Protective Immunity Elicited by VP1 Chimeric Antigens of Bacterial Ghosts against Hand-Foot-and-Mouth Disease Virus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Cells, Plasmids and Virus
2.2. Mice
2.3. Construction and Preparation of pOEVP1 and pOCVP1
2.4. Preparation of Bacterial Ghosts and Whole Cells [18]
2.5. Analysis of Antigens by Fluorescence Activating Cell Sorter (FACS )
2.6. Analysis of Cytotoxicity by MTT(3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide)
2.7. Mice Immunization and Challenge
2.8. Detection of Specific Antibodies
2.9. Statistical Analysis
3. Results
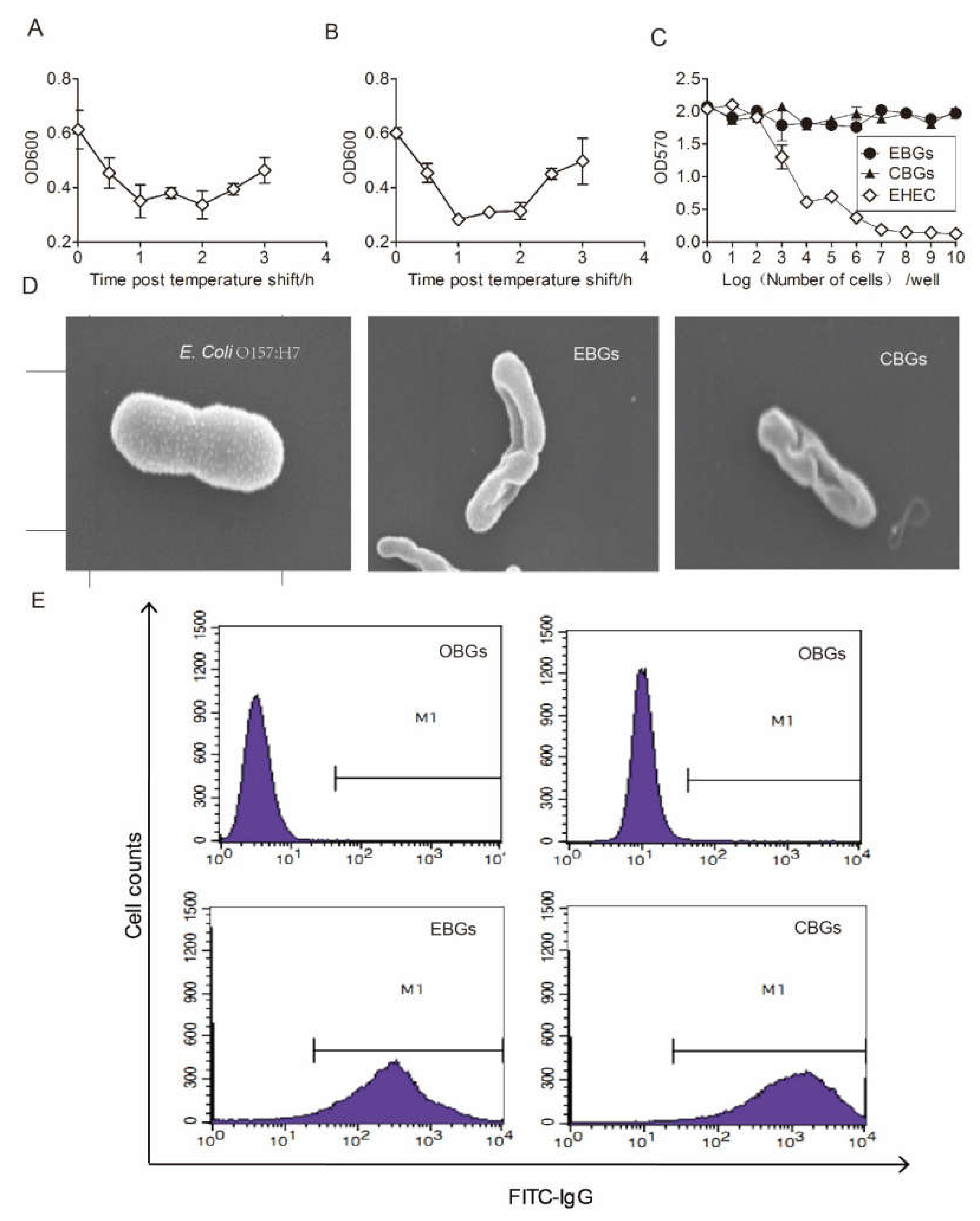
3.1. Preparation and Evaluation of the Chimeric BG Vaccine Candidates
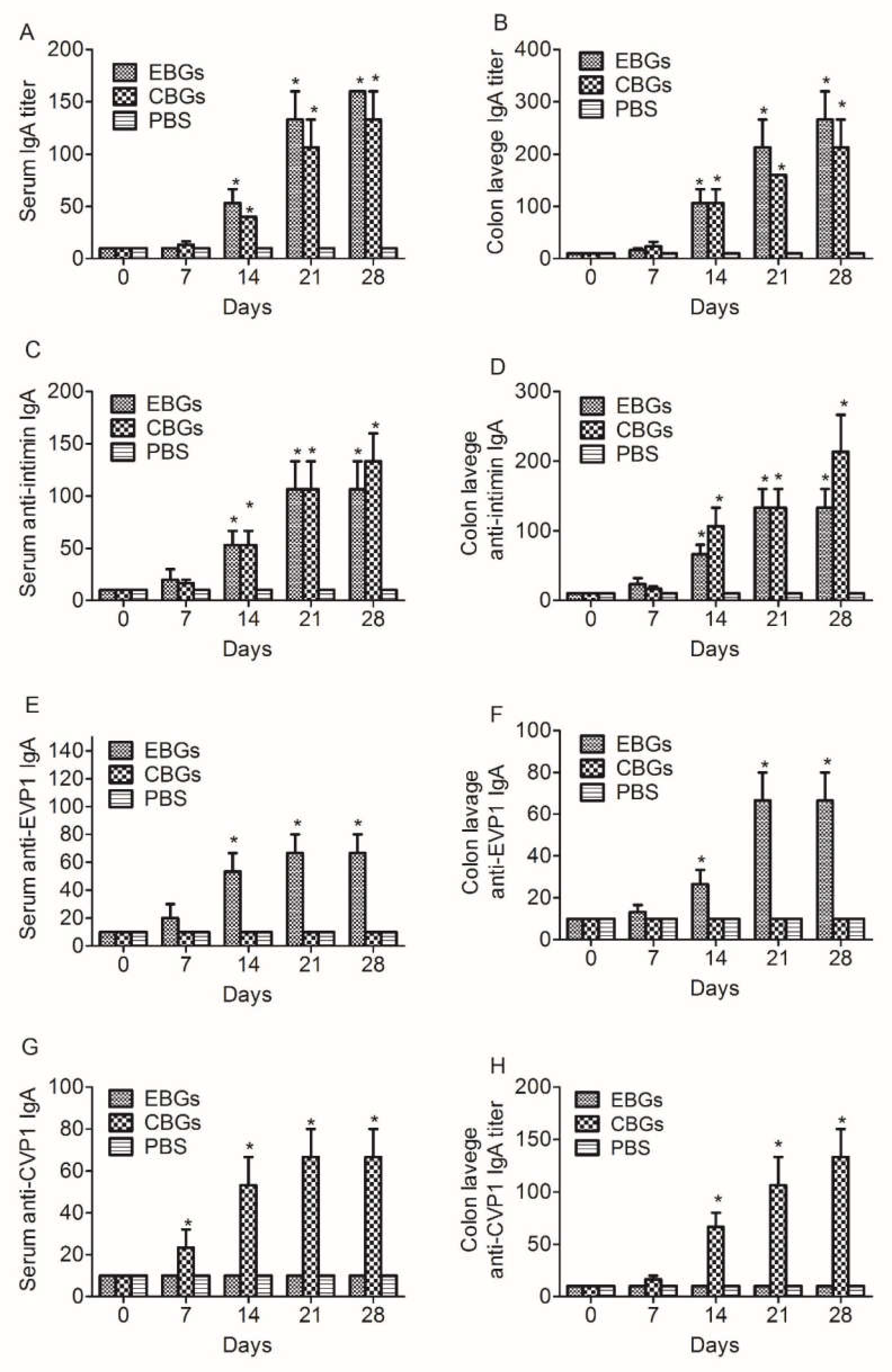
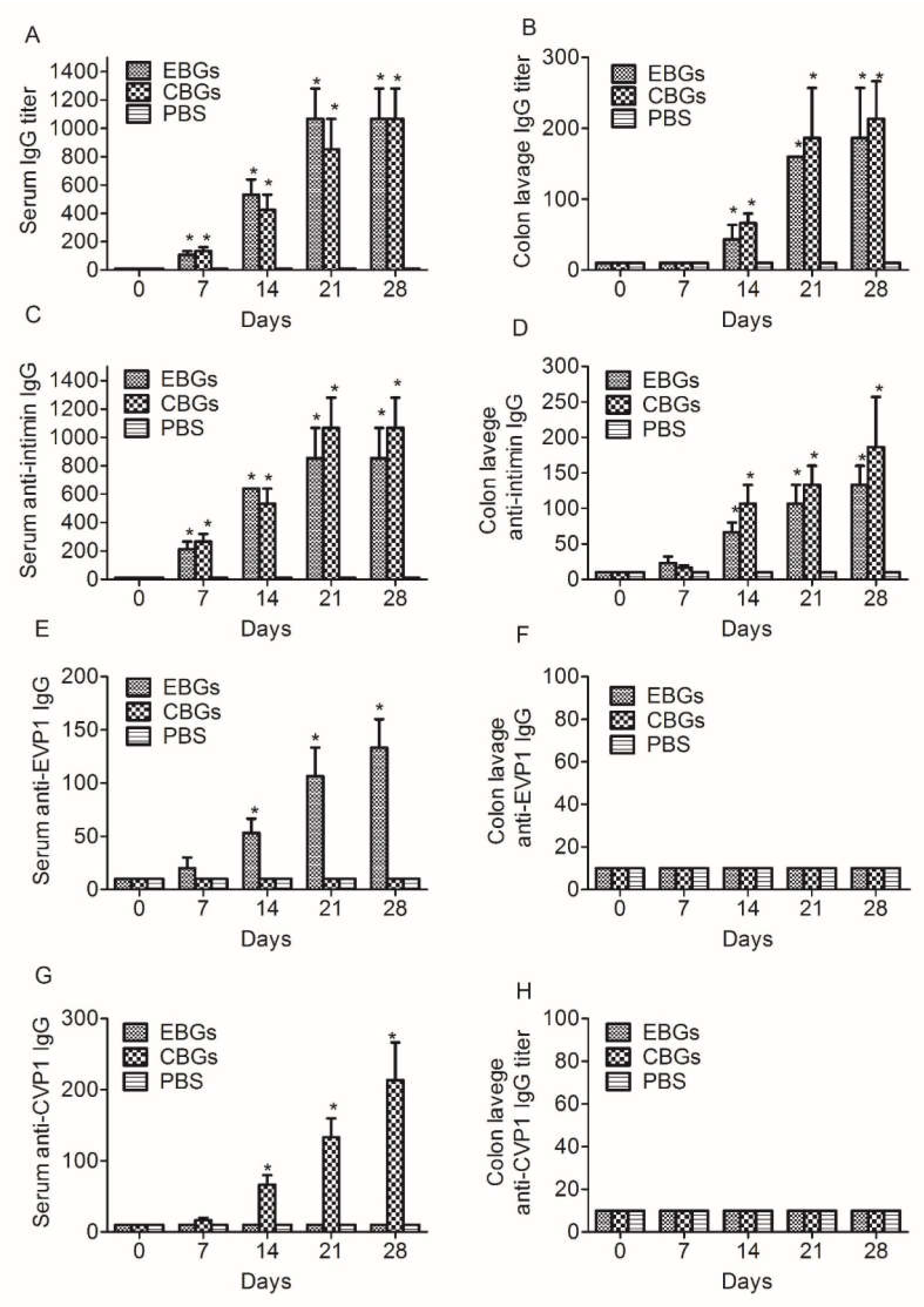
3.2. The Mucosal Immune Response Elicited by Immunization with EBGs or CBGs
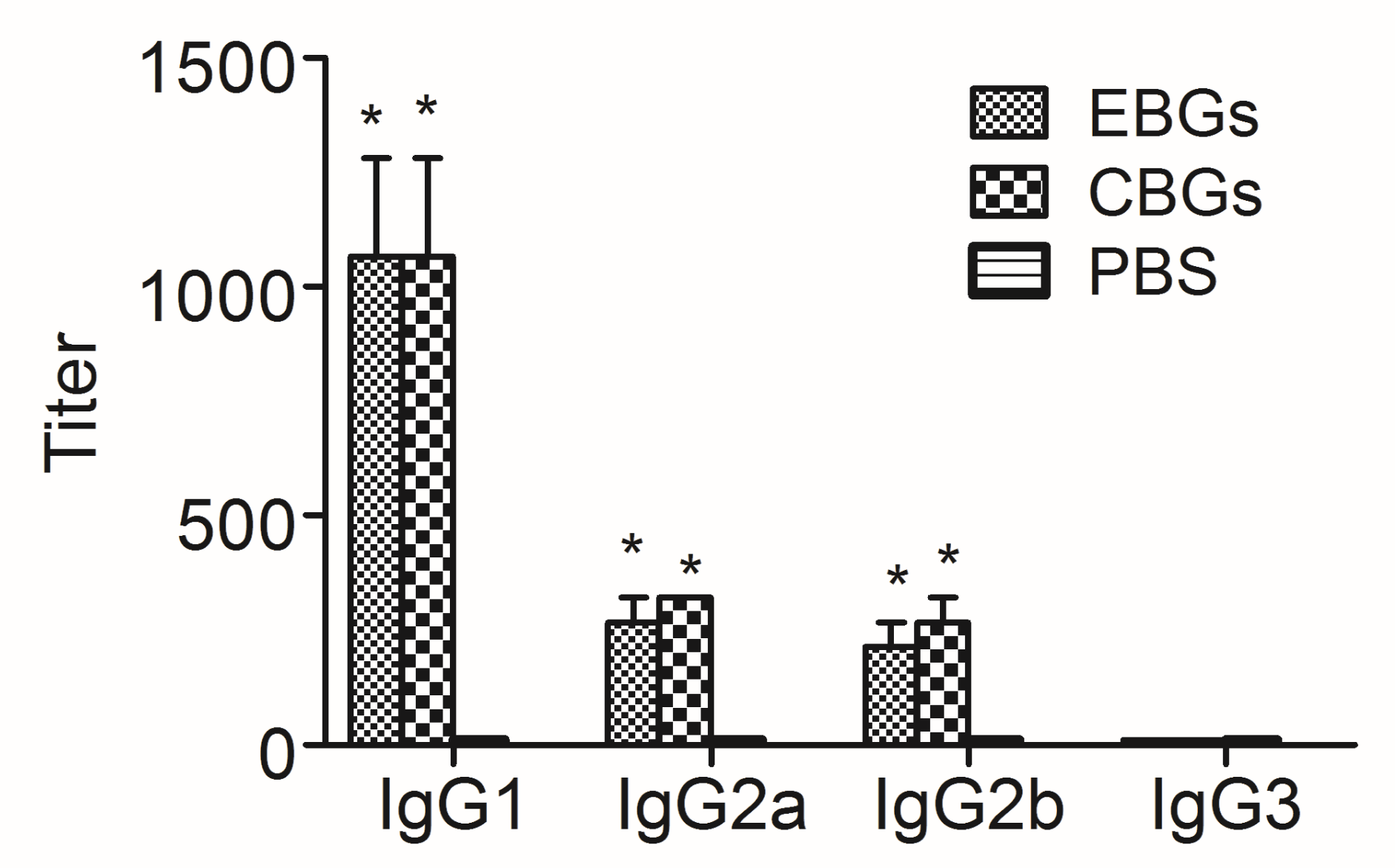
3.3. The Humoral Immune Response against HFMD Virus Elicited by Immunization with EBGs or CBGs
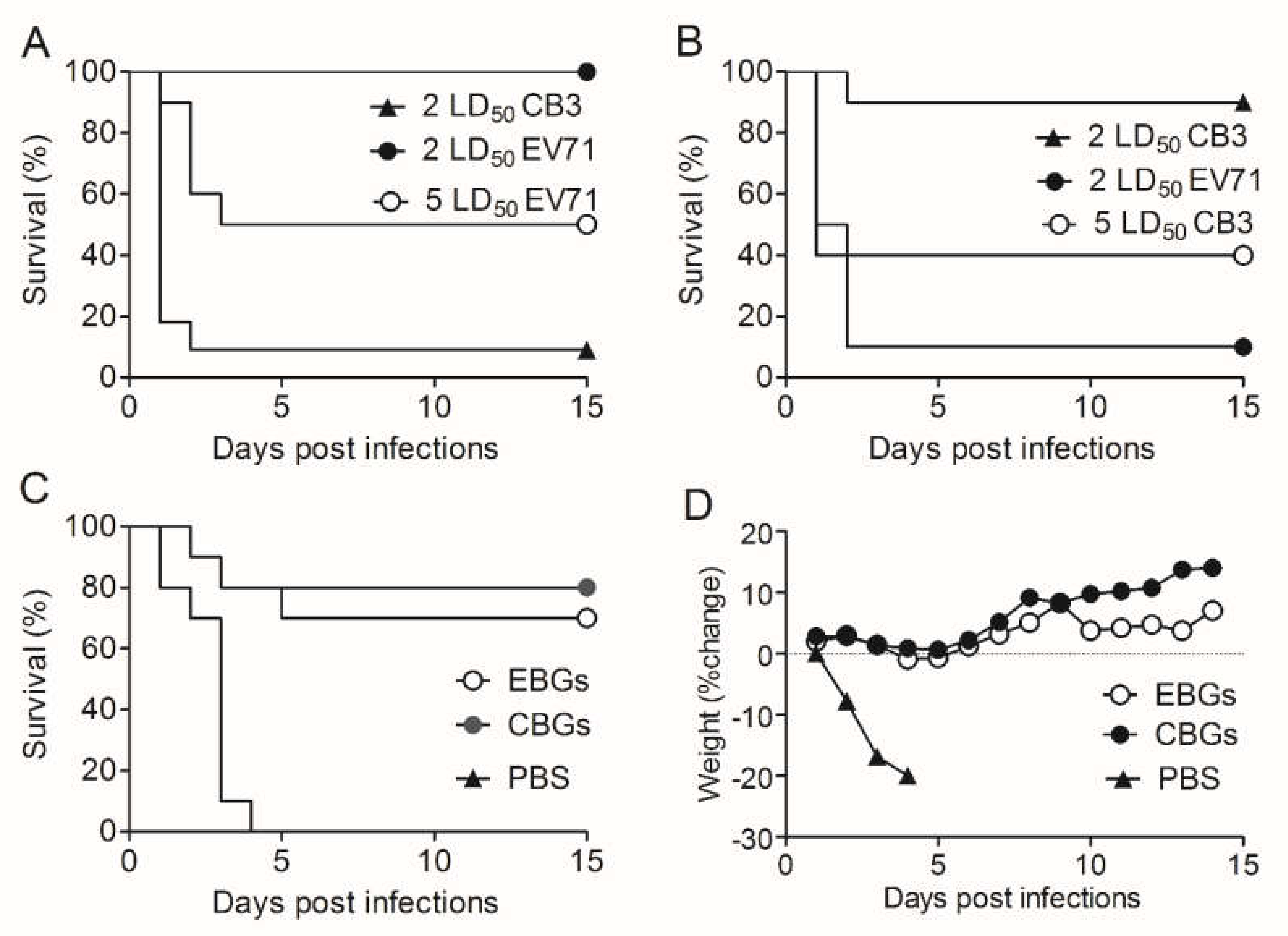
3.4. Efficacy of EBGs and CBGs Immunization in Generating Protective Immunity against the HFMD Virus
