Low-Energy Electron Irradiation Efficiently Inactivates the Gram-Negative Pathogen Rodentibacter pneumotropicus—A New Method for the Generation of Bacterial Vaccines with Increased Efficacy

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Cultivation of Rodentibacter Pneumotropicus
2.3. Pathogen Inactivation
2.3.1. Low-energy Electron Irradiation (LEEI)
2.3.2. Formaldehyde Inactivation
2.4. ELISA
2.5. LPS Based Reporter Assay
2.6. Immunization and Challenge
2.7. Determination of Bacterial Load
2.8. Statistical Analysis
3. Results
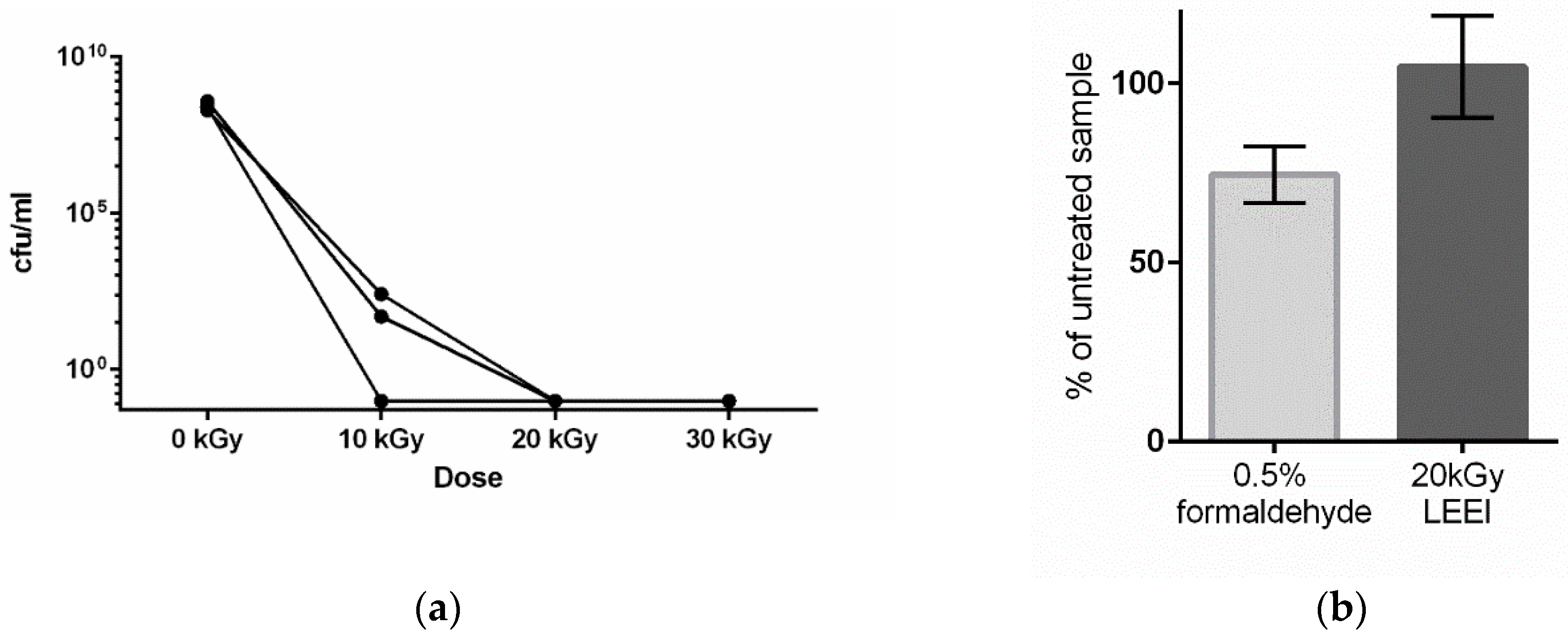
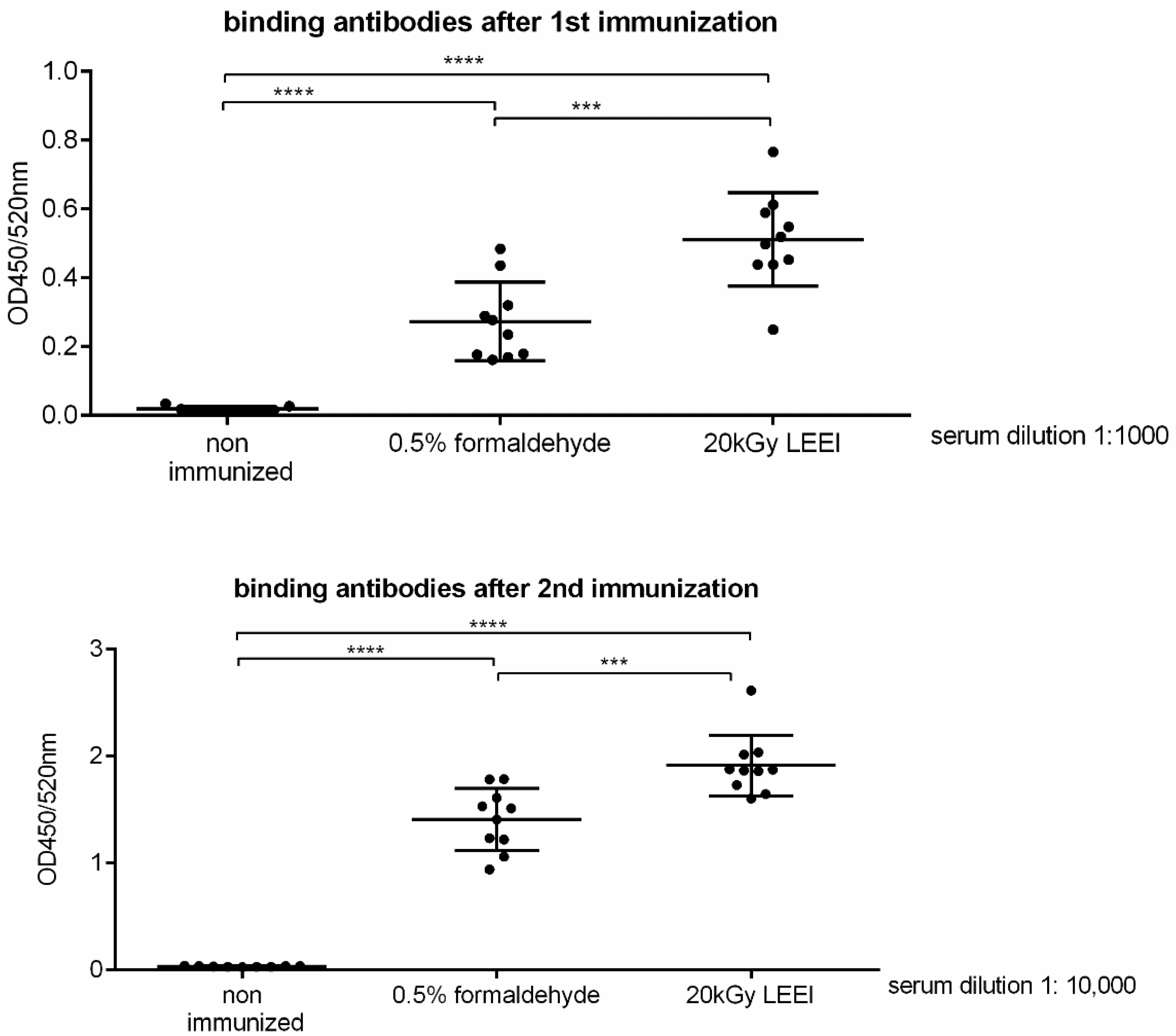
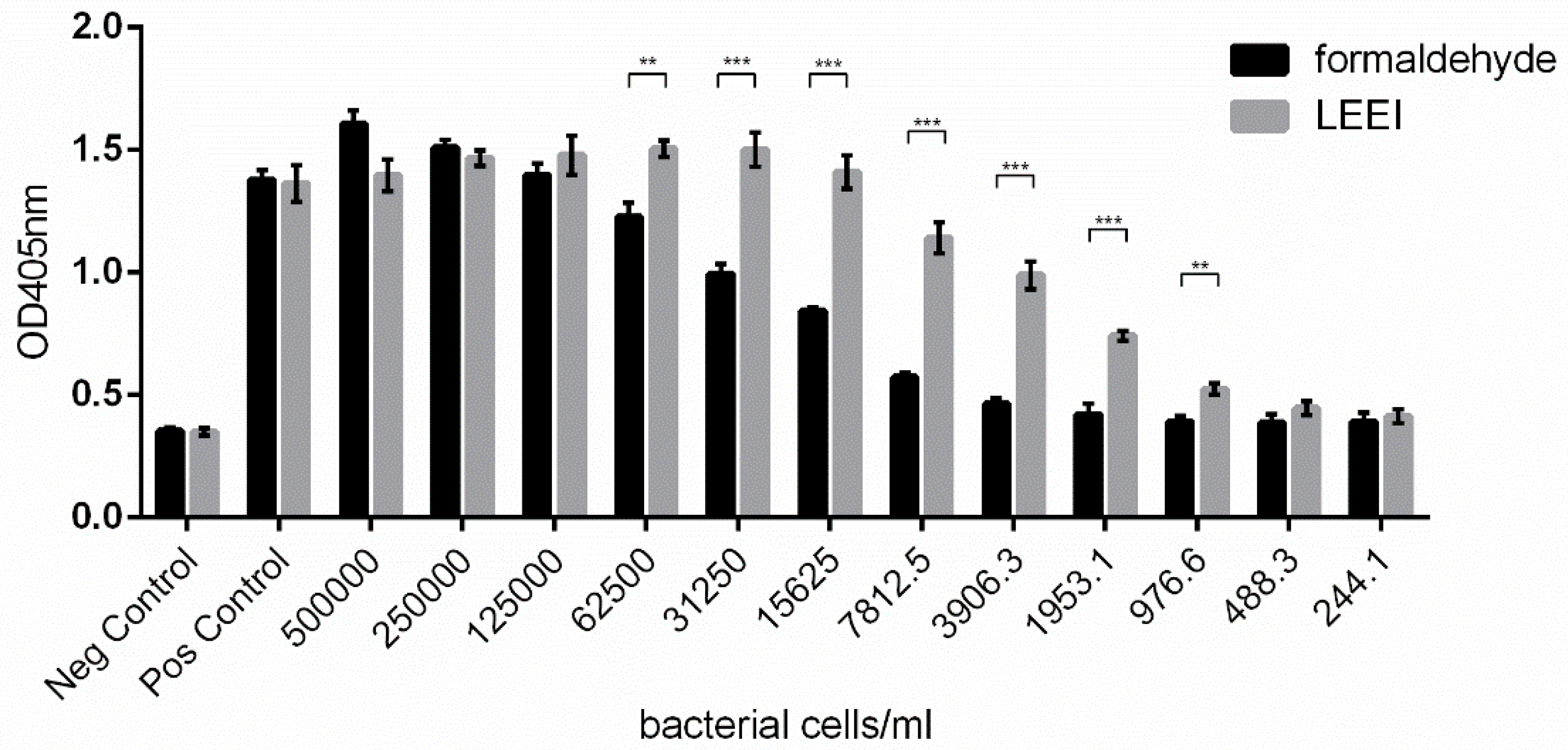
3.1. Low-Energy Electron Irradiation Inactivates Bacteria with High Antigen Conservation
3.2. Low-Energy Electron Irradiation Elicits Strong Immune Responses after Vaccination of Mice
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldstein, M.A.; Tauraso, N.M. Effect of formalin, beta-propiolactone, merthiolate, and ultraviolet light upon influenza virus infectivity chicken cell agglutination, hemagglutination, and antigenicity. Appl. Microbiol. 1970, 19, 290–294. [Google Scholar] [CrossRef] [Green Version]
- Amanna, I.J.; Raué, H.-P.; Slifka, M.K. Development of a new hydrogen peroxide–based vaccine platform. Nat. Med. 2012, 18, 974–979. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.S. Application of radiation technology in vaccines development. Clin. Exp. Vaccine Res. 2015, 4, 145. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.-C.; Chiu, H.-C.; Chen, L.-K.; Chang, G.-J.J.; Chiou, S.-S. Formalin Inactivation of Japanese Encephalitis Virus Vaccine Alters the Antigenicity and Immunogenicity of a Neutralization Epitope in Envelope Protein Domain III. PLoS Negl. Trop. Dis. 2015, 9, e0004167. [Google Scholar] [CrossRef] [Green Version]
