Synthetic Peptides Containing Three Neutralizing Epitopes of Genotype 4 Swine Hepatitis E Virus ORF2 induced Protection against Swine HEV Infection in Rabbit

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
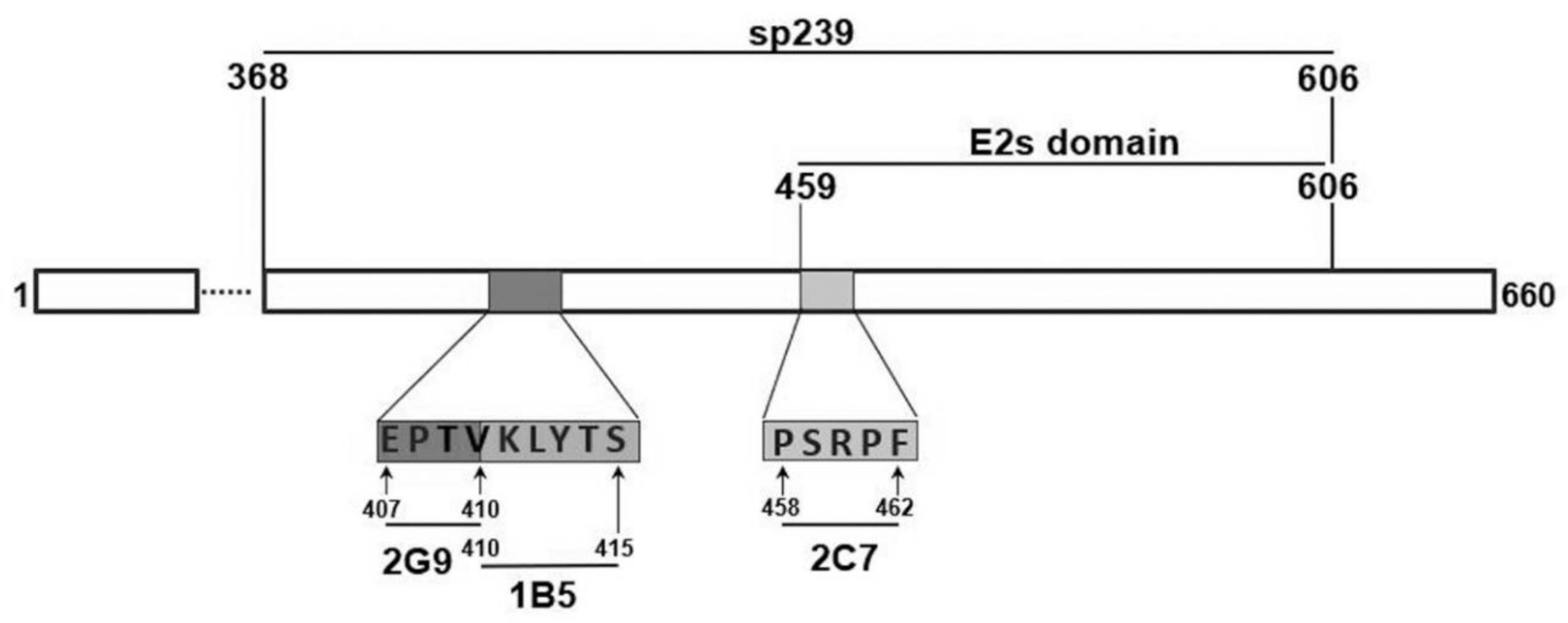
2.1. Synthesis Peptides, sp239, and ORF3 Protein and Virus
