Toxoplasma gondii ADSL Knockout Provides Excellent Immune Protection against a Variety of Strains

Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Parasite Strains
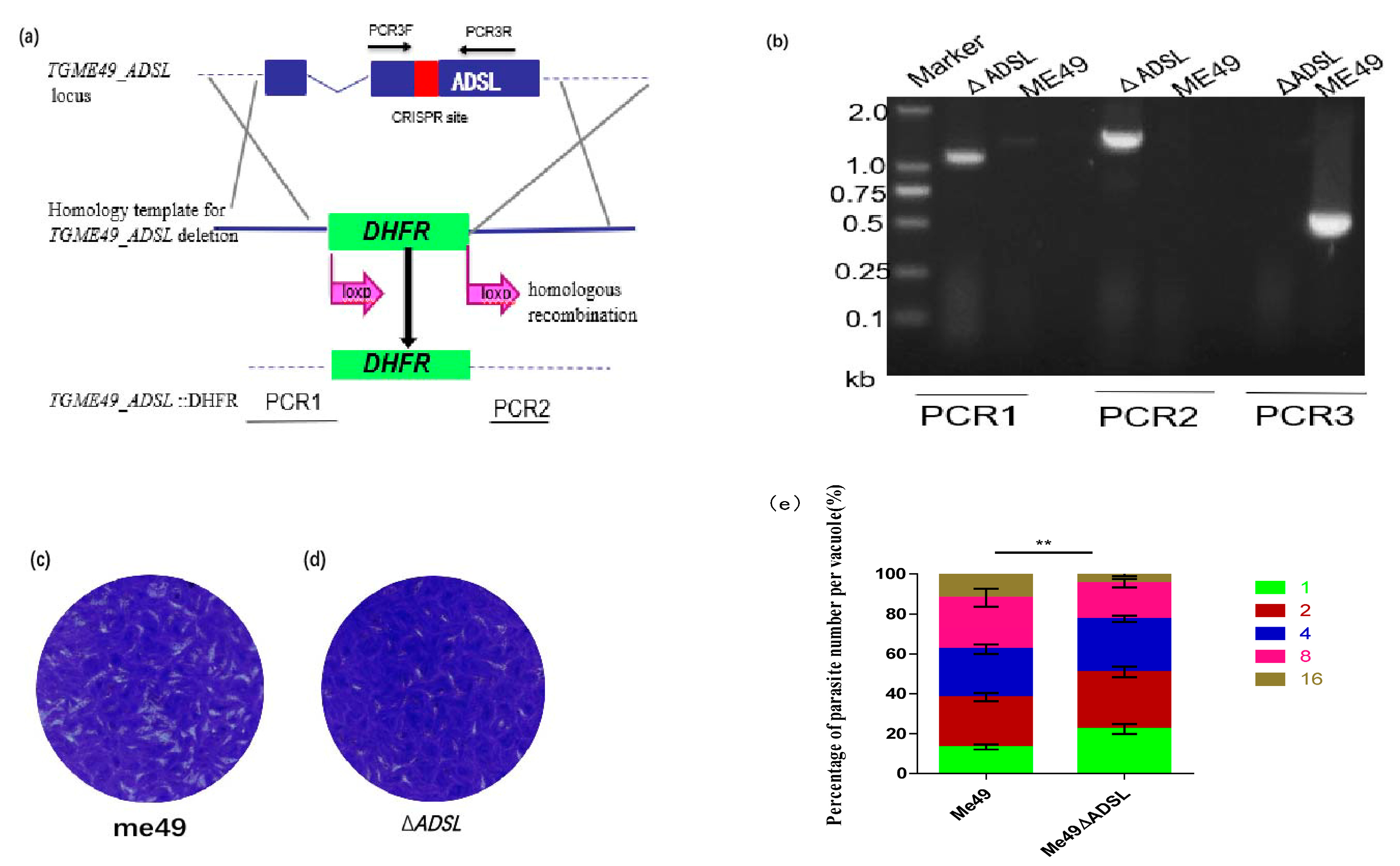
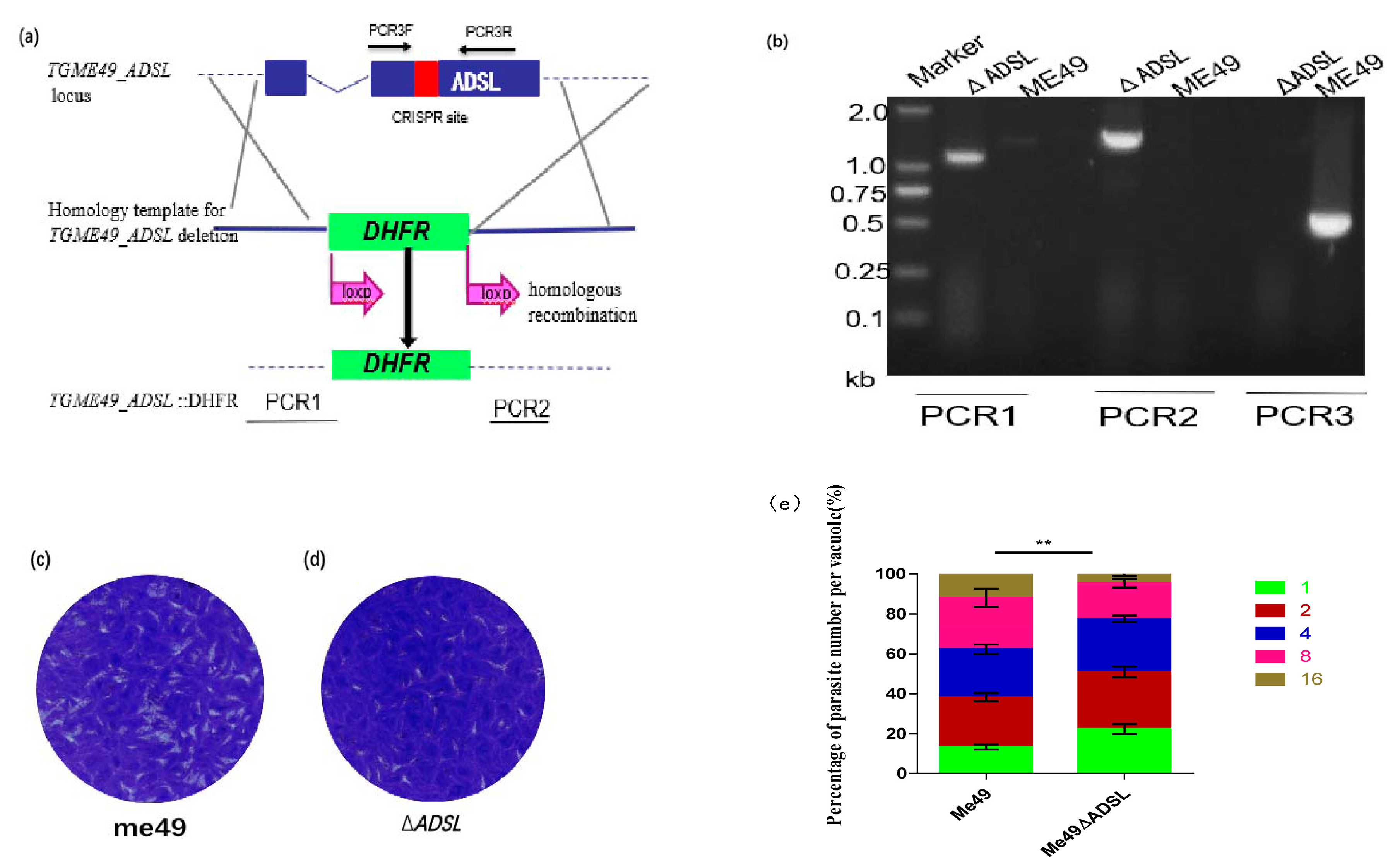
2.2. Disruption of ADSL by the CRISPR-Cas9 System
2.3. Virulence Tests and Cyst Formation of Mutants Versus WT Strains in Mice
2.4. Toxoplasma Plaque Assay
2.5. Assay for Parasite Replication In Vitro
2.6. Protection of the ME49ΔADSL Strain Against Acute Infection, Toxoplasma-Specific lgG levels and Cytokine Detection
2.7. Cytokine Production by Splenocytes After T. gondii Antigen Stimulation
2.8. Statistical Analysis
3. Results
3.1. Generation of ADSL-Deficient Type II T. gondii
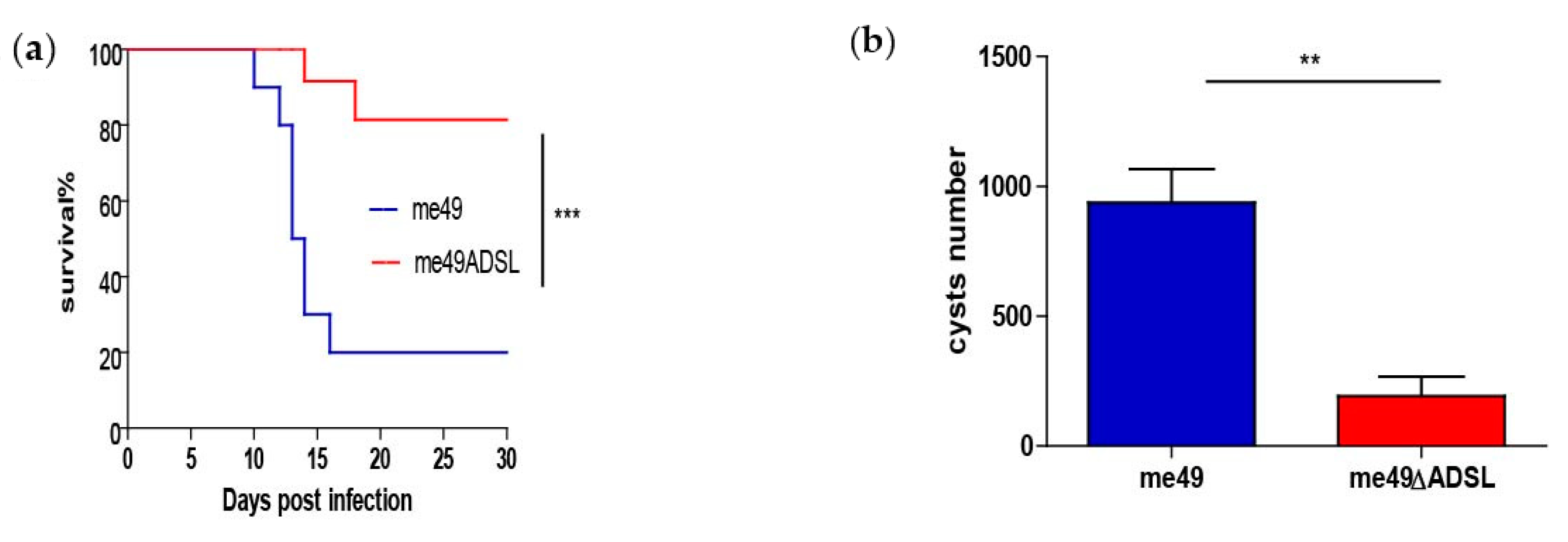
3.2. Effects of ADSL Mutation on Virulence and Reduced Cyst Formation In Vivo
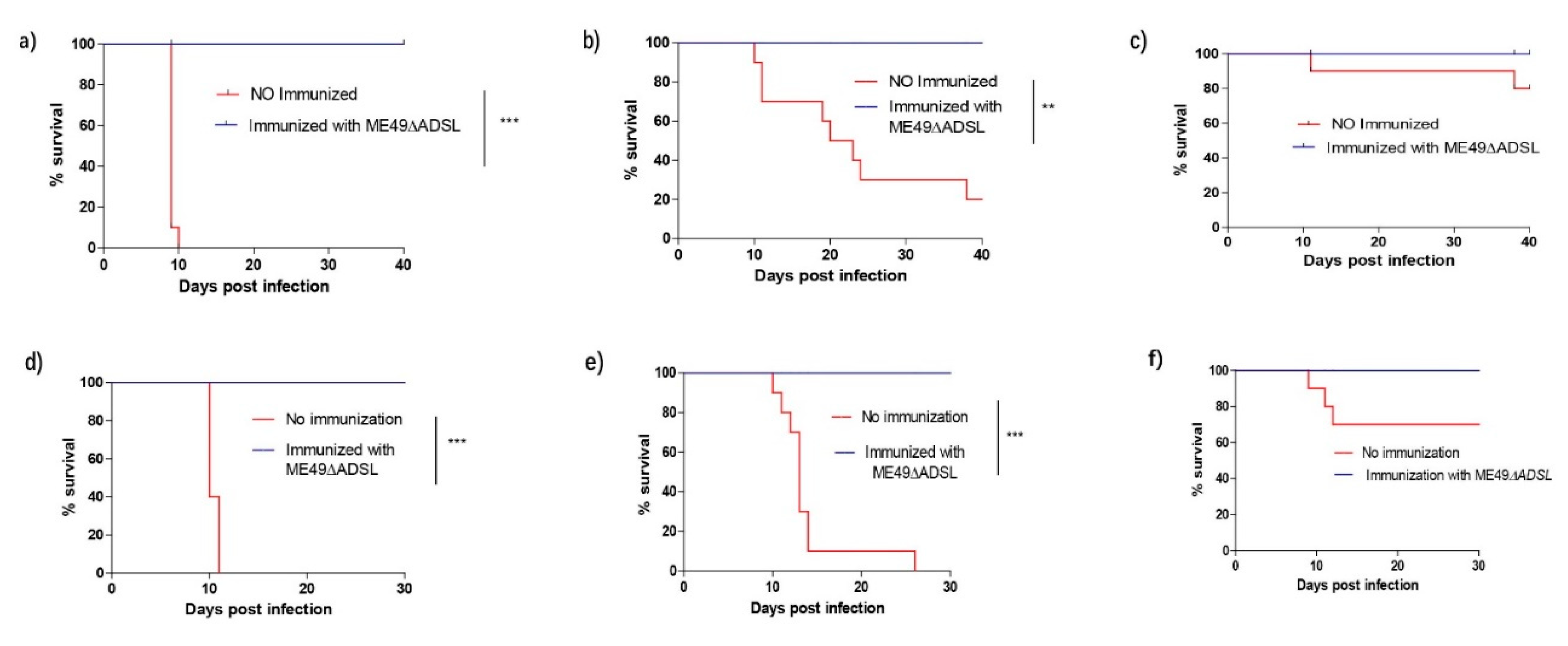
3.3. Immune Responses of Immunized Mice to Acute Tachyzoites Infection
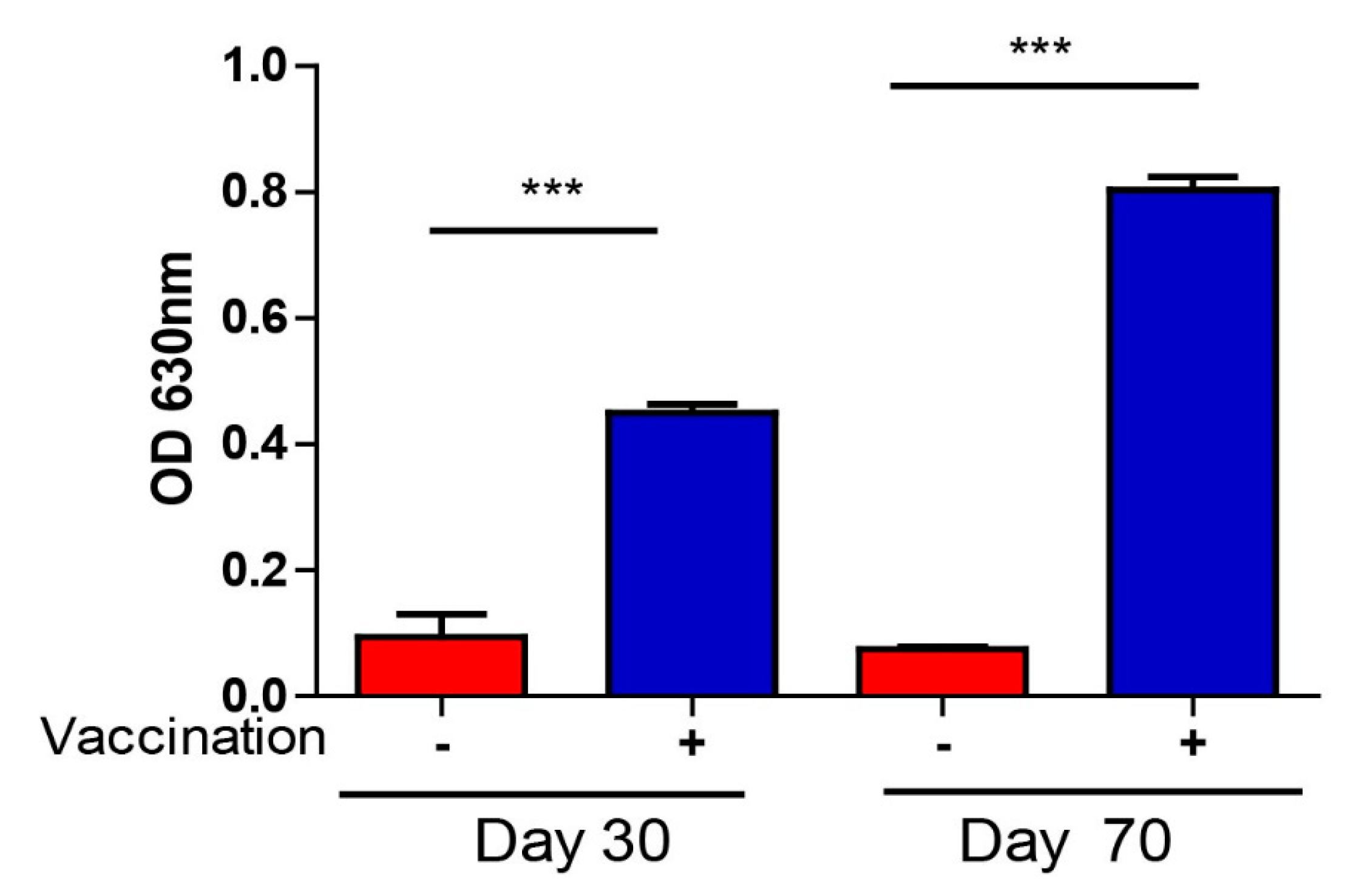
3.4. T. gondii-Specific IgG Antibodies and Cytokine Production After Vaccination
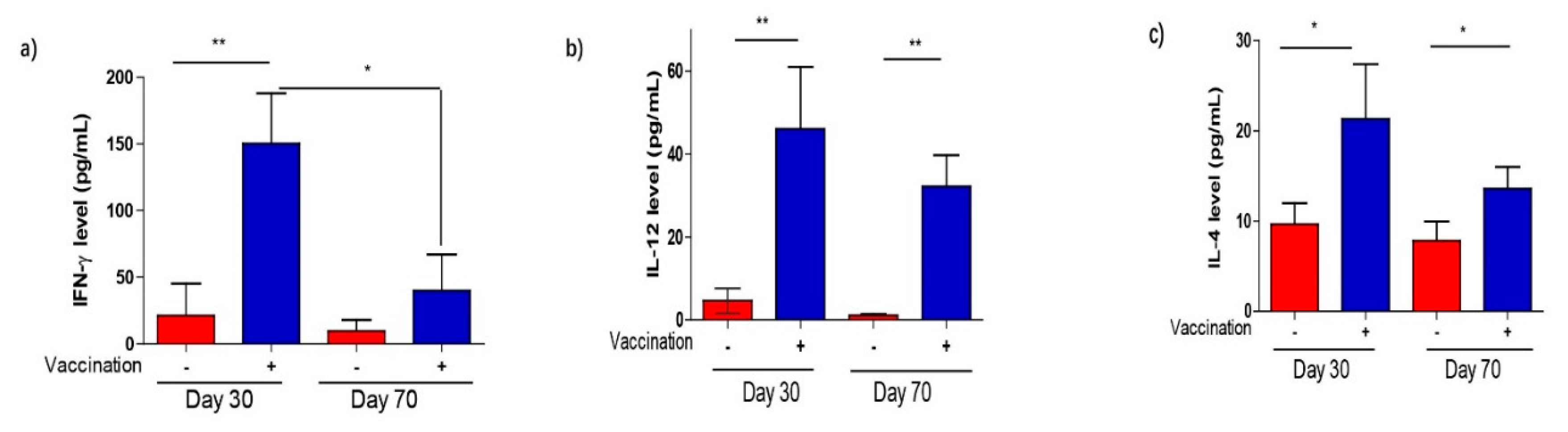
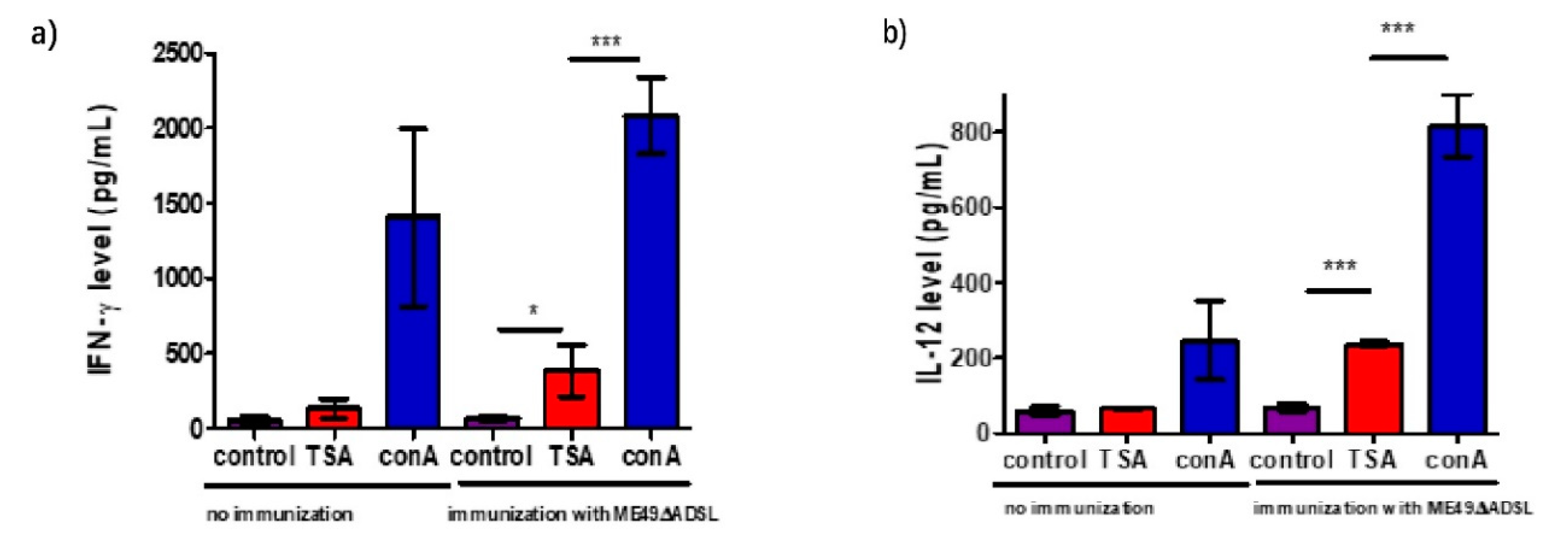
3.5. Cytokine Production After Vaccination in Response to Stimulation with T. gondii Antigen
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Dubey, J.P. The history of Toxoplasma gondii—The first 100 years. J. Eukaryot. Microbiol. 