- Brown, F. Formaldehyde as an inactivant. Vaccine 1995, 13, 231. [Google Scholar] [CrossRef]
- Ferguson, M.; Wood, D.J.; Minor, P.D. Antigenic structure of poliovirus in inactivated vaccines. J. Gen. Virol. 1993, 74, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Babb, R.; Chen, A.; Hirst, T.R.; Kara, E.E.; McColl, S.R.; Ogunniyi, A.D.; Paton, J.C.; Alsharifi, M. Intranasal vaccination with γ-irradiated Streptococcus pneumoniae whole-cell vaccine provides serotype-independent protection mediated by B-cells and innate IL-17 responses. Clin. Sci. 2016, 130, 697–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabral, N.; Martha-Moreno-Lafont, N.S.; Vemulapalli, R. Oral immunization of mice with gamma-irradiated Brucella neotomae induces protection against intraperitoneal and intranasal challenge with virulent B. abortus 2308. PLoS ONE 2014, 9, e107180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordin, A.I.; Pillai, S.D.; Brake, C.; Bagley, K.B.; Bourquin, J.R.; Coleman, M.; Oliveira, F.N.; Mwangi, W.; McMurray, D.N.; Love, C.C.; et al. Immunogenicity of an Electron Beam Inactivated Rhodococcus equi Vaccine in Neonatal Foals. PLoS ONE 2014, 9, e105367. [Google Scholar] [CrossRef] [PubMed]
- Jwa, M.Y.; Jeong, S.; Ko, E.B.; Kim, A.R.; Kim, H.Y.; Kim, S.K.; Seo, H.S.; Yun, C.-H.; Han, S.H. Gamma-irradiation of Streptococcus pneumoniae for the use as an immunogenic whole cell vaccine. J. Microbiol. 2018, 56, 579–585. [Google Scholar] [CrossRef]
- Alizadeh, E.; Orlando, T.M.; Sanche, L. Biomolecular damage induced by ionizing radiation: The direct and indirect effects of low-energy electrons on DNA. Annu. Rev. Phys. Chem. 2015, 66, 379–398. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, F. Chemical Changes Induced in DNA by Ionizing Radiation. In Progress in Nucleic Acid Research and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 1985; Volume 32, pp. 115–154. ISBN 9780125400329. [Google Scholar]