2.2. Immunization of Rabbits with Synthetic Peptides of Three Neutralizing Epitopes of Genotype 4 Swine HEV ORF2
2.3. Virus Challenge and Sample Collection
2.4. Indirect ELISA
2.5. Qualitative and Quantitative Detection of Swine HEV RNA Using RT-PCR
2.6. Determination of Serum Alanine Aminotransferase (ALT) Levels
2.7. Gross and Microscopic Hepatic Lesions
2.8. Statistical Analysis
2.9. Ethics Statement
3. Results
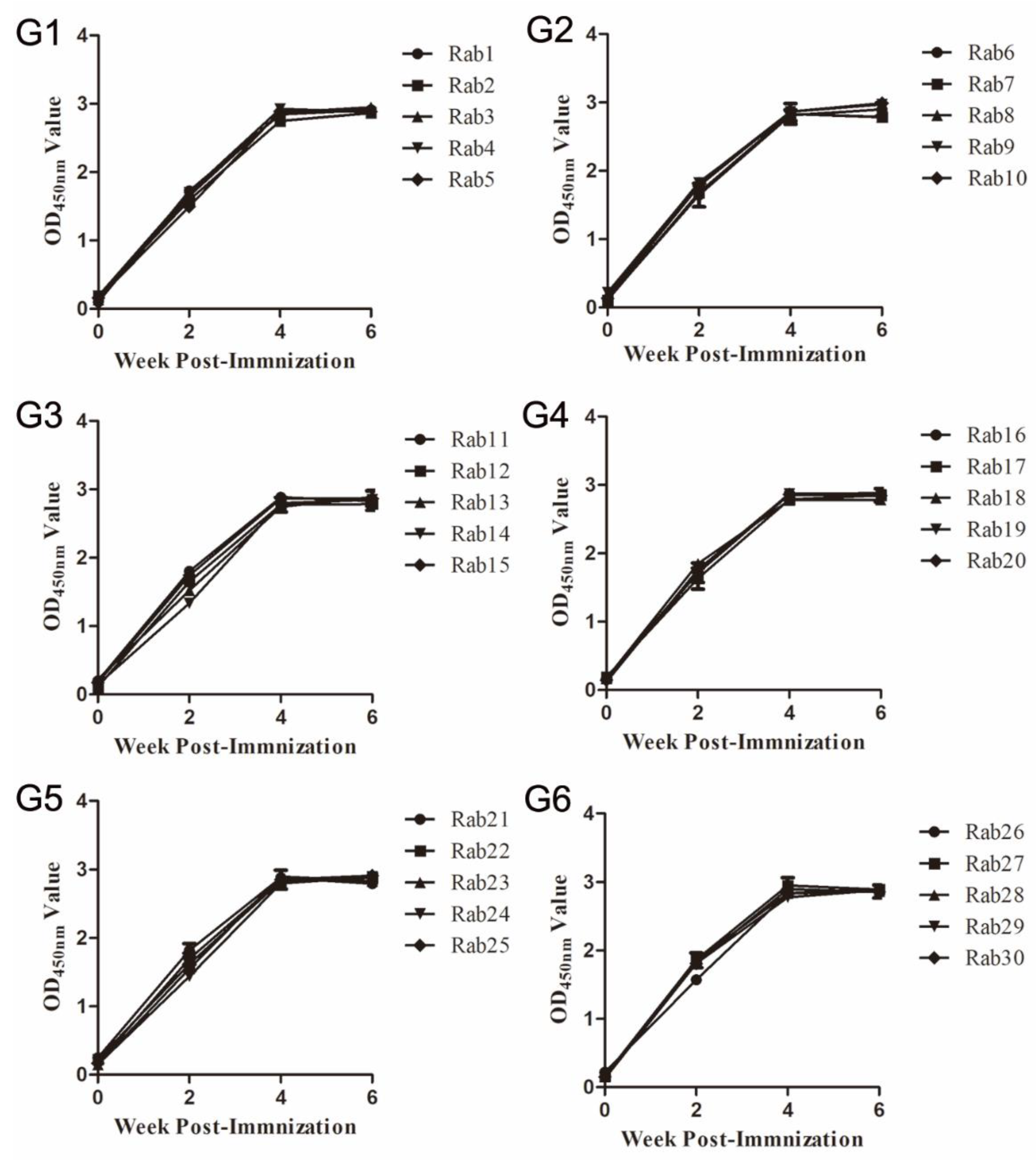
3.1. Immune Responses in Rabbits Immunized with Synthetic Peptides of Three Neutralizing Epitopes of Genotype 4 Swine HEV ORF2