2008, 55, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Luft, B.J.; Conley, F.; Remington, J.S.; Laverdiere, M.; Wagner, K.F.; Levine, J.F.; Craven, P.C.; Strandberg, D.A.; File, T.M.; Rice, N.; et al. Outbreak of central-nervous-system toxoplasmosis in western Europe and North America. Lancet 1983, 1, 781–784. [Google Scholar] [CrossRef]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Remington, J.S. Toxoplasmosis in the adult. Bull. N. Y. Acad. Med. 1974, 50, 211–227. [Google Scholar]
- Roux, C.; Desmont, G.; Mulliez, N.; Gaulier, M.; Tufferaud, G.; Marmor, D.; Herbillon, A. Toxoplasmosis and pregnancy. Evaluation of 2 years of prevention of congenital toxoplasmosis in the maternity ward of Hopital Saint-Antoine (1973-1974). J. Gynecol. Obstet. Biol. Reprod. (Paris) 1976, 5, 249–264. [Google Scholar]
- Dubey, J.P.; Miller, S.; Powell, E.C.; Anderson, W.R. Epizootiologic investigations on a sheep farm with Toxoplasma gondii-induced abortions. J. Am. Vet. Med. Assoc. 1986, 188, 155–158. [Google Scholar]
- Dubey, J.P.; Welcome, F.L. Toxoplasma gondii-induced abortion in sheep. J. Am. Vet. Med. Assoc. 1988, 193, 697–700. [Google Scholar]
- Alday, P.H.; Doggett, J.S. Drugs in development for toxoplasmosis: Advances, challenges, and current status. Drug Des. Dev. Ther. 2017, 11, 273–293. [Google Scholar] [CrossRef] [Green Version]
- Buxton, D.; Innes, E.A. A commercial vaccine for ovine toxoplasmosis. Parasitology 1995, 110, S11–S16. [Google Scholar] [CrossRef]
- Wilkins, M.F.; O’Connell, E. Effect on lambing percentage of vaccinating ewes with Toxoplasma gondii. N. Z. Vet. J. 1983, 31, 181–182. [Google Scholar] [CrossRef]
