Protective Immunity against Canine Distemper Virus in Dogs Induced by Intranasal Immunization with a Recombinant Probiotic Expressing the Viral H Protein

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Virus, and Plasmid
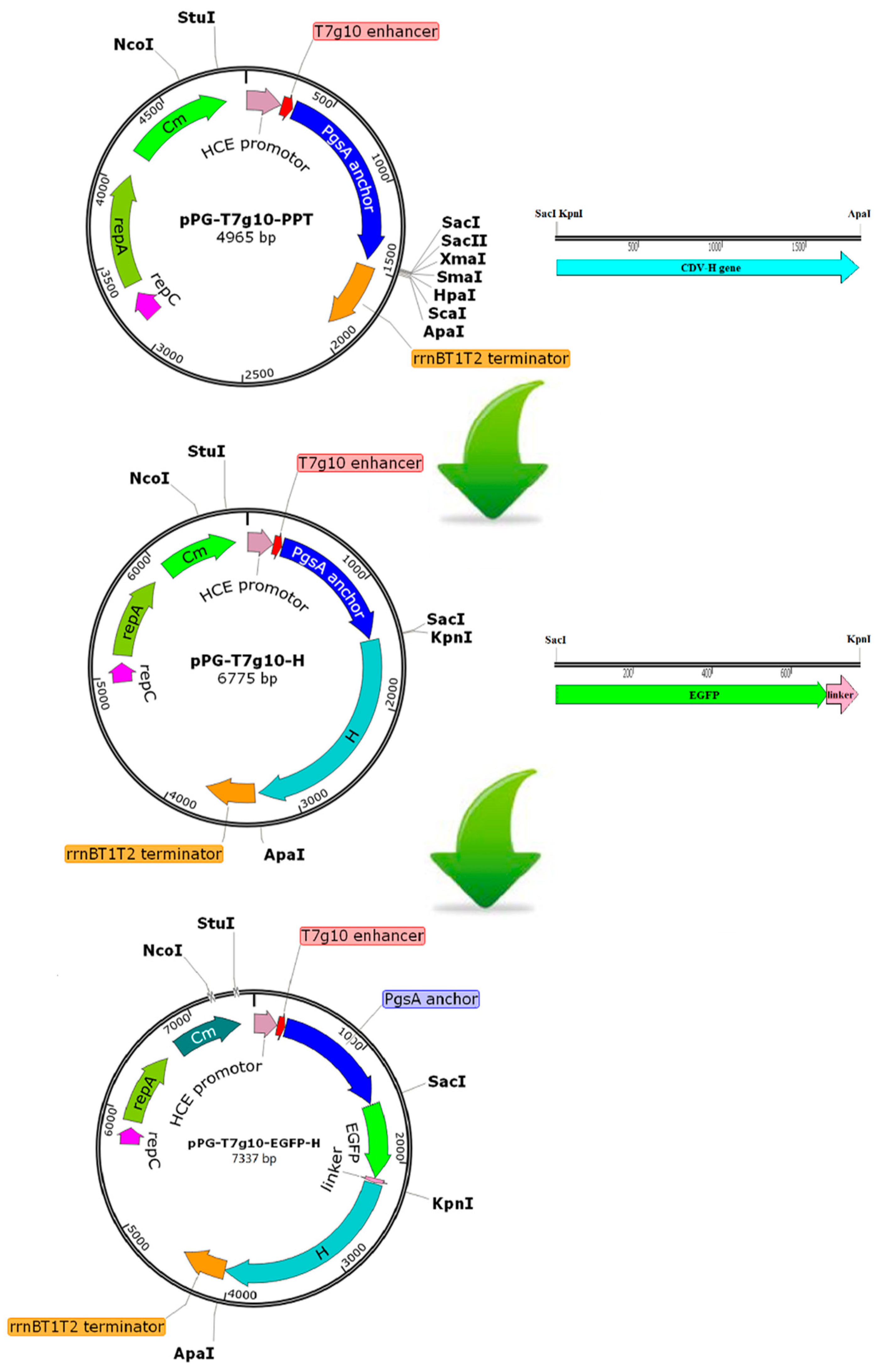
2.2. Construction of Recombinant Lactobacillus Strain
