Protective Immunity against Vibrio harveyi in Grouper Induced by Single Vaccination with Poly (Lactide-co-glycolide) Microparticles Releasing Pleurocidin Peptide and Recombinant Glyceraldehyde-3-phosphate Dehydrogenase


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture
2.2. Preparation of PLG-PLE/rGAPDH MPs
2.3. Inactivated V. harveyi Bacteria
2.4. Bacterial Lysate
2.5. Animals
2.6. Immunization
2.7. Serum Assay
2.8. Lymphocyte Proliferation Assay
2.9. Quantitative Real-Time Reverse Transcriptase-PCR (qRT-PCR)
2.10. Bacterial Challenge
2.11. Statistical Analysis
3. Results
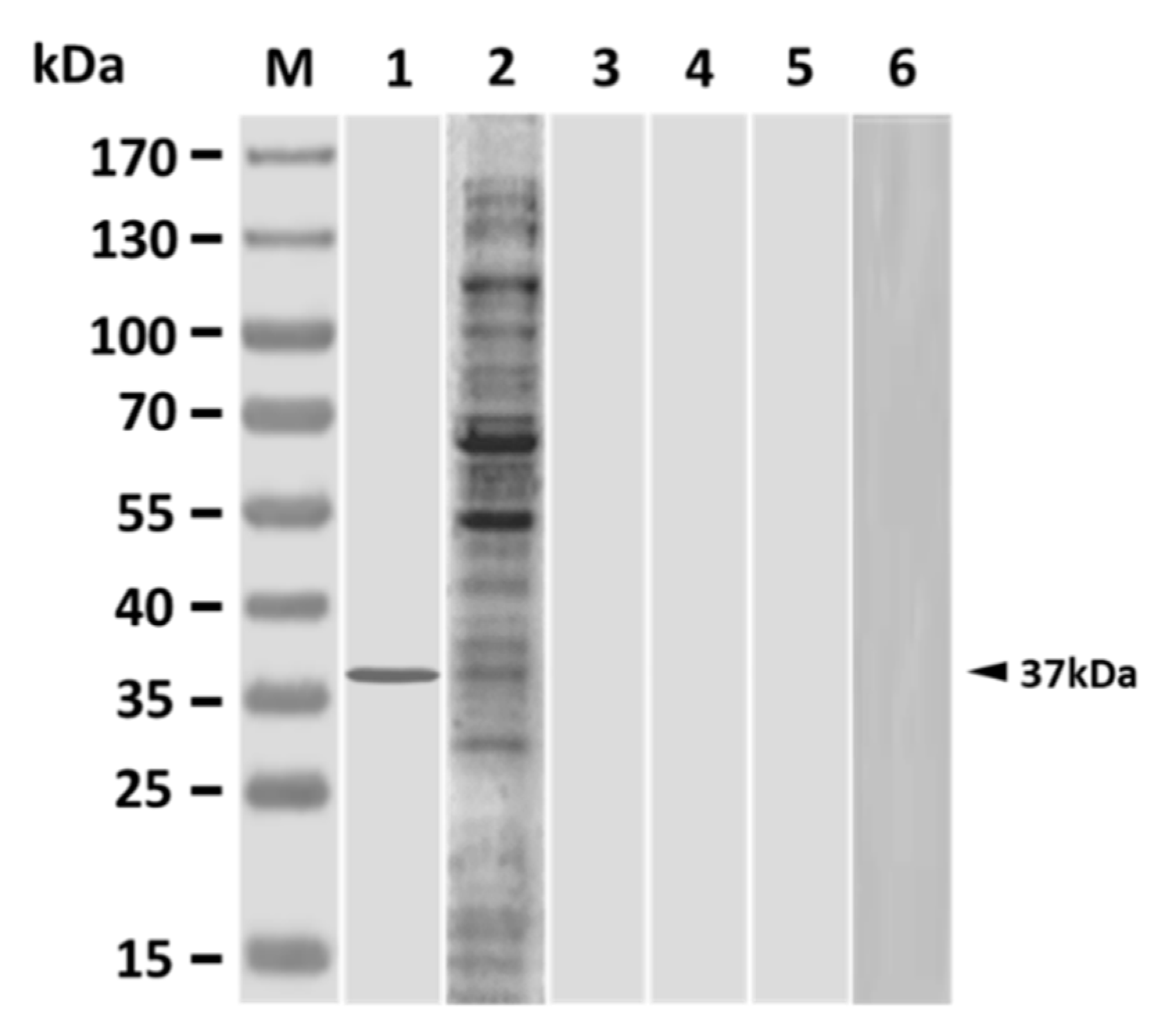
3.1. Antigenic Specificity of Grouper Antisera Following Single Immunization
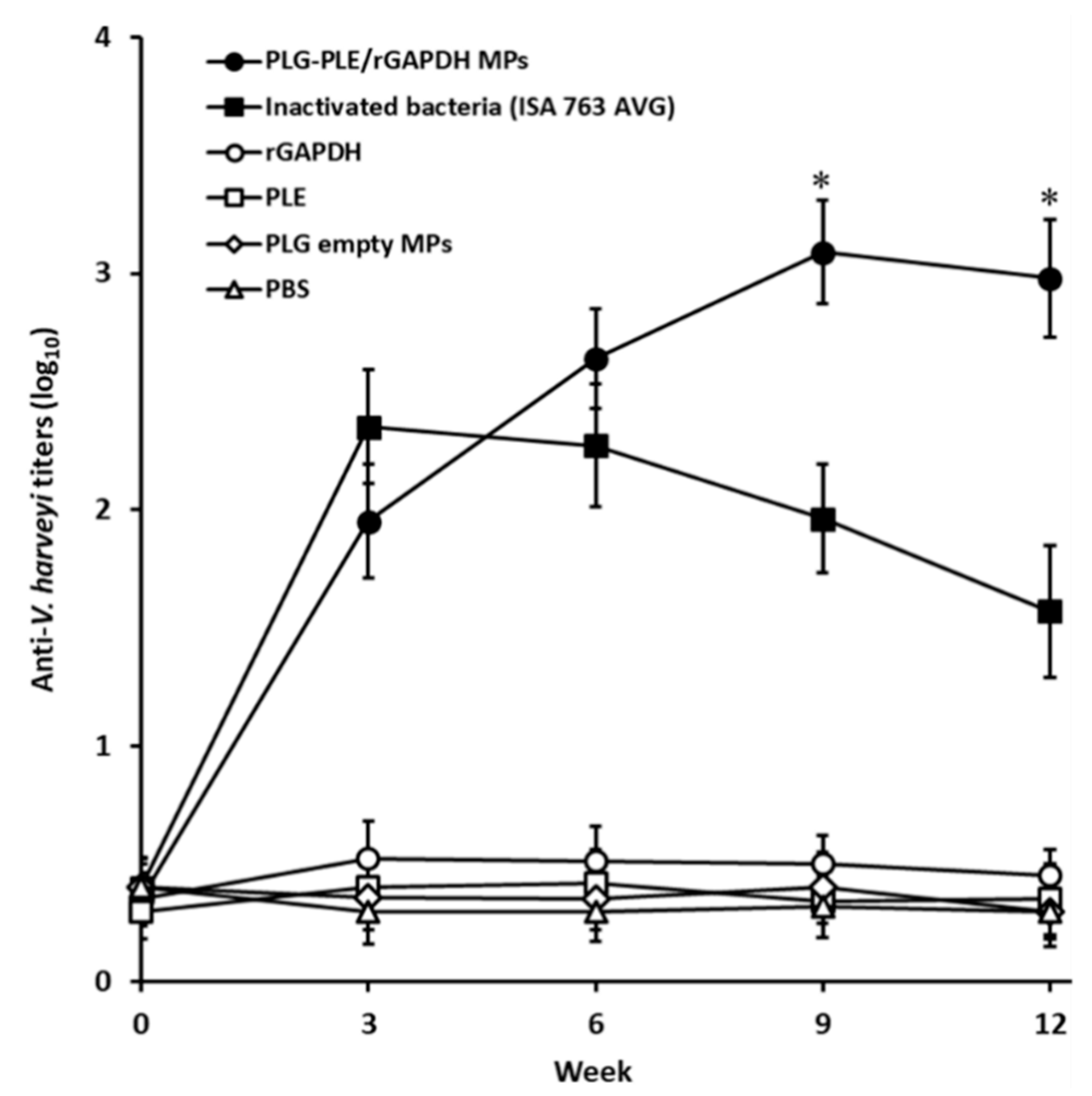
3.2. Sustained High Serum Titers Raised by Single Vaccination with PLG-PLE/rGAPDH MPs
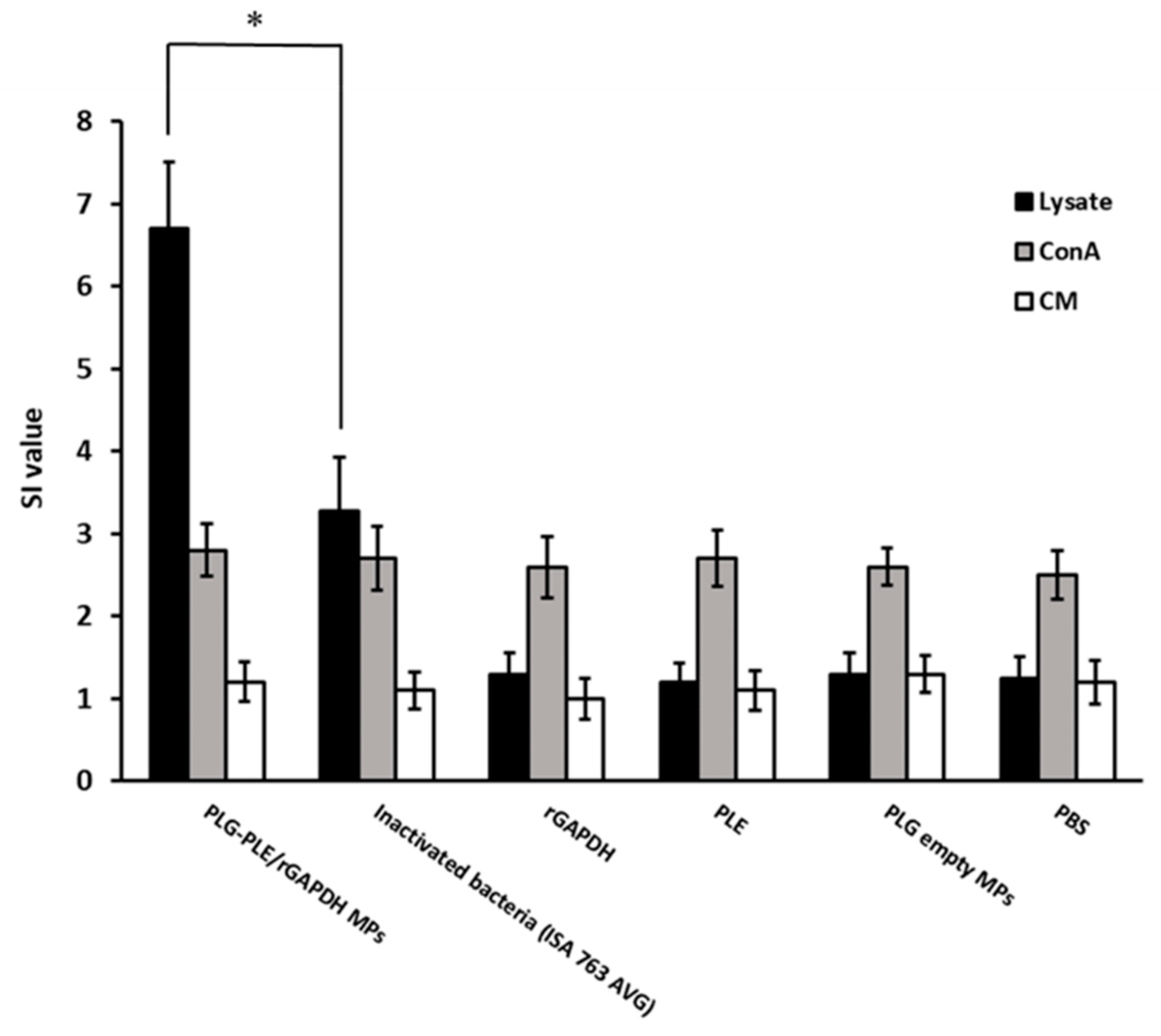
3.3. Robust Lymphocyte Proliferation Produced by Single Vaccination with PLG-PLE/rGAPDH MPs
3.4. High TNF-α Production Following Single Vaccination with PLG-PLE/rGAPDH MPs
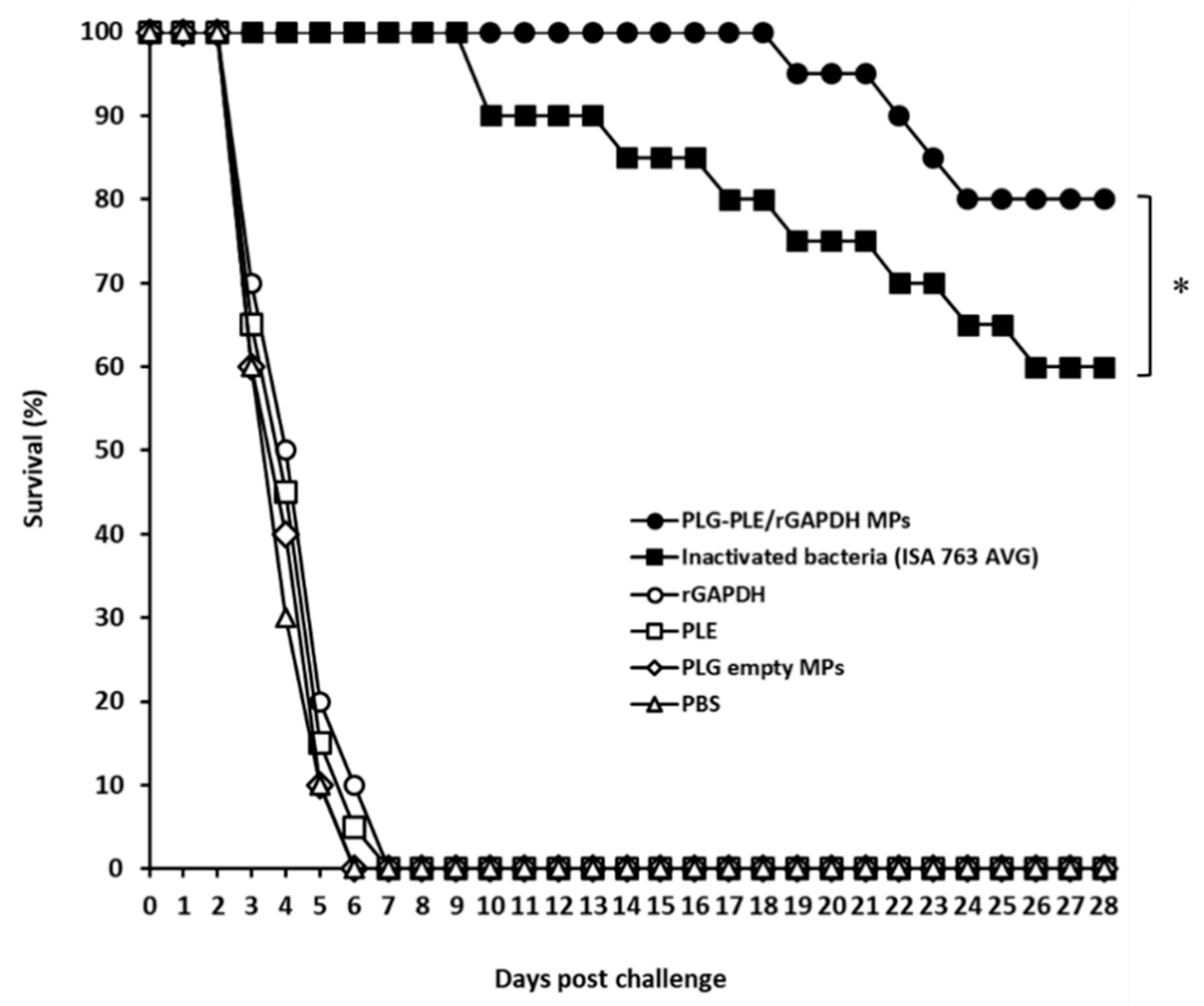
3.5. Strong Protection against V. harveyi in Grouper Induced by Single Vaccination with PLG-PLE/rGAPDH MPs
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Austin, B.; Zhang, X.H. Vibrio harveyi: A significant pathogen of marine vertebrates and invertebrates. Lett. Appl. Microbiol. 2006, 43, 119–124. [Google Scholar] [CrossRef]
- Alvarez, J.D. Vibrio harveyi: A pathogen of penaeid shrimps and fish in Venezuela. J. Fish Dis. 1998, 21, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.M.; Shi, C.Y.; Fan, C.; Jia, D.; Wang, S.Q.; Xie, G.S.; Li, G.Y.; Mo, Z.L.; Huang, J. Isolation, identification and pathogenicity of Vibrio harveyi, the causal agent of skin ulcer disease in juvenile hybrid groupers Epinephelus