- Daryani, A.; Hosseini, A.Z.; Dalimi, A. Immune responses against excreted/secreted antigens of Toxoplasma gondii tachyzoites in the murine model. Vet. Parasitol. 2003, 113, 123–134. [Google Scholar] [CrossRef]
- Waldeland, H.; Frenkel, J.K. Live and killed vaccines against toxoplasmosis in mice. J. Parasitol. 1983, 69, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Cao, A.; Liu, Y.; Wang, J.; Li, X.; Wang, S.; Zhao, Q.; Cong, H.; He, S.; Zhou, H. Toxoplasma gondii: Vaccination with a DNA vaccine encoding T- and B-cell epitopes of SAG1, GRA2, GRA7 and ROP16 elicits protection against acute toxoplasmosis in mice. Vaccine 2015, 33, 6757–6762. [Google Scholar] [CrossRef] [PubMed]
- Dautu, G.; Munyaka, B.; Carmen, G.; Zhang, G.; Omata, Y.; Xuenan, X.; Igarashi, M. Toxoplasma gondii: DNA vaccination with genes encoding antigens MIC2, M2AP, AMA1 and BAG1 and evaluation of their immunogenic potential. Exp. Parasitol. 2007, 116, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.D.; Chang, G.N.; Chao, D. Protective immunity against Toxoplasma gondii in mice induced by a chimeric protein rSAG1/2. Parasitol. Res. 2004, 92, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Bastos, L.M.; Macedo, A.G., Jr.; Silva, M.V.; Santiago, F.M.; Ramos, E.L.; Santos, F.A.; Pirovani, C.P.; Goulart, L.R.; Mineo, T.W.; Mineo, J.R. Toxoplasma gondii-Derived Synthetic Peptides Containing B- and T-Cell Epitopes from GRA2 Protein Are Able to Enhance Mice Survival in a Model of Experimental Toxoplasmosis. Front. Cell. Infect. Microbial. 2016, 6, 59. [Google Scholar] [CrossRef] [Green Version]