- Bayer, L.; Fertey, J.; Ulbert, S.; Grunwald, T. Immunization with an adjuvanted low-energy electron irradiation inactivated respiratory syncytial virus vaccine shows immunoprotective activity in mice. Vaccine 2018, 36, 1561–1569. [Google Scholar] [CrossRef]
- Delrue, I.; Verzele, D.; Madder, A.; Nauwynck, H.J. Inactivated virus vaccines from chemistry to prophylaxis: Merits, risks and challenges. Expert Rev. Vaccines 2012, 11, 695–719. [Google Scholar] [CrossRef] [Green Version]
- Silindir, M.; Özer, A.Y. Sterilization Methods and the Comparison of E-Beam Sterilization with Gamma Radiation Sterilization. FABAD J. Pharm. Sci. 2009, 34, 43–53. [Google Scholar]
- IAEA. IAEA Safety standards. In Radiation Safety of Gamma, Electron and X-ray Irradiation Facilities; IAEA: Vienna, Austria, 2010; ISBN 978-92-0-103710-7. [Google Scholar]
- Fertey, J.; Bayer, L.; Grunwald, T.; Pohl, A.; Beckmann, J.; Gotzmann, G.; Casado, J.; Schönfelder, J.; Rögner, F.-H.; Wetzel, C.; et al. Pathogens Inactivated by Low-Energy-Electron Irradiation Maintain Antigenic Properties and Induce Protective Immune Responses. Viruses 2016, 8, 319. [Google Scholar] [CrossRef] [PubMed]
- Thabet, A.; Schmäschke, R.; Fertey, J.; Bangoura, B.; Schönfelder, J.; Lendner, M.; Ulbert, S.; Daugschies, A. Eimeria tenella oocysts attenuated by low energy electron irradiation (LEEI) induce protection against challenge infection in chickens. Vet. Parasitol. 2019, 266, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Fornefett, J.; Krause, J.; Klose, K.; Fingas, F.; Hassert, R.; Benga, L.; Grunwald, T.; Müller, U.; Schrödl, W.; Baums, C.G. Comparative analysis of humoral immune responses and pathologies of BALB/c and C57BL/6 wildtype mice experimentally infected with a highly virulent Rodentibacter pneumotropicus (Pasteurella pneumotropica) strain. BMC Microbiol. 2018, 18, 314. [Google Scholar] [CrossRef] [PubMed]