fuscoguttatus x Epinephelus lanceolatus. J. Fish Dis. 2017, 40, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Molecular studies, disease status and prophylactic measures in grouper aquaculture: Economic importance, diseases and immunology. Aquaculture 2010, 309, 1–14. [Google Scholar] [CrossRef]
- Liu, P.C.; Lee, K.K.; Chen, S.N. Susceptibility of different isolates of Vibrio harveyi to antibiotics. Microbios 1997, 91, 175–180. [Google Scholar]
- Hoelzer, K.; Bielke, L.; Blake, D.P.; Cox, E.; Cutting, S.M.; Devriendt, B.; Erlacher-Vindel, E.; Goossens, E.; Karaca, K.; Lemiere, S.; et al. Vaccines as alternatives to antibiotics for food producing animals. Part 2: New approaches and potential solutions. Vet. Res. 2018, 49, 70. [Google Scholar] [CrossRef] [Green Version]
- Hastein, T.; Gudding, R.; Evensen, O. Bacterial vaccines for fish--an update of the current situation worldwide. Dev. Biol. 2005, 121, 55–74. [Google Scholar]
- Chuang, S.C.; Huang, W.L.; Kau, S.W.; Yang, Y.P.; Yang, C.D. Pleurocidin Peptide Enhances Grouper Anti-Vibrio harveyi Immunity Elicited by Poly(lactide-co-glycolide)-Encapsulated Recombinant Glyceraldehyde-3-phosphate Dehydrogenase. Vaccines 2014, 2, 380–396. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yu, L.; Qian, R. Characterization of OmpK, GAPDH and their fusion OmpK-GAPDH derived from Vibrio harveyi outer membrane proteins: Their immunoprotective ability against vibriosis in large yellow croaker (Pseudosciaena crocea). J. Appl. Microbiol. 2007, 103, 1587–1599. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, L.; Qian, R. Cloning and expression of Vibrio harveyi OmpK* and GAPDH* genes and their potential application as vaccines in large yellow croakers Pseudosciaena crocea. J. Aquat. Anim. Health 2008, 20, 1–11. [Google Scholar] [CrossRef]