- Hajissa, K.; Zakaria, R.; Suppian, R.; Mohamed, Z. Epitope-based vaccine as a universal vaccination strategy against Toxoplasma gondii infection: A mini-review. J. Adv. Vet. Anim. Res. 2019, 6, 174–182. [Google Scholar] [CrossRef]
- Foroutan, M.; Ghaffarifar, F.; Sharifi, Z.; Dalimi, A.; Jorjani, O. Rhoptry antigens as Toxoplasma gondii vaccine target. Clin. Exp. Vaccine Res. 2019, 8, 4–26. [Google Scholar] [CrossRef]
- Gigley, J.P.; Fox, B.A.; Bzik, D.J. Long-term immunity to lethal acute or chronic type II Toxoplasma gondii infection is effectively induced in genetically susceptible C57BL/6 mice by immunization with an attenuated type I vaccine strain. Infect. Immun. 2009, 77, 5380–5388. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.L.; Elsheikha, H.M.; Zhu, W.N.; Chen, K.; Li, T.T.; Yue, D.M.; Zhang, X.X.; Huang, S.Y.; Zhu, X.Q. Immunization with Toxoplasma gondii GRA17 Deletion Mutant Induces Partial Protection and Survival in Challenged Mice. Front. Immunol. 2017, 8, 730. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.L.; Li, T.T.; Elsheikha, H.M.; Chen, K.; Cong, W.; Yang, W.B.; Bai, M.J.; Huang, S.Y.; Zhu, X.Q. Live Attenuated Pru: Deltacdpk2 Strain of Toxoplasma gondii Protects against Acute, Chronic, and Congenital Toxoplasmosis. J. Infect. Dis. 2018, 218, 768–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, N.; Zhou, T.; Liang, X.; Ye, S.; Zhao, P.; Yang, J.; Zhou, Y.; Zhao, J.; Shen, B. A Lactate Fermentation Mutant of Toxoplasma Stimulates Protective Immunity Against Acute and Chronic Toxoplasmosis. Front. Immunol. 2018, 9, 1814. [Google Scholar] [CrossRef] [Green Version]