3.5. Efficacy of EBGs and CBGs Immunizations in Generating Cross-Protective Immunity against E. coli O157:H788321
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, B.; Li, J.; Wang, Y.; Du, N.; Sun, L.; Xiao, H.; Zhao, Y.; Bao, W.; Zhang, W. Understanding the epidemiological characteristics of EV71 and CVA16 infection to aid the diagnosis and treatment of hand, foot, and mouth disease. J. Med Virol. 2019, 91, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Pathinayake, P.S.; Hsu, A.C.; Wark, P.A. Innate immunity and immune evasion by enterovirus 71. Viruses 2015, 7, 6613–6630. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Cheng, P.Y.; Chin, C.L.; Huang, L.M.; Lin, S.Y.; Chiang, B.L. Fibroblast-stimulating lipopeptide-1 as a potential mucosal adjuvant enhances mucosal and systemic immune responses to enterovirus 71 vaccine. Vaccine 2018, 36, 4331–4338. [Google Scholar] [CrossRef]
- Wang, L.C.; Kao, C.M.; Ling, P.; Su, I.J.; Chang, T.M.; Chen, S.H. CD4 T-cell-independent antibody response reduces enterovirus 71 lethality in mice by decreasing tissue viral loads. Clin. Dev. Immunol. 2012, 2012, 580696. [Google Scholar] [CrossRef]