- Ningqiu, L.; Junjie, B.; Shuqin, W.; Xiaozhe, F.; Haihua, L.; Xing, Y.; Cunbin, S. An outer membrane protein, OmpK, is an effective vaccine candidate for Vibrio harveyi in orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2008, 25, 829–833. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yang, Z.; Bai, J.; Fu, X.; Liu, L.; Shi, C.; Wu, S. A shared antigen among Vibrio species: Outer membrane protein-OmpK as a versatile vibriosis vaccine candidate in orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2010, 28, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.P.; Hu, Y.H.; Sun, B.G.; Sun, L. Immunological study of the outer membrane proteins of Vibrio harveyi: Insights that link immunoprotectivity to interference with bacterial infection. Fish Shellfish Immunol. 2013, 35, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Sirover, M.A. On the functional diversity of glyceraldehyde-3-phosphate dehydrogenase: Biochemical mechanisms and regulatory control. Biochim. Biophys. Acta 2011, 1810, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C. Will advances in fish immunology change vaccination strategies? Fish Shellfish Immunol. 2008, 25, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Heegaard, P.M.; Dedieu, L.; Johnson, N.; Le Potier, M.F.; Mockey, M.; Mutinelli, F.; Vahlenkamp, T.; Vascellari, M.; Sorensen, N.S. Adjuvants and delivery systems in veterinary vaccinology: Current state and future developments. Arch. Virol. 2011, 156, 183–202. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, S.M.; Safhi, M.M.; Kannadasan, M.; Sukumaran, N. Vaccine adjuvants—Current status and prospects on controlled release adjuvancity. Saudi Pharm. J. 2011, 19, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J. Biol. Chem. 1997, 272, 12008–12013. [Google Scholar] [CrossRef] [Green Version]