- Krug, E.C.; Marr, J.J.; Berens, R.L. Purine metabolism in Toxoplasma gondii. J. Biol. Chem. 1989, 264, 10601–10607. [Google Scholar] [PubMed]
- Perrotto, J.; Keister, D.B.; Gelderman, A.H. Incorporation of precursors into Toxoplasma DNA. J. Protozool. 1971, 18, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, K.; Darling, J.A.; Fohl, L.M.; Sullivan, W.J., Jr.; Donald, R.G.; Pfefferkorn, E.R.; Ullman, B.; Roos, D.S. Purine salvage pathways in the apicomplexan parasite Toxoplasma gondii. J. Biol. Chem. 2004, 279, 31221–31227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roos, D.S.; Donald, R.G.; Morrissette, N.S.; Moulton, A.L. Molecular tools for genetic dissection of the protozoan parasite Toxoplasma gondii. Methods Cell Biol. 1994, 45, 27–63. [Google Scholar]
- Shen, B.; Brown, K.; Long, S.; Sibley, L.D. Development of CRISPR/Cas9 for Efficient Genome Editing in Toxoplasma gondii. Methods Mol. Biol. 2017, 1498, 79–103. [Google Scholar] [CrossRef]
- Shen, B.; Brown, K.M.; Lee, T.D.; Sibley, L.D. Efficient gene disruption in diverse strains of Toxoplasma gondii using CRISPR/CAS9. MBio 2014, 5, e01114. [Google Scholar] [CrossRef] [Green Version]
- Pishkari, S.; Shojaee, S.; Keshavarz, H.; Salimi, M.; Mohebali, M. Evaluation of Toxoplasma gondii soluble, whole and excretory/secretary antigens for diagnosis of toxoplasmosis by ELISA test. J. Parasit. Dis. 2017, 41, 289–291. [Google Scholar] [CrossRef] [Green Version]
