Engineering of VLPs. Front. Immunol. 2020, 11, 1100. [Google Scholar] [CrossRef] [PubMed]
- Tars, K. ssRNA Phages: Life Cycle, Structure and Applications. In Biocommunication of Phages; Springer International Publishing: Cham, Switzerland, 2020; pp. 261–292. [Google Scholar]
- Tumban, E.; Peabody, J.; Tyler, M.; Peabody, D.S.; Chackerian, B. VLPs Displaying a Single L2 Epitope Induce Broadly Cross-Neutralizing Antibodies against Human Papillomavirus. PLoS ONE 2012, 7, e49751. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Yadav, R.; Kunda, N.K.; Anderson, D.; Bruckner, E.; Miller, E.K.; Basu, R.; Muttil, P.; Tumban, E. Oral Immunization with Bacteriophage MS2-L2 VLPs Protects against Oral and Genital Infection with Multiple HPV Types Associated with Head & Neck Cancers and Cervical Cancer. Antivir. Res. 2019, 166, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Ord, R.L.; Caldeira, J.C.; Rodriguez, M.; Noe, A.; Chackerian, B.; Peabody, D.S.; Gutierrez, G.; Lobo, C.A. A Malaria Vaccine Candidate Based on an Epitope of the Plasmodium Falciparum RH5 Protein. Malar. J. 2014, 13, 326. [Google Scholar] [CrossRef] [PubMed]
- Jegerlehner, A.; Zabel, F.; Langer, A.; Dietmeier, K.; Jennings, G.T.; Saudan, P.; Bachmann, M.F. Bacterially Produced Recombinant Influenza Vaccines Based on Virus-Like Particles. PLoS ONE 2013, 8, e78947. [Google Scholar] [CrossRef] [PubMed]
- Tissot, A.C.; Renhofa, R.; Schmitz, N.; Cielens, I.; Meijerink, E.; Ose, V.; Jennings, G.T.; Saudan, P.; Pumpens, P.; Bachmann, M.F. Versatile Virus-Like Particle Carrier for Epitope Based Vaccines. PLoS ONE 2010, 5, e9809. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, S.R.; Janowski, A.B.; Zhao, G.; Barouch, D.; Wang, D. Hyperexpansion of RNA Bacteriophage Diversity. PLoS Biol. 2016, 14, e1002409. [Google Scholar] [CrossRef] [PubMed]
- Liekniņa, I.; Kalniņš, G.; Akopjana, I.; Bogans, J.; Šišovs, M.; Jansons, J.; Rūmnieks, J.; Tārs, K. Production and Characterization of Novel ssRNA Bacteriophage Virus-like Particles from Metagenomic Sequencing Data. J. Nanobiotechnol. 2019, 17, 61. [Google Scholar] [CrossRef]
- Liekniņa, I.; Černova, D.; Rūmnieks, J.; Tārs, K. Novel ssRNA Phage VLP Platform for Displaying Foreign Epitopes by Genetic Fusion. Vaccine 2020, 38, 6019–6026. [Google Scholar] [CrossRef]
- Rūmnieks, J.; Liekniņa, I.; Kalniņš, G.; Šišovs, M.; Akopjana, I.; Bogans, J.; Tārs, K. Three-Dimensional Structure of 22 Uncultured ssRNA Bacteriophages: Flexibility of the Coat Protein Fold and Variations in Particle Shapes. Sci. Adv. 2020, 6, eabc0023. [Google Scholar] [CrossRef] [PubMed]
- Shishovs, M.; Rumnieks, J.; Diebolder, C.; Jaudzems, K.; Andreas, L.B.; Stanek, J.; Kazaks, A.; Kotelovica, S.; Akopjana, I.; Pintacuda, G.; et al. Structure of AP205 Coat Protein Reveals Circular Permutation in ssRNA Bacteriophages. J. Mol. Biol. 2016, 428, 4267–4279. [Google Scholar] [CrossRef]
- Dümmler, A.; Lawrence, A.-M.; de Marco, A. Simplified Screening for the Detection of Soluble Fusion Constructs Expressed in E. coli Using a Modular Set of Vectors. Microb. Cell Fact. 2005, 4, 34. [Google Scholar] [CrossRef] [PubMed]
- De Filette, M.; Jou, W.M.; Birkett, A.; Lyons, K.; Schultz, B.; Tonkyro, A.; Resch, S.; Fiers, W. Universal Influenza A Vaccine: Optimization of M2-Based Constructs. Virology 2005, 337, 149–161. [Google Scholar] [CrossRef]
- Robinson, C.R.; Sauer, R.T. Optimizing the Stability of Single-Chain Proteins by Linker Length and Composition Mutagenesis. Proc. Natl. Acad. Sci. USA 1998, 95, 5929–5934. [Google Scholar] [CrossRef]
- Ravin, N.V.; Blokhina, E.A.; Kuprianov, V.V.; Stepanova, L.A.; Shaldjan, A.A.; Kovaleva, A.A.; Tsybalova, L.M.; Skryabin, K.G. Development of a Candidate Influenza Vaccine Based on Virus-like Particles Displaying Influenza M2e Peptide into the Immunodominant Loop Region of Hepatitis B Core Antigen: Insertion of Multiple Copies of M2e Increases Immunogenicity and Protective Efficiency. Vaccine 2015, 33, 3392–3397. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling Protein Tertiary and Quaternary Structure Using Evolutionary Information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like Particles: Preparation, Immunogenicity and Their Roles as Nanovaccines and Drug Nanocarriers. J. Nanobiotechnol. 2021, 19, 59. [Google Scholar] [CrossRef]