- Cole, A.M.; Darouiche, R.O.; Legarda, D.; Connell, N.; Diamond, G. Characterization of a fish antimicrobial peptide: Gene expression, subcellular localization, and spectrum of activity. Antimicrob. Agents Chemother. 2000, 44, 2039–2045. [Google Scholar] [CrossRef] [Green Version]
- Peter Chiou, P.; Khoo, J.; Bols, N.C.; Douglas, S.; Chen, T.T. Effects of linear cationic alpha-helical antimicrobial peptides on immune-relevant genes in trout macrophages. Dev. Comp. Immunol. 2006, 30, 797–806. [Google Scholar] [CrossRef]
- Pundir, P.; Catalli, A.; Leggiadro, C.; Douglas, S.E.; Kulka, M. Pleurocidin, a novel antimicrobial peptide, induces human mast cell activation through the FPRL1 receptor. Mucosal Immunol. 2014, 7, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, M.; Jenssen, H.; Kindrachuk, J.; Scott, W.R.; Elliott, M.; Hilpert, K.; Cheng, J.T.; Hancock, R.E.; Straus, S.K. Structural studies of a peptide with immune modulating and direct antimicrobial activity. Chem. Biol. 2010, 17, 970–980. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Singh, M.; O’Hagan, D.T. Poly(lactide-co-glycolide) microparticles for the development of single-dose controlled-release vaccines. Adv. Drug Deliv. Rev. 1998, 32, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; O’Hagan, D.T.; Singh, M. The long-term potential of biodegradable poly(lactide-co-glycolide) microparticles as the next-generation vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 1731–1742. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.C.; Chung, Y.C.; Yang, C.D. Protective immunity against toxoplasmosis in mice induced by single-dose immunization with rSAG1/2 protein released from poly(lactide-co-glycolide) microparticles. Parasite 2017, 24, 5. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.L.; Chuang, S.C.; Yang, C.D. Anti-Idiotype Vaccine Provides Protective Immunity Against Vibrio harveyi in Grouper (Epinephelus coioides). Vaccines 2019, 7, 210. [Google Scholar] [CrossRef] [Green Version]