- Melzer, T.C.; Cranston, H.J.; Weiss, L.M.; Halonen, S.K. Host Cell Preference of Toxoplasma gondii Cysts in Murine Brain: A Confocal Study. J. Neuroparasitol. 2010, 1. [Google Scholar] [CrossRef]
- Xia, N.; Yang, J.; Ye, S.; Zhang, L.; Zhou, Y.; Zhao, J.; David Sibley, L.; Shen, B. Functional analysis of Toxoplasma lactate dehydrogenases suggests critical roles of lactate fermentation for parasite growth in vivo. Cell. Microbiol. 2018, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burg, J.L.; Perelman, D.; Kasper, L.H.; Ware, P.L.; Boothroyd, J.C. Molecular analysis of the gene encoding the major surface antigen of Toxoplasma gondii. J. Immunol. 1988, 141, 3584–3591. [Google Scholar] [PubMed]
- Kasper, L.H. Isolation and characterization of a monoclonal anti-P30 antibody resistant mutant of Toxoplasma gondii. Parasite Immunol. 1987, 9, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Saraav, I.; Wang, Q.; Brown, K.M.; Sibley, L.D. Secretory Microneme Proteins Induce T-Cell Recall Responses in Mice Chronically Infected with Toxoplasma gondii. mSphere 2019, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturge, C.R.; Yarovinsky, F. Complex immune cell interplay in the gamma interferon response during Toxoplasma gondii infection. Infect. Immun. 2014, 82, 3090–3097. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Orellana, M.A.; Schreiber, R.D.; Remington, J.S. Interferon-gamma: The major mediator of resistance against Toxoplasma gondii. Science 1988, 240, 516–518. [Google Scholar] [CrossRef]