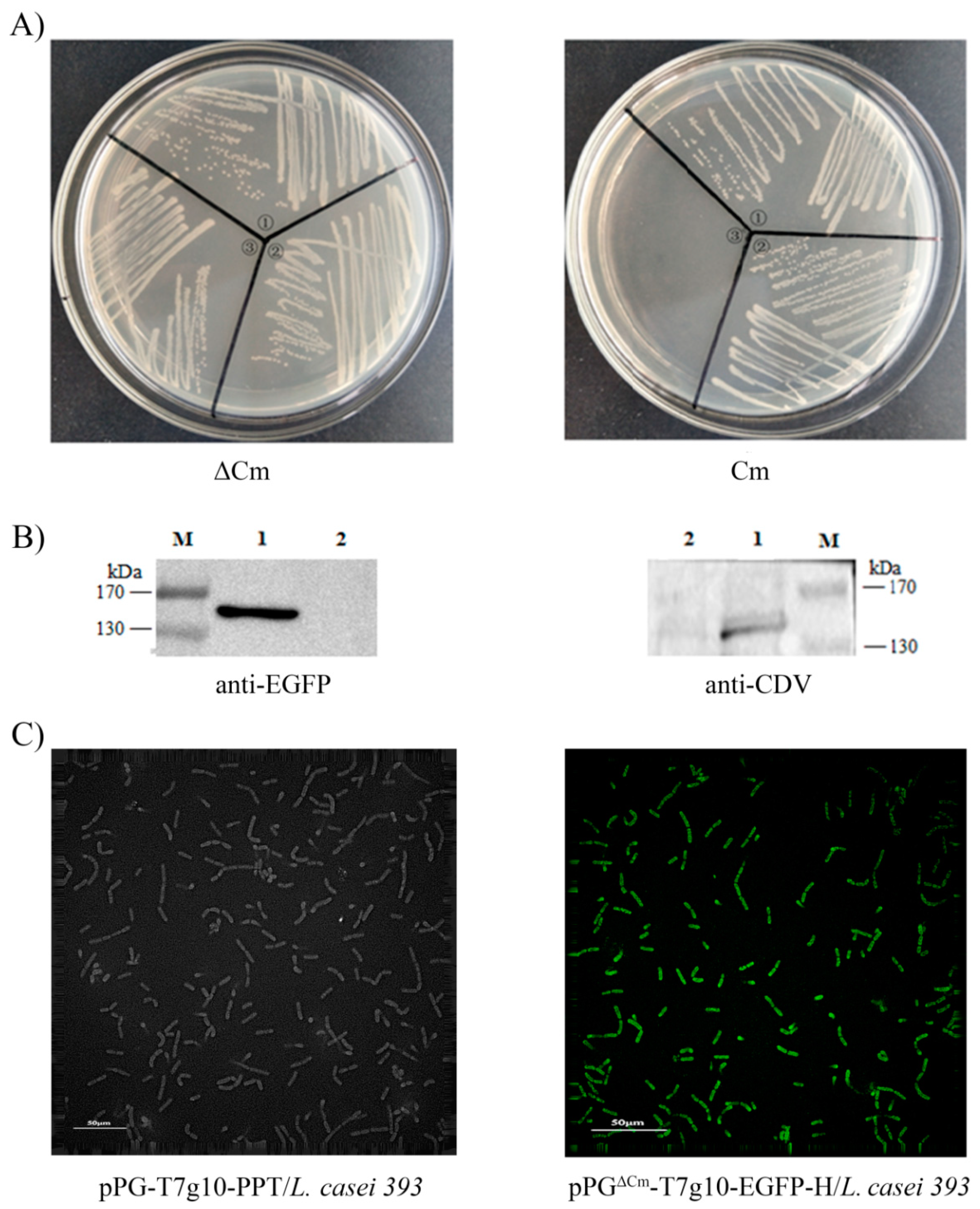
2.3. Identification of Proteins Expressed by the Recombinant Lactobacillus
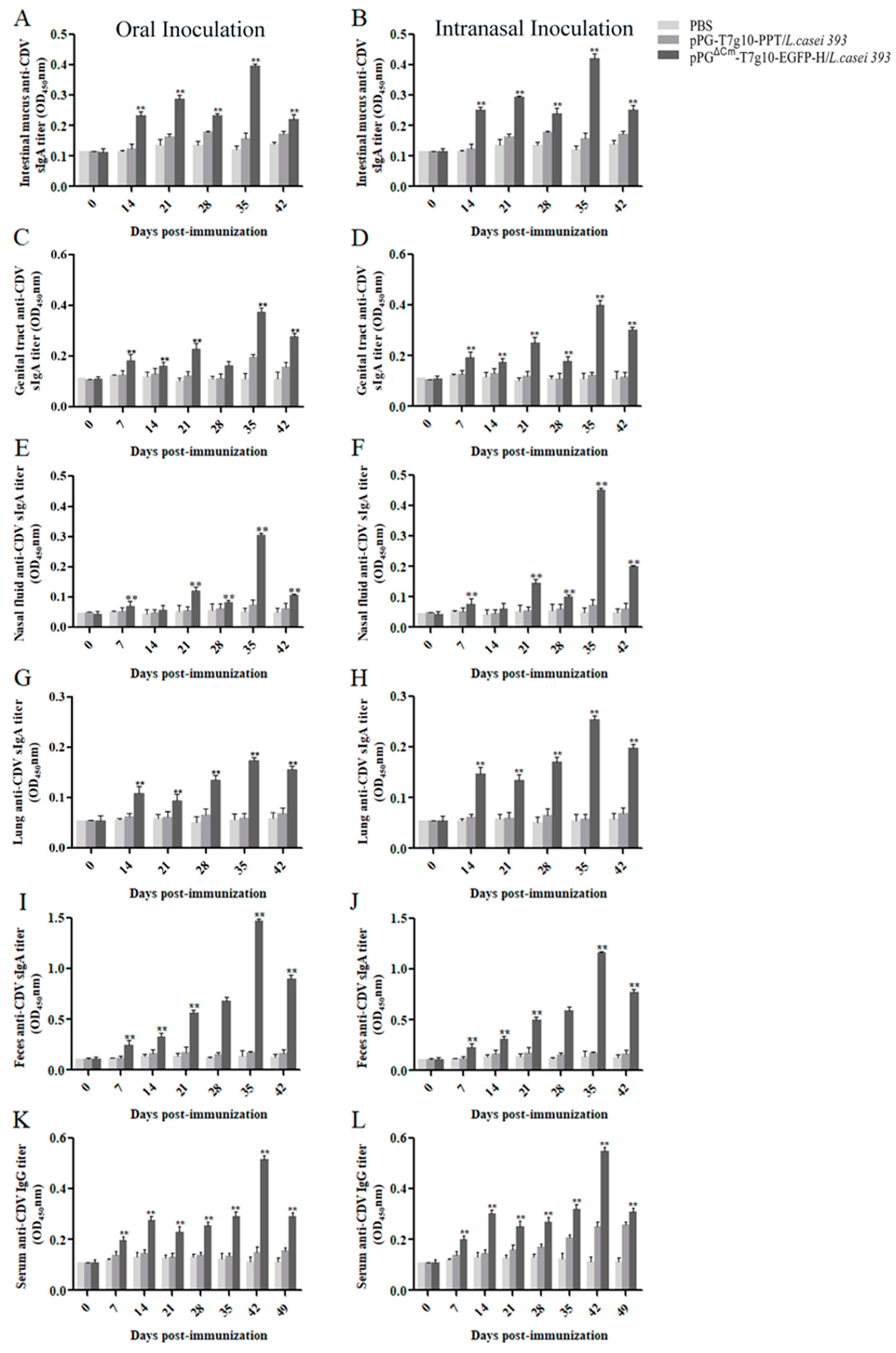
2.4. Immunization and Specimen Collection
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Detection of Recombinant L. Casei 393 on Specific T Cell and Lymphocyte Proliferation in Mice