- Tung, W.S.; Bakar, S.A.; Sekawi, Z.; Rosli, R. DNA vaccine constructs against enterovirus 71 elicit immune response in mice. Genet. Vaccines Ther. 2007, 5, 6. [Google Scholar] [CrossRef]
- Crawford, N.W.; Graham, S.M. EV71 vaccine: protection from a previously neglected disease. Lancet 2013, 381, 1968–1970. [Google Scholar] [CrossRef]
- Park, K.B.; Lim, B.K.; Ye, M.B.; Chung, S.Y.; Nam, J.H. A peptide vaccine based on a B-cell epitope on the VP1 protein of enterovirus 70 induces a strong antibody response. Acta. Virologica. 2012, 56, 337–342. [Google Scholar] [CrossRef]
- Xu, L.; He, D.; Li, Z.; Zheng, J.; Yang, L.; Yu, M.; Yu, H.; Chen, Y.; Que, Y.; Shih, J.W.; et al. Protection against lethal enterovirus 71 challenge in mice by a recombinant vaccine candidate containing a broadly cross-neutralizing epitope within the VP2 EF loop. Theranostics 2014, 4, 498–513. [Google Scholar] [CrossRef]
- Witte, A.; Wanner, G.; Sulzner, M.; Lubitz, W. Dynamics of PhiX174 protein E-mediated lysis of Escherichia coli. Arch. Microbiol. 1992, 157, 381–388. [Google Scholar] [CrossRef]