- Chuang, S.C.; Yang, C.D. Sustained release of recombinant surface antigen 2 (rSAG2) from poly(lactide-co-glycolide) microparticles extends protective cell-mediated immunity against Toxoplasma gondii in mice. Parasitology 2014, 141, 1657–1666. [Google Scholar] [CrossRef]
- Dan, X.M.; Zhang, T.W.; Li, Y.W.; Li, A.X. Immune responses and immune-related gene expression profile in orange-spotted grouper after immunization with Cryptocaryon irritans vaccine. Fish Shellfish Immunol. 2013, 34, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Luo, X.C.; Dan, X.M.; Huang, X.Z.; Qiao, W.; Zhong, Z.P.; Li, A.X. Orange-spotted grouper (Epinephelus coioides) TLR2, MyD88 and IL-1beta involved in anti-Cryptocaryon irritans response. Fish Shellfish Immunol. 2011, 30, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.C.; Ko, J.C.; Chen, C.P.; Du, J.T.; Yang, C.D. Induction of long-lasting protective immunity against Toxoplasma gondii in BALB/c mice by recombinant surface antigen 1 protein encapsulated in poly (lactide-co-glycolide) microparticles. Parasit. Vectors 2013, 6, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, S.C.; Ko, J.C.; Chen, C.P.; Du, J.T.; Yang, C.D. Encapsulation of chimeric protein rSAG1/2 into poly(lactide-co-glycolide) microparticles induces long-term protective immunity against Toxoplasma gondii in mice. Exp. Parasitol. 2013, 134, 430–437. [Google Scholar] [CrossRef]
- Raman, C.; Berkland, C.; Kim, K.; Pack, D.W. Modeling small-molecule release from PLG microspheres: Effects of polymer degradation and nonuniform drug distribution. J. Control. Release 2005, 103, 149–158. [Google Scholar] [CrossRef]