2.7. Canine Distemper Virus (CDV) Neutralization by immune Dog Sera
2.8. Challenge Experiment in Dogs
2.9. Real-Time RT-PCR (qRT-PCR) Analysis
2.10. Statistical Analysis
3. Results
3.1. Construction of the Recombinant Lactobacillus
3.2. Immune Responses Induced in Mice
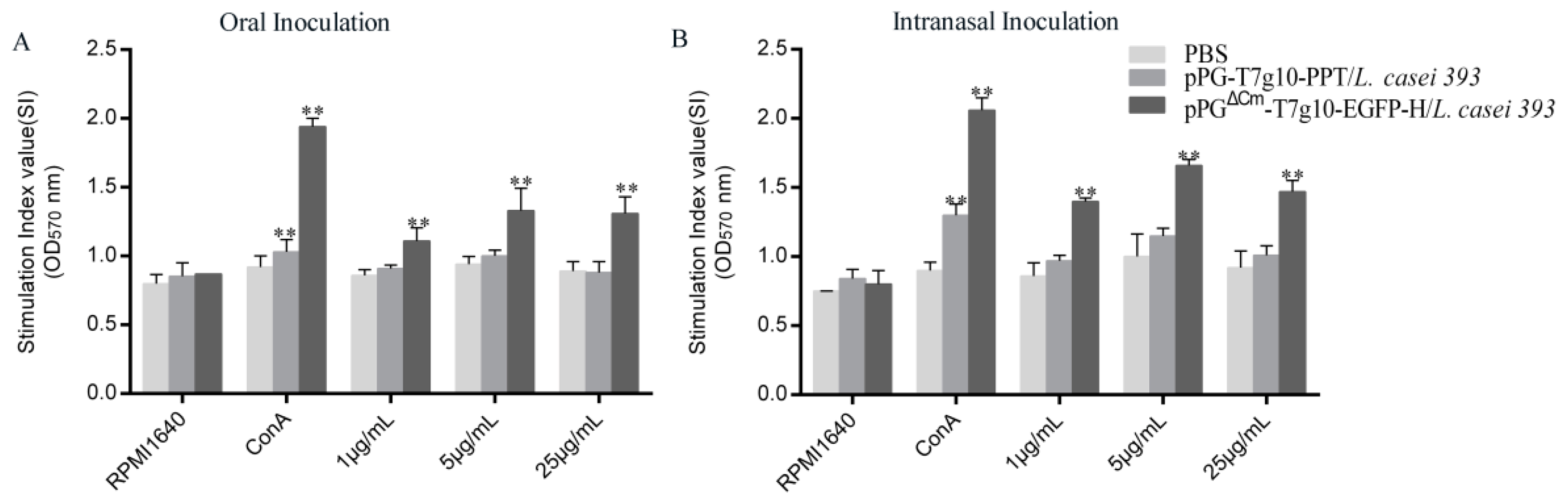
3.3. Effect of Recombinant L. casei 393 on Specific T Cell and Lymphocyte Proliferation in Mice
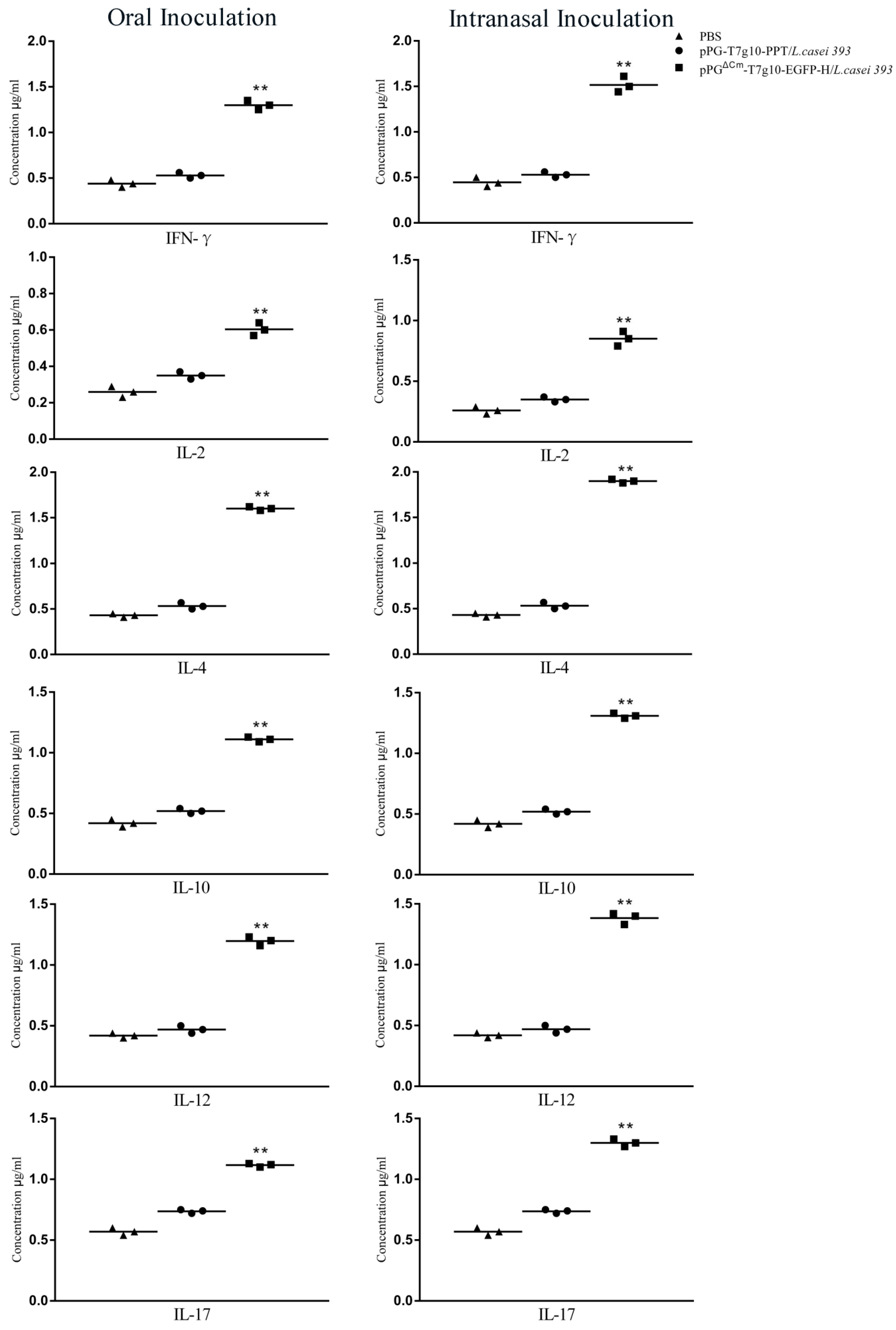
3.4. Cytokine Levels in Mice
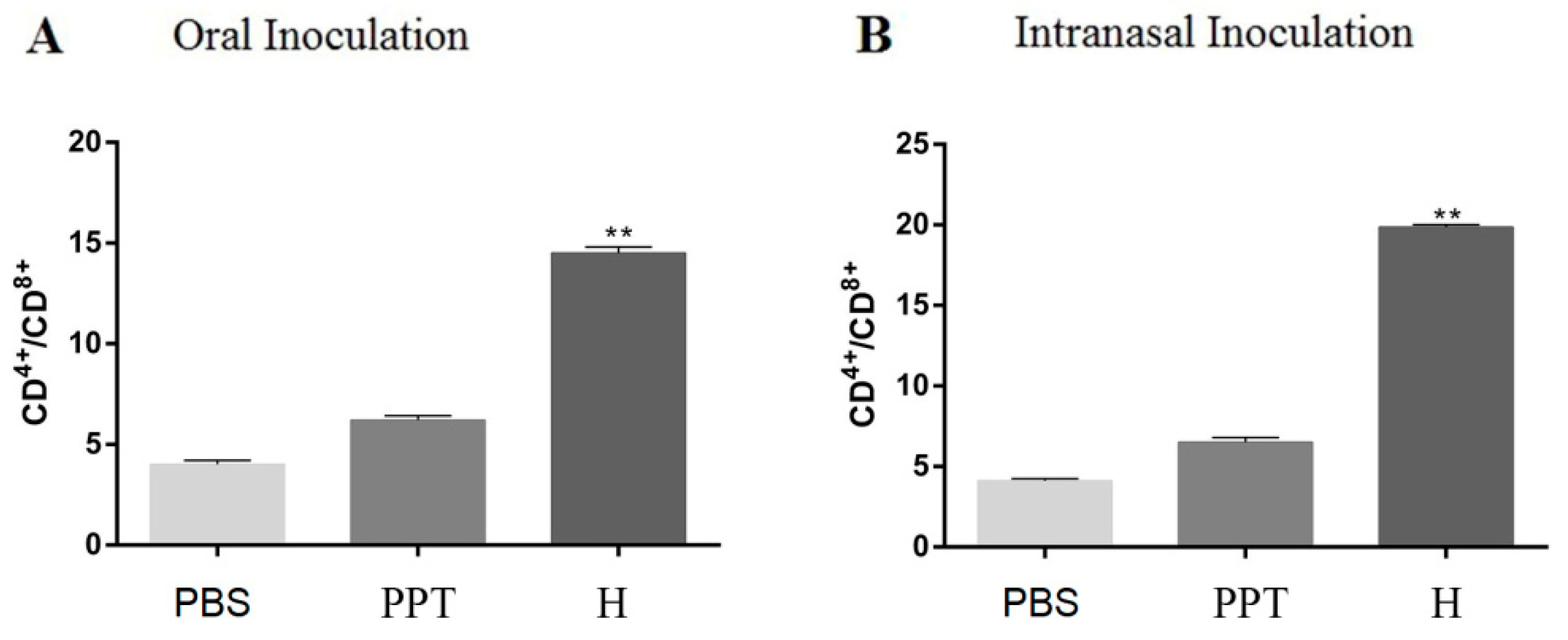
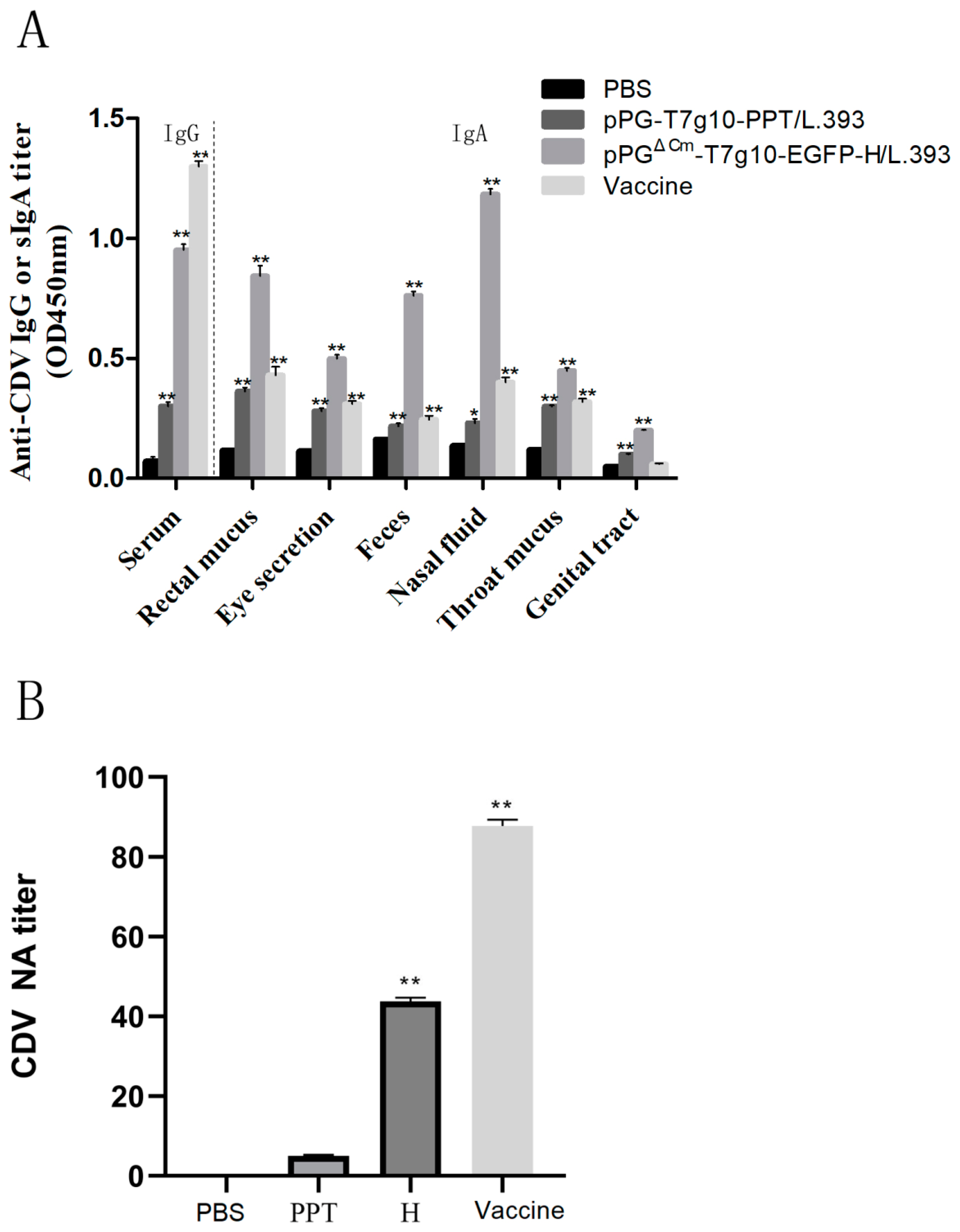
3.5. Immune Response Induced in Dogs.
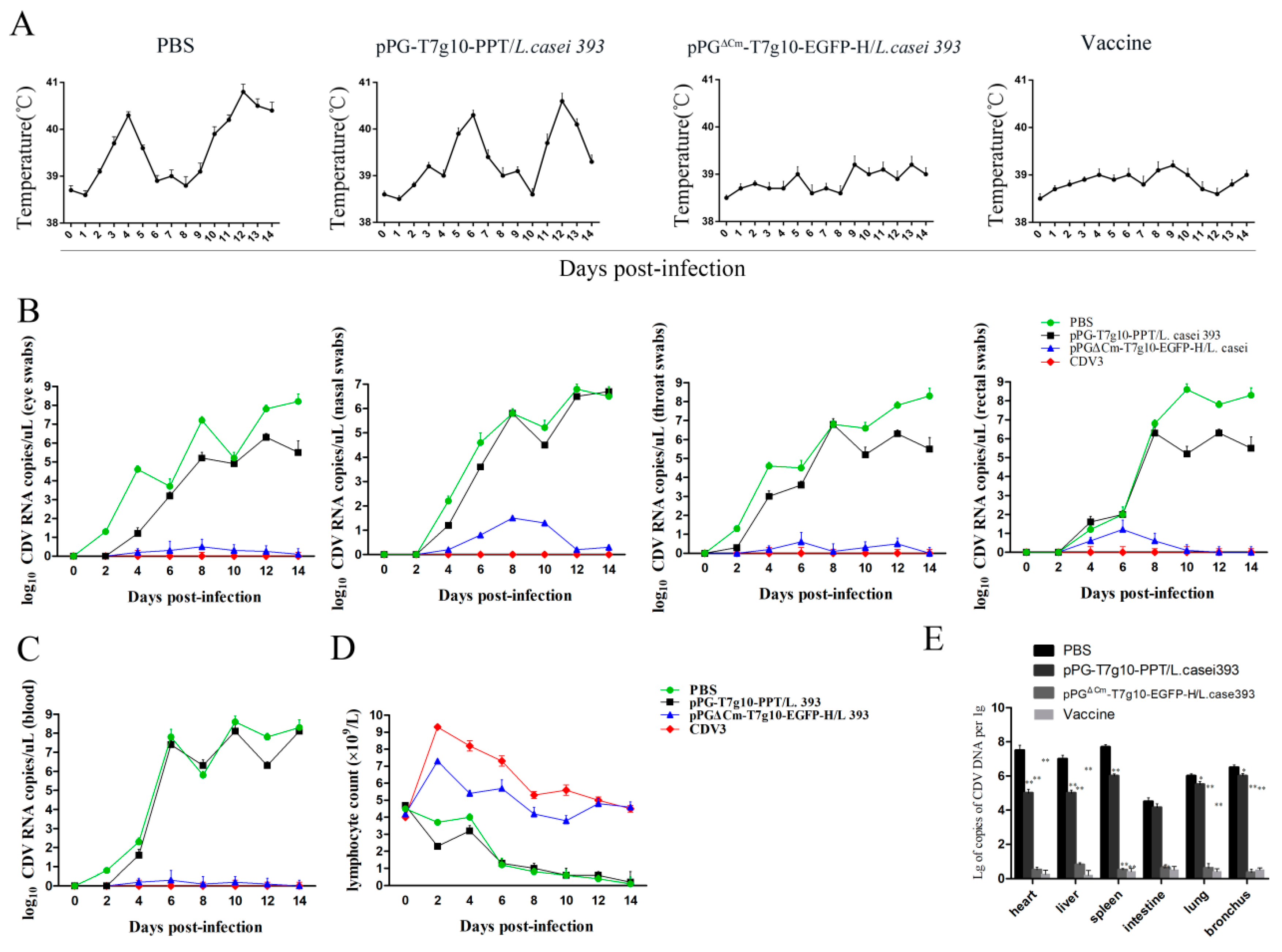
3.6. Protection Afforded by the Probiotic Vaccine against CDV Challengein Dogs
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Beineke, A.; Baumgartner, W.; Wohlsein, P. Cross-species transmission of canine distemper virus-an update. One Health 2015, 1, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Gutierrez, M.; Ruiz-Saenz, J. Diversity of susceptible hosts in canine distemper virus infection: A systematic review and data synthesis. BMC Vet. Res. 2016, 12, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, T. Morbillivirus infections, with special emphasis on morbilliviruses of carnivores. Vet. Microbiol. 1999, 69, 3–13. [Google Scholar] [CrossRef]
- Moss, W.J.; Griffin, D.E. Global measles elimination. Nat. Rev. Microbiol. 2006, 4, 900. [Google Scholar] [CrossRef] [PubMed]
- von Messling, V.; Springfeld, C.; Devaux, P.; Cattaneo, R. A ferret model of canine distemper virus virulence and immunosuppression. J. Virol. 2003, 77, 12579–12591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ek-Kommonen, C.; Sihvonen, L.; Pekkanen, K.; Rikula, U.; Nuotio, L. Outbreak off canine distemper in vaccinated dogs in Finland. Vet. Rec. 1997, 141, 380–383. [Google Scholar] [CrossRef] [PubMed]
- von Messling, V.; Svitek, N.; Cattaneo, R. Receptor (SLAM CD150) recognition and the V protein sustain swift lymphocyte-based invasion of mucosal tissue and lymphatic organs by a morbillivirus. J. Virol. 2006, 80, 6084–6092. [Google Scholar] [CrossRef] [Green Version]
- Wenzlow, N.; Plattet, P.; Wittek, R.; Zurbriggen, A.; Grone, A. Immunohistochemical demonstration of the putative canine distemper virus receptor CD150 in dogs with and without distemper. Vet. Pathol. 2007, 44, 943–948. [Google Scholar] [CrossRef] [Green Version]
- Delpeut, S.; Noyce, R.S.; Richardson, C.D. The V domain of dog PVRL4 (nectin-4) mediates canine distemper virus entry and virus cell-to-cell spread. Virology 2014, 454, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Ludlow, M.; Rennick, L.J.; Nambulli, S.; de Swart, R.L.; Duprex, W.P. Using the ferret model to study morbillivirus entry, spread, transmission and cross-species infection. Curr. Opin. Virol. 2014, 4, 15–23. [Google Scholar] [CrossRef]