- Benga, L.; Benten, W.P.M.; Engelhardt, E.; Bleich, A.; Gougoula, C.; Sager, M. Development of a multiplex PCR assay based on the 16S–23S rRNA internal transcribed spacer for the detection and identification of rodent Pasteurellaceae. J. Microbiol. Methods 2013, 95, 256–261. [Google Scholar] [CrossRef]
- Burger-Kentischer, A.; Abele, I.S.; Finkelmeier, D.; Wiesmüller, K.-H.; Rupp, S. A new cell-based innate immune receptor assay for the examination of receptor activity, ligand specificity, signalling pathways and the detection of pyrogens. J. Immunol. Methods 2010, 358, 93–103. [Google Scholar] [CrossRef]
- See, S.B.; Thomas, W.R. Protective anti-outer membrane protein immunity against Pasteurella pneumotropica infection of mice. Microbes Infect. 2013, 15, 470–479. [Google Scholar] [CrossRef]
- Scott, G.H.; McCaul, T.F.; Williams, J.C. Inactivation of Coxiella burnetii by gamma irradiation. J. Gen. Microbiol. 1989, 135, 3263–3270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanislavsky, E.S.; Makarenko, T.A.; Kholodkova, E.V.; Lugowski, C. R-form lipopolysaccharides (LPS) of Gram-negative bacteria as possible vaccine antigens. FEMS Immunol. Med. Microbiol. 1997, 18, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Zariri, A.; van der Ley, P. Biosynthetically engineered lipopolysaccharide as vaccine adjuvant. Expert Rev. Vaccines 2015, 14, 861–876. [Google Scholar] [CrossRef] [PubMed]