- Sturesson, C.; Carlfors, J. Incorporation of protein in PLG-microspheres with retention of bioactivity. J. Control. Release 2000, 67, 171–178. [Google Scholar] [CrossRef]
- Kavanagh, O.V.; Earley, B.; Murray, M.; Foster, C.J.; Adair, B.M. Antigen-specific IgA and IgG responses in calves inoculated intranasally with ovalbumin encapsulated in poly(DL-lactide-co-glycolide) microspheres. Vaccine 2003, 21, 4472–4480. [Google Scholar] [CrossRef]
- Uchida, M.; Natsume, H.; Kishino, T.; Seki, T.; Ogihara, M.; Juni, K.; Kimura, M.; Morimoto, Y. Immunization by particle bombardment of antigen-loaded poly-(DL-lactide-co-glycolide) microspheres in mice. Vaccine 2006, 24, 2120–2130. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. The Function of Fish Cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Lam, F.W.; Wu, S.Y.; Lin, S.J.; Lin, C.C.; Chen, Y.M.; Wang, H.C.; Chen, T.Y.; Lin, H.T.; Lin, J.H. The expression of two novel orange-spotted grouper (Epinephelus coioides) TNF genes in peripheral blood leukocytes, various organs, and fish larvae. Fish Shellfish Immunol. 2011, 30, 618–629. [Google Scholar] [CrossRef]
- Hu, Y.H.; Deng, T.; Sun, B.G.; Sun, L. Development and efficacy of an attenuated Vibrio harveyi vaccine candidate with cross protectivity against Vibrio alginolyticus. Fish Shellfish Immunol. 2012, 32, 1155–1161. [Google Scholar] [CrossRef]
- Ye, M.; Kim, S.; Park, K. Issues in long-term protein delivery using biodegradable microparticles. J. Control. Release 2010, 146, 241–260. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Nucleotide Sequence (5′ → 3′) | Amplification Size (bp) | Accession Number/Reference |
|---|---|---|---|
| TNF-α (F) | GTGTCCTGCTGTTTGCTTGGTA | 207 | FJ009049/[29] |
| TNF-α (R) | CAGTGTCCGACTTGATTAGTGCTT | ||