- Witte, A.; Lubitz, W. Biochemical characterization of phi X174-protein-E-mediated lysis of Escherichia coli. Eur. J. Biochem. 1989, 180, 393–398. [Google Scholar] [CrossRef]
- Riedmann, E.M.; Kyd, J.M.; Cripps, A.W.; Lubitz, W. Bacterial ghosts as adjuvant particles. Expert Rev. Vaccines 2007, 6, 241–253. [Google Scholar] [CrossRef]
- Jiao, H.; Yang, H.; Zhao, D.; Chen, J.; Zhang, Q.; Liang, J.; Yin, Y.; Kong, G.; Li, G. Design and immune characterization of a novel Neisseria gonorrhoeae DNA vaccine using ghosts as vector and adjuvant. Vaccine 2018, 36, 4532–4539. [Google Scholar] [CrossRef]
- Eko, F.O.; Schukovskaya, T.; Lotzmanova, E.Y.; Firstova, V.V.; Emalyanova, N.V.; Klueva, S.N.; Kravtzov, A.L.; Livanova, L.F.; Kutyrev, V.V.; Igietseme, J.U.; et al. Evaluation of the protective efficacy of Vibrio cholerae ghost (VCG) candidate vaccines in rabbits. Vaccine 2003, 21, 3663–3674. [Google Scholar] [CrossRef]