- von Messling, V.; Milosevic, D.; Cattaneo, R. Tropism illuminated: Lymphocyte-based pathways blazed by lethal morbillivirus through the host immune system. Proc. Natl. Acad. Sci. USA 2004, 101, 14216–14221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roethlisberger, A.; Wiener, D.; Schweizer, M.; Peterhans, E.; Zurbriggen, A.; Plattet, P. Two Domains of the V Protein of Virulent Canine Distemper Virus Selectively Inhibit STAT1 and STAT2 Nuclear Import. J. Virol. 2010, 84, 6328–6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svitek, N.; Gerhauser, I.; Goncalves, C.; Grabski, E.; Doering, M.; Kalinke, U.; Anderson, D.E.; Cattaneo, R.; von Messling, V. Morbillivirus Control of the Interferon Response: Relevance of STAT2 and mda5 but Not STAT1 for Canine Distemper Virus Virulence in Ferrets. J. Virol. 2014, 88, 2941–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blixenkrone-Moller, M.; Svansson, V.; Have, P.; Orvell, C.; Appel, M.; Pedersen, I.R.; Dietz, H.H.; Henriksen, P. Studies on manifestations of canine distemper virus infection in an urban dog population. Vet. Microbiol. 1993, 37, 163–173. [Google Scholar] [CrossRef]
- Martella, V.; Blixenkrone-Moller, M.; Elia, G.; Lucente, M.S.; Cirone, F.; Decaro, N.; Nielsen, L.; Banyai, K.; Carmichael, L.E.; Buonavoglia, C. Lights and shades on an historical vaccine canine distemper virus, the Rockborn strain. Vaccine 2011, 29, 1222–1227. [Google Scholar] [CrossRef]
- Liu, P.C.; Chen, C.A.; Chen, C.M.; Yen, C.H.; Lee, M.H.; Chuang, C.K.; Tu, C.F.; Su, B.L. Application of xenogeneic anti-canine distemper virus antibodies in treatment of canine distemper puppies. J. Small Anim. Pract. 2016, 57, 626–630. [Google Scholar] [CrossRef]
- Deem, S.L.; Spelman, L.H.; Yates, R.A.; Montali, R.J. Canine distemper in terrestrial carnivores: A review. J. Zoo Wildl. Med. 2000, 31, 441–451. [Google Scholar]
- Sawatsky, B.; von Messling, V. Canine Distemper Viruses Expressing a Hemagglutinin without N-Glycans Lose Virulence but Retain Immunosuppression. J. Virol. 2010, 84, 2753–2761. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-G.; Guan, X.-T.; Liu, Z.-M.; Tian, C.-Y.; Cui, L.-C. Immunogenicity in Swine of Orally Administered Recombinant Lactobacillus plantarum Expressing Classical Swine Fever Virus E2 Protein in Conjunction with Thymosin alpha-1 as an Adjuvant. Appl. Environ. Microbiol. 2015, 81, 3745–3752. [Google Scholar] [CrossRef] [Green Version]
- Xin, K.Q.; Hoshino, Y.; Toda, Y.; Igimi, S.; Kojima, Y.; Jounai, N.; Ohba, K.; Kushiro, A.; Kiwaki, M.; Hamajima, K.; et al. Immunogenicity and protective efficacy of orally administered recombinant Lactococcus lactis expressing surface-bound HIV. Environ. Blood 2003, 102, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Mannam, P.; Jones, K.F.; Geller, B.L. Mucosal vaccine made from live, recombinant Lactococcus lactis protects mice against pharyngeal infection with Streptococcus pyogenes. Infect. Immun. 2004, 72, 3444–3450. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, L.; Huang, X.; Ma, S.; Yu, M.; Shi, W.; Qiao, X.; Tang, L.; Xu, Y.; Li, Y. Oral Delivery of Probiotics Expressing Dendritic Cell-Targeting Peptide Fused with Porcine Epidemic Diarrhea Virus COE Antigen: A Promising Vaccine Strategy against PEDV. Viruses 2017, 9, 312. [Google Scholar] [CrossRef]