- Hemmati, F.; Hemmati-Dinarvand, M.; Karimzade, M.; Rutkowska, D.; Eskandari, M.H.; Khanizadeh, S.; Afsharifar, A. Plant-Derived VLP: A Worthy Platform to Produce Vaccine against SARS-CoV-2. Biotechnol. Lett. 2022, 44, 45–57. [Google Scholar] [CrossRef]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major Findings and Recent Advances in Virus–like Particle (VLP)-Based Vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, N.; Chuan, Y.P.; Lua, L.H.L.; Middelberg, A.P.J. Modular Engineering of a Microbially-Produced Viral Capsomere Vaccine for Influenza. Chem. Eng. Sci. 2013, 103, 12–20. [Google Scholar] [CrossRef]
- Bessa, J.; Schmitz, N.; Hinton, H.J.; Schwarz, K.; Jegerlehner, A.; Bachmann, M.F. Efficient induction of mucosal and systemic immune responses by virus-like particles administered intranasally: Implications for vaccine design. Eur. J. Immunol. 2008, 38, 114–126. [Google Scholar] [CrossRef]
- Khalaj-Hedayati, A.; Chua, C.L.L.; Smooker, P.; Lee, K.W. Nanoparticles in Influenza Subunit Vaccine Development: Immunogenicity Enhancement. Influenza Other Respir. Viruses 2020, 14, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Kirsteina, A.; Akopjana, I.; Bogans, J.; Lieknina, I.; Jansons, J.; Skrastina, D.; Kazaka, T.; Tars, K.; Isakova-Sivak, I.; Mezhenskaya, D.; et al. Construction and Immunogenicity of a Novel Multivalent Vaccine Prototype Based on Conserved Influenza Virus Antigens. Vaccines 2020, 8, 197. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, Z.; Golchin, M.; Khalili-Yazdi, A.; Khalili, M. Immunological Properties of the SLLTEVET Epitope of Influenza A Virus in Multiple Display on Filamentous M13 Phage. Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Ibañez, L.I.; Van den Bossche, V.; Roose, K.; Youssef, S.A.; de Bruin, A.; Fiers, W.; Saelens, X. Protection against Influenza A Virus Challenge with M2e-Displaying Filamentous Escherichia coli Phages. PLoS ONE 2015, 10, e0126650. [Google Scholar] [CrossRef]
- Li, M.; Chen, C.; Wang, X.; Guo, P.; Feng, H.; Zhang, X.; Zhang, W.; Gu, C.; Zhu, J.; Wen, G.; et al. T4 Bacteriophage Nanoparticles Engineered through CRISPR Provide a Versatile Platform for Rapid Development of Flu Mucosal Vaccines. Antivir. Res. 2023, 217, 105688. [Google Scholar] [CrossRef]
- Li, M.; Guo, P.; Chen, C.; Feng, H.; Zhang, W.; Gu, C.; Wen, G.; Rao, V.B.; Tao, P. Bacteriophage T4 Vaccine Platform for Next-Generation Influenza Vaccine Development. Front. Immunol. 2021, 12, 745625. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wang, Y.W.; Tang, Y.H.; Zheng, Q.S.; Hou, J.B. Construction and Immunogenicity of Recombinant Bacteriophage T7 Vaccine Expressing M2e Peptides of Avian Influenza Virus. Bing Du Xue Bao 2013, 29, 376–381. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasyagin, E.A.; Zykova, A.A.; Mardanova, E.S.; Nikitin, N.A.; Shuklina, M.A.; Ozhereleva, O.O.; Stepanova, L.A.; Tsybalova, L.M.; Blokhina, E.A.; Ravin, N.V. Influenza A Vaccine Candidates Based on Virus-like Particles Formed by Coat Proteins of Single-Stranded RNA Phages Beihai32 and PQ465. Vaccines 2024, 12, 1033. https://doi.org/10.3390/vaccines12091033
Vasyagin EA, Zykova AA, Mardanova ES, Nikitin NA, Shuklina MA, Ozhereleva OO, Stepanova LA, Tsybalova LM, Blokhina EA, Ravin NV. Influenza A Vaccine Candidates Based on Virus-like Particles Formed by Coat Proteins of Single-Stranded RNA Phages Beihai32 and PQ465. Vaccines. 2024; 12(9):1033. https://doi.org/10.3390/vaccines12091033
Chicago/Turabian StyleVasyagin, Egor A., Anna A. Zykova, Eugenia S. Mardanova, Nikolai A. Nikitin, Marina A. Shuklina, Olga O. Ozhereleva, Liudmila A. Stepanova, Liudmila M. Tsybalova, Elena A. Blokhina, and Nikolai V. Ravin. 2024. "Influenza A Vaccine Candidates Based on Virus-like Particles Formed by Coat Proteins of Single-Stranded RNA Phages Beihai32 and PQ465" Vaccines 12, no. 9: 1033. https://doi.org/10.3390/vaccines12091033
APA StyleVasyagin, E. A., Zykova, A. A., Mardanova, E. S., Nikitin, N. A., Shuklina, M. A., Ozhereleva, O. O., Stepanova, L. A., Tsybalova, L. M., Blokhina, E. A., & Ravin, N. V. (2024). Influenza A Vaccine Candidates Based on Virus-like Particles Formed by Coat Proteins of Single-Stranded RNA Phages Beihai32 and PQ465. Vaccines, 12(9), 1033. https://doi.org/10.3390/vaccines12091033