- Panthel, K.; Jechlinger, W.; Matis, A.; Rohde, M.; Szostak, M.; Lubitz, W.; Haas, R. Generation of Helicobacter pylori ghosts by PhiX protein E-mediated inactivation and their evaluation as vaccine candidates. Infect. Immun. 2003, 71, 109–116. [Google Scholar] [CrossRef]
- Cai, K.; Tu, W.; Liu, Y.; Li, T.; Wang, H. Novel fusion antigen displayed-bacterial ghosts vaccine candidate against infection of Escherichia coli O157:H7. Sci. Rep. 2015, 5, 17479. [Google Scholar] [CrossRef]
- Chong, P.; Liu, C.C.; Chow, Y.H.; Chou, A.H.; Klein, M. Review of enterovirus 71 vaccines. Clin. Infect. Dis.: Off. Publ. Infect. Dis. Soc. Am. 2015, 60, 797–803. [Google Scholar] [CrossRef]
- Pistor, S.; Hobom, G. Expression of viral hemagglutinin on the surface of E. coli. Klin. Wochenschr. 1988, 66, 110–116. [Google Scholar] [CrossRef]
- Mader, H.J.; Szostak, M.P.; Hensel, A.; Lubitz, W.; Haslberger, A.G. Endotoxicity does not limit the use of bacterial ghosts as candidate vaccines. Vaccine 1997, 15, 195–202. [Google Scholar] [CrossRef]
- Cai, K.; Gao, X.; Li, T.; Hou, X.; Wang, Q.; Liu, H.; Xiao, L.; Tu, W.; Liu, Y.; Shi, J.; et al. Intragastric immunization of mice with enterohemorrhagic Escherichia coli O157:H7 bacterial ghosts reduces mortality and shedding and induces a Th2-type dominated mixed immune response. Can. J. Microbiol. 2010, 56, 389–398. [Google Scholar] [CrossRef]