- Gao, X.; Ma, Y.; Wang, Z.; Bai, J.; Jia, S.; Feng, B.; Jiang, Y.; Cui, W.; Tang, L.; Li, Y.; et al. Oral immunization of mice with a probiotic Lactobacillus casei constitutively expressing the alpha-toxoid induces protective immunity against Clostridium perfringens alpha-toxin. Virulence 2019, 10, 166–179. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Feng, B.; Niu, C.; Jia, S.; Sun, C.; Wang, Z.; Jiang, Y.; Cui, W.; Wang, L.; Xu, Y. Dendritic Cell Targeting of Bovine Viral Diarrhea Virus E2 Protein Expressed by Lactobacillus casei Effectively Induces Antigen-Specific Immune Responses via Oral Vaccination. Viruses 2019, 11, 575. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Qi, R.; Chen, C.; Yin, J.; Ma, S.; Shi, W.; Wu, Y.; Ge, J.; Jiang, Y.; Tang, L.; et al. Immunogenicity of recombinant Lactobacillus casei-expressing F4 (K88) fimbrial adhesin FaeG in conjunction with a heat-labile enterotoxin A (LTAK63) and heat-labile enterotoxin B (LTB) of enterotoxigenic Escherichia coli as an oral adjuvant in mice. J. Appl. Microbiol. 2017, 122, 506–515. [Google Scholar] [CrossRef]
- Marteau, P.; Rambaud, J.C. Potential of using lactic acid bacteria for therapy and immunomodulation in man. FEMS Microbiol. Rev. 1993, 12, 207–220. [Google Scholar] [CrossRef]
- Lee, J.S.; Shin, K.S.; Pan, J.G.; Kim, C.J. Surface-displayed viral antigens on Salmonella carrier vaccine. Nat. Biotechnol. 2000, 18, 645–648. [Google Scholar] [CrossRef]
- Shata, M.T.; Hone, D.M. Vaccination with a Shigella DNA vaccine vector induces antigen-specific CD8(+) T cells and antiviral protective immunity. J. Virol. 2001, 75, 9665–9670. [Google Scholar] [CrossRef] [Green Version]
- Shaw, D.M.; Gaerthe, B.; Leer, R.J.; Van Der Stap, J.G.; Smittenaar, C.; Heijne Den Bak-Glashouwer, M.; Thole, J.E.; Tielen, F.J.; Pouwels, P.H.; Havenith, C.E. Engineering the microflora to vaccinate the mucosa: Serum immunoglobulin G responses and activated draining cervical lymph nodes following mucosal application of tetanus toxin fragment C-expressing lactobacilli. Immunology 2000, 100, 510–518. [Google Scholar] [CrossRef]
- Christensen, H.R.; Frokiaer, H.; Pestka, J.J. Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. J. Immunol. 2002, 168, 171–178. [Google Scholar] [CrossRef]
- del Rio, B.; Dattwyler, R.J.; Aroso, M.; Neves, V.; Meirelles, L.; Seegers, J.F.M.L.; Gomes-Solecki, M. Oral immunization with recombinant Lactobacillus plantarum induces a protective immune response in mice with Lyme disease. Clin. Vaccine Immunol. 2008, 15, 1429–1435. [Google Scholar] [CrossRef] [Green Version]