| β actin (F) | TGCTGTCCCTGTATGCCTCT | 225 | AY510710/[30] |
| β actin (R) | CCTTGATGTCACGCACGAT |
| Group a | TNF-α (Fold Change) b | |
|---|---|---|
| Lysate | Con A | |
| PLG-PLE/rGAPDH MPs | 12.4 ± 1.5 c | 2.7 ± 0.9 f |
| Inactivated bacteria (ISA 763 AVG) | 3.2 ± 0.5 d | 2.8 ± 0.6 f |
| rGAPDH | 1.4 ± 0.3 e | 2.7 ± 0.7 f |
| PLE | 1.2 ± 0.4 e | 2.6 ± 0.5 f |
| PLG empty MPs | 1.1 ± 0.2 e | 2.5 ± 0.7 f |
| PBS | 0.9 ± 0.3 e | 2.4 ± 0.5 f |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.-P.; Chuang, S.-C.; Yang, C.-D. Protective Immunity against Vibrio harveyi in Grouper Induced by Single Vaccination with Poly (Lactide-co-glycolide) Microparticles Releasing Pleurocidin Peptide and Recombinant Glyceraldehyde-3-phosphate Dehydrogenase. Vaccines 2020, 8, 33. https://doi.org/10.3390/vaccines8010033
Liu S-P, Chuang S-C, Yang C-D. Protective Immunity against Vibrio harveyi in Grouper Induced by Single Vaccination with Poly (Lactide-co-glycolide) Microparticles Releasing Pleurocidin Peptide and Recombinant Glyceraldehyde-3-phosphate Dehydrogenase. Vaccines. 2020; 8(1):33. https://doi.org/10.3390/vaccines8010033
Chicago/Turabian StyleLiu, Shang-Pin, Shu-Chun Chuang, and Chung-Da Yang. 2020. "Protective Immunity against Vibrio harveyi in Grouper Induced by Single Vaccination with Poly (Lactide-co-glycolide) Microparticles Releasing Pleurocidin Peptide and Recombinant Glyceraldehyde-3-phosphate Dehydrogenase" Vaccines 8, no. 1: 33. https://doi.org/10.3390/vaccines8010033
APA StyleLiu, S.-P., Chuang, S.-C., & Yang, C.-D. (2020). Protective Immunity against Vibrio harveyi in Grouper Induced by Single Vaccination with Poly (Lactide-co-glycolide) Microparticles Releasing Pleurocidin Peptide and Recombinant Glyceraldehyde-3-phosphate Dehydrogenase. Vaccines, 8(1), 33. https://doi.org/10.3390/vaccines8010033