- Hieke, A.-S.C.; Pillai, S.D. Escherichia coli Cells Exposed to Lethal Doses of Electron Beam Irradiation Retain Their Ability to Propagate Bacteriophages and Are Metabolically Active. Front. Microbiol. 2018, 9, 2138. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fertey, J.; Bayer, L.; Kähl, S.; Haji, R.M.; Burger-Kentischer, A.; Thoma, M.; Standfest, B.; Schönfelder, J.; Portillo Casado, J.; Rögner, F.-H.; et al. Low-Energy Electron Irradiation Efficiently Inactivates the Gram-Negative Pathogen Rodentibacter pneumotropicus—A New Method for the Generation of Bacterial Vaccines with Increased Efficacy. Vaccines 2020, 8, 113. https://doi.org/10.3390/vaccines8010113
Fertey J, Bayer L, Kähl S, Haji RM, Burger-Kentischer A, Thoma M, Standfest B, Schönfelder J, Portillo Casado J, Rögner F-H, et al. Low-Energy Electron Irradiation Efficiently Inactivates the Gram-Negative Pathogen Rodentibacter pneumotropicus—A New Method for the Generation of Bacterial Vaccines with Increased Efficacy. Vaccines. 2020; 8(1):113. https://doi.org/10.3390/vaccines8010113
Chicago/Turabian StyleFertey, Jasmin, Lea Bayer, Sophie Kähl, Rukiya M. Haji, Anke Burger-Kentischer, Martin Thoma, Bastian Standfest, Jessy Schönfelder, Javier Portillo Casado, Frank-Holm Rögner, and et al. 2020. "Low-Energy Electron Irradiation Efficiently Inactivates the Gram-Negative Pathogen Rodentibacter pneumotropicus—A New Method for the Generation of Bacterial Vaccines with Increased Efficacy" Vaccines 8, no. 1: 113. https://doi.org/10.3390/vaccines8010113
APA StyleFertey, J., Bayer, L., Kähl, S., Haji, R. M., Burger-Kentischer, A., Thoma, M., Standfest, B., Schönfelder, J., Portillo Casado, J., Rögner, F.-H., Baums, C. G., Grunwald, T., & Ulbert, S. (2020). Low-Energy Electron Irradiation Efficiently Inactivates the Gram-Negative Pathogen Rodentibacter pneumotropicus—A New Method for the Generation of Bacterial Vaccines with Increased Efficacy. Vaccines, 8(1), 113. https://doi.org/10.3390/vaccines8010113