- Brisbin, J.T.; Gong, J.; Parvizi, P.; Sharif, S. Effects of Lactobacilli on Cytokine Expression by Chicken Spleen and Cecal Tonsil Cells. Clin. Vaccine Immunol. 2010, 17, 1337–1343. [Google Scholar] [CrossRef] [Green Version]
- Vintini, E.; Villena, J.; Alvarez, S.; Medina, M. Administration of a probiotic associated with nasal vaccination with inactivated Lactococcus lactis-PppA induces effective protection against pneumoccocal infection in young mice. Clin. Exp. Immunol. 2010, 159, 351–362. [Google Scholar] [CrossRef]
- Song, B.-F.; Ju, L.-Z.; Li, Y.-J.; Tang, L.-J. Chromosomal Insertions in the Lactobacillus casei upp Gene That Are Useful for Vaccine Expression. Appl. Environ. Microbiol. 2014, 80, 3321–3326. [Google Scholar] [CrossRef] [Green Version]
- Ashiuchi, M.; Nawa, C.; Kamei, T.; Song, J.J.; Hong, S.P.; Sung, M.H.; Soda, K.; Misono, H. Physiological and biochemical characteristics of poly gamma-glutamate synthetase complex of Bacillus subtilis. Eur. J. Biochem. 2001, 268, 5321–5328. [Google Scholar] [CrossRef]
- Olins, P.O.; Rangwala, S.H. A novel sequence element derived from bacteriophage T7 mRNA acts as an enhancer of translation of the lacZ gene in Escherichia coli. J. Biol. Chem. 1989, 264, 16973–16976. [Google Scholar]
- Ashiuchi, M.; Soda, K.; Misono, H. A poly-gamma-glutamate synthetic system of Bacillus subtilis IFO 3336: Gene cloning and biochemical analysis of poly-gamma-glutamate produced by Escherichia coli clone cells. Biochem. Biophys. Res. Commun. 1999, 263, 6–12. [Google Scholar] [CrossRef]
- Ge, J.-W.; Liu, D.-Q.; Li, Y.-J. Construction of recombinant lactobacilli expressing the core neutralizing epitope (COE) of porcine epidemic diarrhea virus and a fusion protein consisting of COE and Escherichia coli heat-labile enterotoxin B, and comparison of the immune responses by orogastric immunization. Can. J. Microbiol. 2012, 58, 1258–1267. [Google Scholar]
- Yu, M.; Wang, L.; Ma, S.; Wang, X.; Wang, Y.; Xiao, Y.; Jiang, Y.; Qiao, X.; Tang, L.; Xu, Y.; et al. Immunogenicity of eGFP-Marked Recombinant Lactobacillus casei against Transmissible Gastroenteritis Virus and Porcine Epidemic Diarrhea Virus. Viruses 2017, 9, 274. [Google Scholar] [CrossRef] [Green Version]
- Stephensen, C.B.; Welter, J.; Thaker, S.R.; Taylor, J.; Tartaglia, J.; Paoletti, E. Canine distemper virus (CDV) infection of ferrets as a model for testing Morbillivirus vaccine strategies: NYVAC- and ALVAC-based CDV recombinants protect against symptomatic infection. J. Virol. 1997, 71, 1506–1513. [Google Scholar]
- Iwatsuki, K.; Tokiyoshi, S.; Hirayama, N.; Nakamura, K.; Ohashi, K.; Wakasa, C.; Mikami, T.; Kai, C. Antigenic differences in the H proteins of canine distemper viruses. Vet. Microbiol. 2000, 71, 281–286. [Google Scholar] [CrossRef]
- Bi, Z.; Xia, X.; Wang, Y.; Mei, Y. Development and characterization of neutralizing monoclonal antibodies against canine distemper virus hemagglutinin protein. Microbiol. Immunol. 2015, 59, 202–208. [Google Scholar] [CrossRef]