- Fanghong, C.; Xiong, L.; Nan, N.; Zhan, L.; Nianzhi, N.; Deyan, L.; Hui, W. VP1 of Enterovirus 71 Protects Mice Against Enterovirus 71 and Coxsackievirus B3 in Lethal Challenge Experiment. Front. Immunol 2019, 10, 2564. [Google Scholar]
- Wu, C.Y.; Yu, S.L.; Chen, Y.T.; Chen, Y.H.; Hsiao, P.W.; Chow, Y.H.; Chen, J.R. The mature EV71 virion induced a broadly cross-neutralizing VP1 antibody against subtypes of the EV71 virus. PLoS One 2019, 14, e0210553. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, J.; Li, B.; Guo, L.; Li, H.; Song, J.; Yang, Z.; Li, H.; Fan, H.; Huang, X.; et al. Correction: A Novel Neutralizing Antibody Specific to the DE Loop of VP1 Can Inhibit EV-D68 Infection in Mice. J. Immunol. 2019, 202, 1905–1906. [Google Scholar] [CrossRef]
- Walcher, P.; Mayr, U.B.; Azimpour-Tabrizi, C.; Eko, F.O.; Jechlinger, W.; Mayrhofer, P.; Alefantis, T.; Mujer, C.V.; DelVecchio, V.G.; Lubitz, W. Antigen discovery and delivery of subunit vaccines by nonliving bacterial ghost vectors. Expert Rev. Vaccines 2004, 3, 681–691. [Google Scholar] [CrossRef]