- Schwartzman, J.D.; Pfefferkorn, E.R. Toxoplasma gondii: Purine synthesis and salvage in mutant host cells and parasites. Exp. Parasitol. 1982, 53, 77–86. [Google Scholar] [CrossRef]
- Yarovinsky, F. Innate immunity to Toxoplasma gondii infection. Nat. Rev. Immunol. 2014, 14, 109–121. [Google Scholar] [CrossRef]
- Hunter, C.A.; Sibley, L.D. Modulation of innate immunity by Toxoplasma gondii virulence effectors. Nat. Rev. Microbiol. 2012, 10, 766–778. [Google Scholar] [CrossRef] [Green Version]
- Gazzinelli, R.T.; Wysocka, M.; Hieny, S.; Scharton-Kersten, T.; Cheever, A.; Kuhn, R.; Muller, W.; Trinchieri, G.; Sher, A. In the absence of endogenous IL-10, mice acutely infected with Toxoplasma gondii succumb to a lethal immune response dependent on CD4 + T cells and accompanied by overproduction of IL-12, IFN-gamma and TNF-alpha. J. Immunol. 1996, 157, 798–805. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Use |
|---|---|---|
| gRNA-ADSL-Fw | 5′- GACACTGCTCAATTTCGTCGGTTTTAGAGCTAGAAATAGC-3′ | To construct the ADSL specific CRISPR plasmid |
| gRNA-R | 5′-AACTTGACAT CCCCA TTTAC-3′ | To construct the ADSL specific CRISPR plasmid |
| pUC19-Fw pUC19-Rv | 5′-GGCGTAATCATGGTCATAGC-3′ 5′-CTCGAATTCACTGGCCGTCG-3′ | Amplification of pUC19 for Gibson assembly Amplification of pUC19 for Gibson assembly |
| loxp-DHFR*-loxp-Fw loxp-DHFR*-loxp-Rv 5′-GGACACGCTGAACTTGTGGC-3′ | 5′-CAACCCGCGCAGAAGACATC-3′ | Amplification of loxp-DHFR*-loxp for Gibson assembly Amplification of loxp-DHFR*-loxp for Gibson assembly |
| U5ADSL-Fw pADSL: loxp- DHFR*-loxp construction | 5′-GTTGTAAAACGACGGCCAGTCTAATTTTTGCCGGGTCTGG-3′ | Amplification of 5′-homology of ADSL for |