- Bi, Z.W.; Wang, Y.S.; Wang, X.L.; Xia, X.X. Phylogenetic analysis of canine distemper virus in domestic dogs in Nanjing, China. Arch. Virol. 2015, 160, 523–527. [Google Scholar] [CrossRef]
- Bronson, E.; Deem, S.L.; Sanchez, C.; Murray, S. Serologic response to a canarypox-vectored canine distemper virus vaccine in the giant panda (Ailuropoda melanoleuca). J. Zoo Wildl. Med. 2007, 38, 363–366. [Google Scholar] [CrossRef]
- Coke, R.L.; Backues, K.A.; Hoover, J.P.; Saliki, J.T.; Ritchey, J.W.; West, G.D. Serologic responses after vaccination of fennec foxes (Vulpes zerda) and meerkats (Suricata suricatta) with a live, canarypox-vectored canine distemper virus vaccine. J. Zoo Wildl. Med. 2005, 36, 326–331. [Google Scholar] [CrossRef]
- Alimolaei, M.; Golchin, M.; Daneshvar, H. Oral immunization of mice against Clostridium perfringens epsilon toxin with a Lactobacillus casei vector vaccine expressing epsilon toxoid. Infect. Genet. Evol. 2016, 40, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Kourkoutas, Y.; Bosnea, L.; Taboukos, S.; Baras, C.; Lambrou, D.; Kanellaki, M. Probiotic cheese production using Lactobacillus casei cells immobilized on fruit pieces. J. Dairy Sci. 2006, 89, 1439–1451. [Google Scholar] [CrossRef]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; Aindelis, G.; Tompoulidou, E.; Lamprianidou, E.E.; Saxami, G.; Ypsilantis, P.; Lampri, E.S.; Simopoulos, C.; et al. Lactobacillus casei Exerts Anti-Proliferative Effects Accompanied by Apoptotic Cell Death and Up-Regulation of TRAIL in Colon Carcinoma Cells. PLoS ONE 2016, 11, e0147960. [Google Scholar] [CrossRef]
- Sidira, M.; Galanis, A.; Ypsilantis, P.; Karapetsas, A.; Progaki, Z.; Simopoulos, C.; Kourkoutas, Y. Effect of Probiotic-Fermented Milk Administration on Gastrointestinal Survival of Lactobacillus casei ATCC 393 and Modulation of Intestinal Microbial Flora. J. Mol. Microbiol. Biotechnol. 2010, 19, 224–230. [Google Scholar] [CrossRef]
- Mohamadzadeh, M.; Duong, T.; Sandwick, S.J.; Hoover, T.; Klaenhammer, T.R. Dendritic cell targeting of Bacillus anthracis protective antigen expressed by Lactobacillus acidophilus protects mice from lethal challenge. Proc. Natl. Acad. Sci. USA 2009, 106, 4331–4336. [Google Scholar] [CrossRef] [Green Version]
- Sim, A.C.N.; Lin, W.; Tan, G.K.X.; Sim, M.S.T.; Chow, V.T.K.; Alonso, S. Induction of neutralizing antibodies against dengue virus type 2 upon mucosal administration of a recombinant Lactococcus lactis strain expressing envelope domain III antigen. Vaccine 2008, 26, 1145–1154. [Google Scholar] [CrossRef]
- Liew, F.Y.; McInnes, I.B. The role of innate mediators in inflammatory response. Mol. Immunol. 2002, 38, 887–890. [Google Scholar] [CrossRef]
- Basso, A.S.; Cheroutre, H.; Mucida, D. More stories on Th17 cells. Cell Res. 2009, 19, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Romagnani, S.; Maggi, E.; Liotta, F.; Cosmi, L.; Annunziato, F. Properties and origin of human Th17 cells. Mol. Immunol. 2009, 47, 3–7. [Google Scholar] [CrossRef]