- Jechlinger, W.; Haller, C.; Resch, S.; Hofmann, A.; Szostak, M.P.; Lubitz, W. Comparative immunogenicity of the hepatitis B virus core 149 antigen displayed on the inner and outer membrane of bacterial ghosts. Vaccine 2005, 23, 3609–3617. [Google Scholar] [CrossRef]
- Ebensen, T.; Paukner, S.; Link, C.; Kudela, P.; de Domenico, C.; Lubitz, W.; Guzman, C.A. Bacterial ghosts are an efficient delivery system for DNA vaccines. J. Immunol. 2004, 172, 6858–6865. [Google Scholar] [CrossRef]
- Klose, M.; Jahnig, F.; Hindennach, I.; Henning, U. Restoration of membrane incorporation of an Escherichia coli outer membrane protein (OmpA) defective in membrane insertion. J. Biol. Chem. 1989, 264, 21842–21847. [Google Scholar]
- Dornmair, K.; Kiefer, H.; Jahnig, F. Refolding of an integral membrane protein. OmpA of Escherichia coli. J. Biol. Chem. 1990, 265, 18907–18911. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, S.; Nan, N.; Sun, Y.; He, Z.; Li, J.; Chen, F.; Li, T.; Ning, N.; Wang, J.; Li, Z.; et al. Protective Immunity Elicited by VP1 Chimeric Antigens of Bacterial Ghosts against Hand-Foot-and-Mouth Disease Virus. Vaccines 2020, 8, 61. https://doi.org/10.3390/vaccines8010061
Gong S, Nan N, Sun Y, He Z, Li J, Chen F, Li T, Ning N, Wang J, Li Z, et al. Protective Immunity Elicited by VP1 Chimeric Antigens of Bacterial Ghosts against Hand-Foot-and-Mouth Disease Virus. Vaccines. 2020; 8(1):61. https://doi.org/10.3390/vaccines8010061
Chicago/Turabian StyleGong, Saisai, Nan Nan, Yakun Sun, Zhili He, Jiajia Li, Fanghong Chen, Tao Li, Nianzhi Ning, Jianxin Wang, Zhan Li, and et al. 2020. "Protective Immunity Elicited by VP1 Chimeric Antigens of Bacterial Ghosts against Hand-Foot-and-Mouth Disease Virus" Vaccines 8, no. 1: 61. https://doi.org/10.3390/vaccines8010061
APA StyleGong, S., Nan, N., Sun, Y., He, Z., Li, J., Chen, F., Li, T., Ning, N., Wang, J., Li, Z., Luo, D., & Wang, H. (2020). Protective Immunity Elicited by VP1 Chimeric Antigens of Bacterial Ghosts against Hand-Foot-and-Mouth Disease Virus. Vaccines, 8(1), 61. https://doi.org/10.3390/vaccines8010061