| U5ADSL-Rv pADSL: loxp- DHFR*-loxp construction | 5′-GATGTCTTCTGCGCGGGTTG CGAGACGAAGAAGAGAGC-3′ | Amplification of 5′-homology of ADSL for |
| U3ADSL-Fw pADSL: loxp -DHFR*-loxp construction | 5′-GCCACAAGTTCAGCGTGTCCACCTCAGTTGTCGGCACCGT-3′ | Amplification of 3′-homology of ADSL for |
| U3ADSL-Rv pADSL: loxp--DHFR*-loxp construction | 5′-GCTATGACCATGATTACGCCCGCAGACTCAAATCTATTC-3′ | Amplification of 3′-homology of ADSL for |
| 5′-UpU5 ADSL | 5′-GCTCTTGCTTTCGTCGCTGTC-3′ | PCR1 of ΔADSL:DHFR |
| 3′-InDHFR*-Fw | 5′-CGTGACCACGCCAAAGTAG-3′ | PCR1 of ΔADSL:DHFR |
| 5′-InDHFR*-Rv | 5′-GCACTTGCAGGATGAATTCC-3′ | PCR2 of ΔADSL:DHFR |
| 3′-DnU3ADSL | 5′-GTGTCTCGCACATGCGCGTT-3′ | PCR2 of ΔADSL:DHFR |
| 5′-UpgRNA ADSL | 5′- AACGCAATCAAGACGCTGGC-3′ | PCR3 of ΔADSL:DHFR |
| 3′-DngRNA ADSL | 5′- CGTCACTGTACCAGCGAGCAG-3′ | PCR3 of ΔADSL:DHFR |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Tang, D.; Yang, C.; Yang, J.; Fang, R. Toxoplasma gondii ADSL Knockout Provides Excellent Immune Protection against a Variety of Strains. Vaccines 2020, 8, 16. https://doi.org/10.3390/vaccines8010016
Wang L, Tang D, Yang C, Yang J, Fang R. Toxoplasma gondii ADSL Knockout Provides Excellent Immune Protection against a Variety of Strains. Vaccines. 2020; 8(1):16. https://doi.org/10.3390/vaccines8010016
Chicago/Turabian StyleWang, Luyao, Ding Tang, Chenghang Yang, Jing Yang, and Rui Fang. 2020. "Toxoplasma gondii ADSL Knockout Provides Excellent Immune Protection against a Variety of Strains" Vaccines 8, no. 1: 16. https://doi.org/10.3390/vaccines8010016