- Boniface, K.; Blom, B.; Liu, Y.-J.; Malefyt, R.W. From interleukin-23 to T-helper 17 cells: Human T-helper cell differentiation revisited. Immunol. Rev. 2008, 226, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11, 45–53. [Google Scholar] [CrossRef]
- Neutra, M.R.; Kozlowski, P.A. Mucosal vaccines: The promise and the challenge. Nat. Rev. Immunol. 2006, 6, 148–158. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primers | Sequences (5′–3′) | Application |
|---|---|---|---|
| H | H1 | GAGGAGCTC1CTCGGTACCATGCTCTCCTACCAAGACAAG | For lactic acid bacteria expression |
| H2 | CTTGGGCCC1TCAAGGTTTTGAACGGTTACATGAG | ||
| H3 | CCAGGATCC1ATGGTTGGGGTCAAAAAATCTAT | For E. coli expression | |
| H4 | CTTGTCGAC1TCAAGGTTTTGAACGGTTACATGAG | ||
| H5 | TGACAGCAACGGTTCACAAGATGG | For qRT-PCR | |
| H6 | CAGAGACCAATACAGGCACCATCC | ||
| EGFP | E1 | ATGGTGAGCAAGGG | For amplification of EGFP |
| E2 | TCACTTGTACAGCTCGTC |
| Group Pathway | Dose | |
|---|---|---|
| PBS | Oral | 200 μL |
| Intranasal | 20 μL | |
| pPG-T7g10-PPT/L. casei 393 | Oral | 1010 CFU/200 μL |
| Intranasal | 1010 CFU/20 μL | |
| pPGΔCm-T7g10-EGFP-H/L. casei 393 | Oral | 1010 CFU/200 μL |
| Intranasal | 1010 CFU/20 μL | |
| Group | Immune Route | No. | Deaths | Survival | Cumulative Mortality |
|---|---|---|---|---|---|
| PBS | Intranasal | 8 | 8 | 0 | 100% |
| pPG-T7g10-PPT/L. casei 393 | Intranasal | 8 | 8 | 0 | 100% |
| pPGΔCm-T7g10-EGFP-H/L. casei 393 | Intranasal | 8 | 0 | 8 | 0% |
| Vaccine | Intranasal | 8 | 0 | 8 | 0% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Jia, S.; Zheng, D.; Li, F.; Wang, S.; Wang, L.; Qiao, X.; Cui, W.; Tang, L.; Xu, Y.; et al. Protective Immunity against Canine Distemper Virus in Dogs Induced by Intranasal Immunization with a Recombinant Probiotic Expressing the Viral H Protein. Vaccines 2019, 7, 213. https://doi.org/10.3390/vaccines7040213
Jiang Y, Jia S, Zheng D, Li F, Wang S, Wang L, Qiao X, Cui W, Tang L, Xu Y, et al. Protective Immunity against Canine Distemper Virus in Dogs Induced by Intranasal Immunization with a Recombinant Probiotic Expressing the Viral H Protein. Vaccines. 2019; 7(4):213. https://doi.org/10.3390/vaccines7040213
Chicago/Turabian StyleJiang, Yanping, Shuo Jia, Dianzhong Zheng, Fengsai Li, Shengwen Wang, Li Wang, Xinyuan Qiao, Wen Cui, Lijie Tang, Yigang Xu, and et al. 2019. "Protective Immunity against Canine Distemper Virus in Dogs Induced by Intranasal Immunization with a Recombinant Probiotic Expressing the Viral H Protein" Vaccines 7, no. 4: 213. https://doi.org/10.3390/vaccines7040213
APA StyleJiang, Y., Jia, S., Zheng, D., Li, F., Wang, S., Wang, L., Qiao, X., Cui, W., Tang, L., Xu, Y., Xia, X., & Li, Y. (2019). Protective Immunity against Canine Distemper Virus in Dogs Induced by Intranasal Immunization with a Recombinant Probiotic Expressing the Viral H Protein. Vaccines, 7(4), 213. https://doi.org/10.3390/vaccines7040213