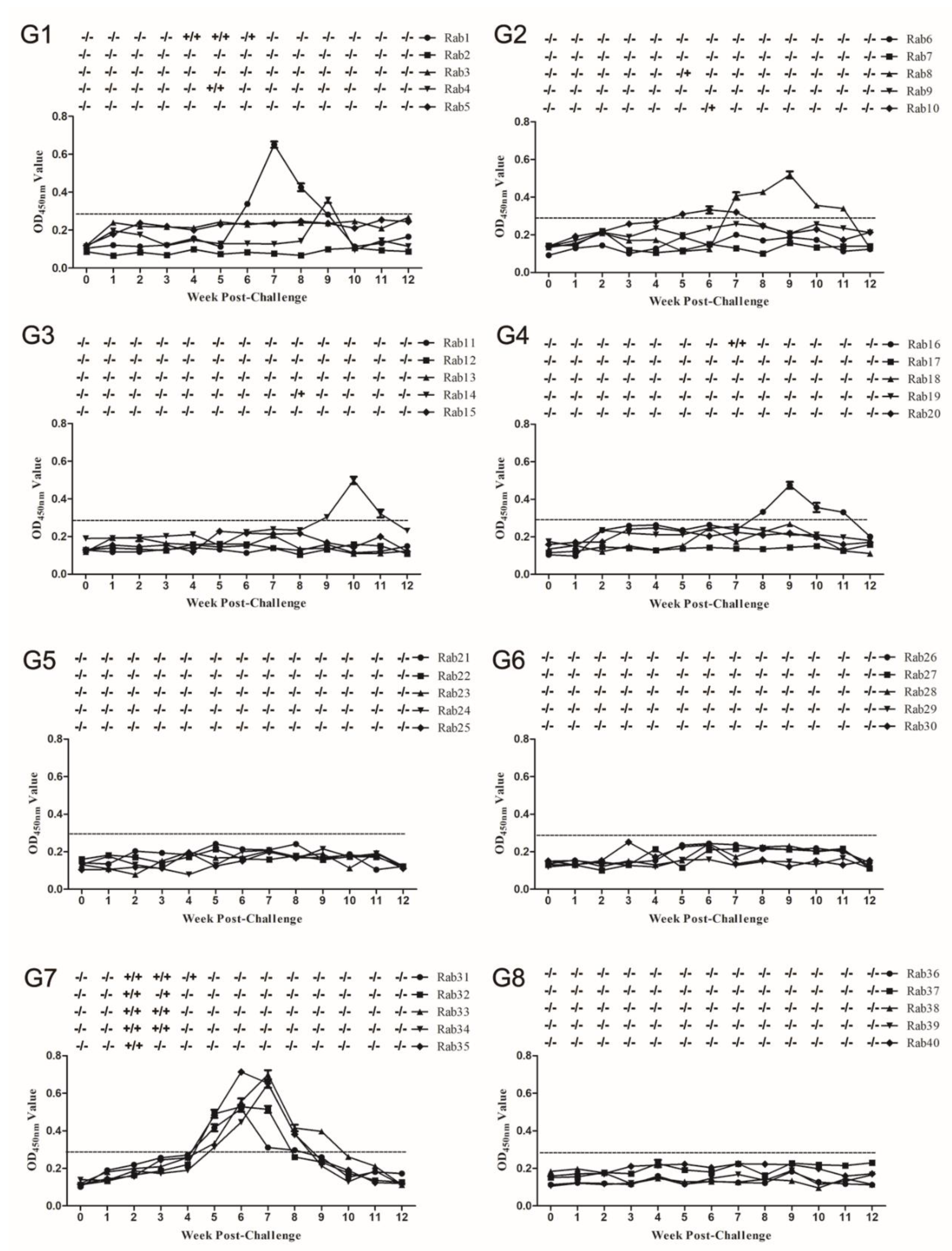
3.2. Measurements of Serum Antibody and HEV RNA Levels in Serum and Feces after Challenge
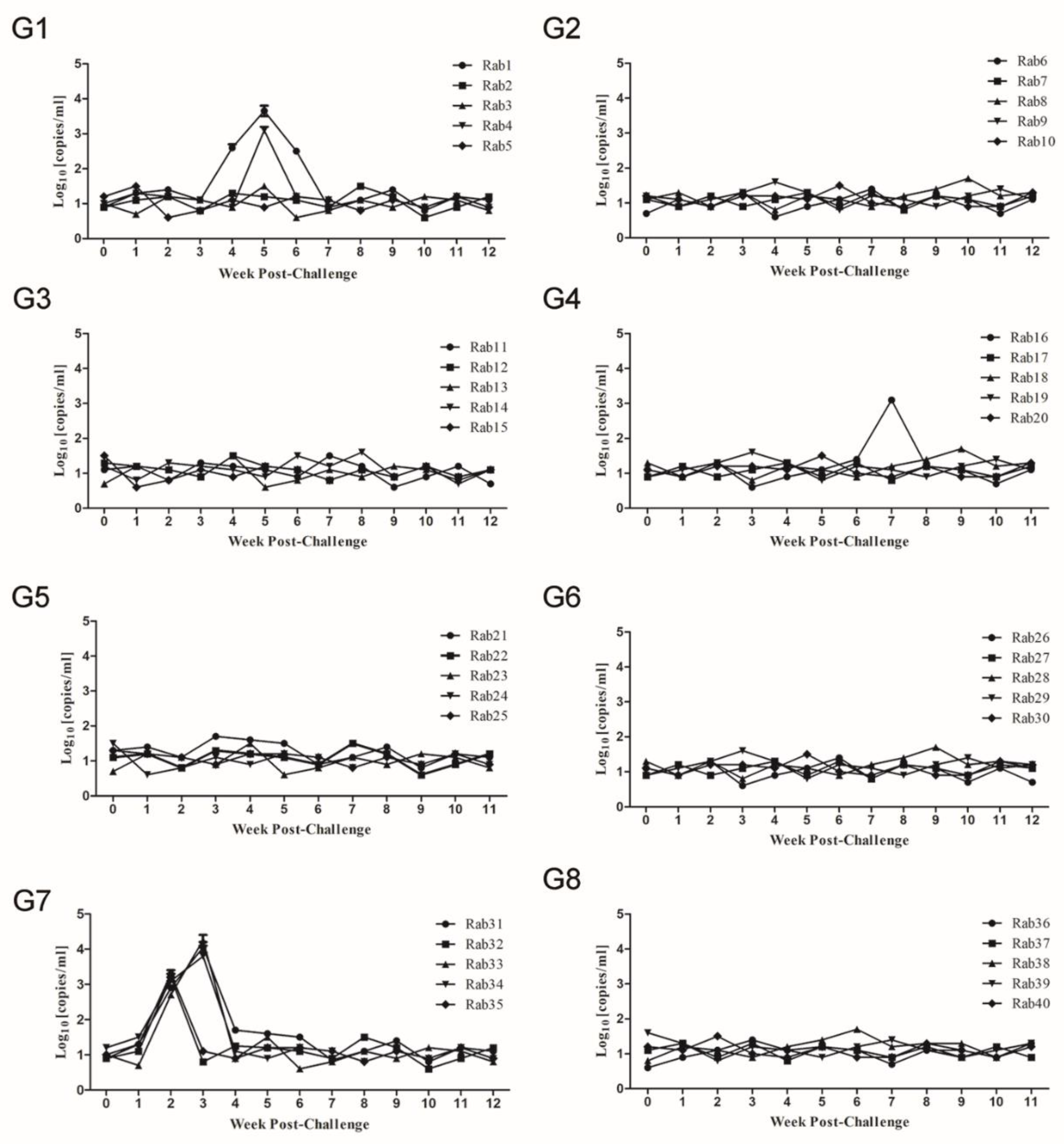
3.3. Quantitation of HEV RNA in Serum Samples
3.4. ALT evaluation in Serum Samples
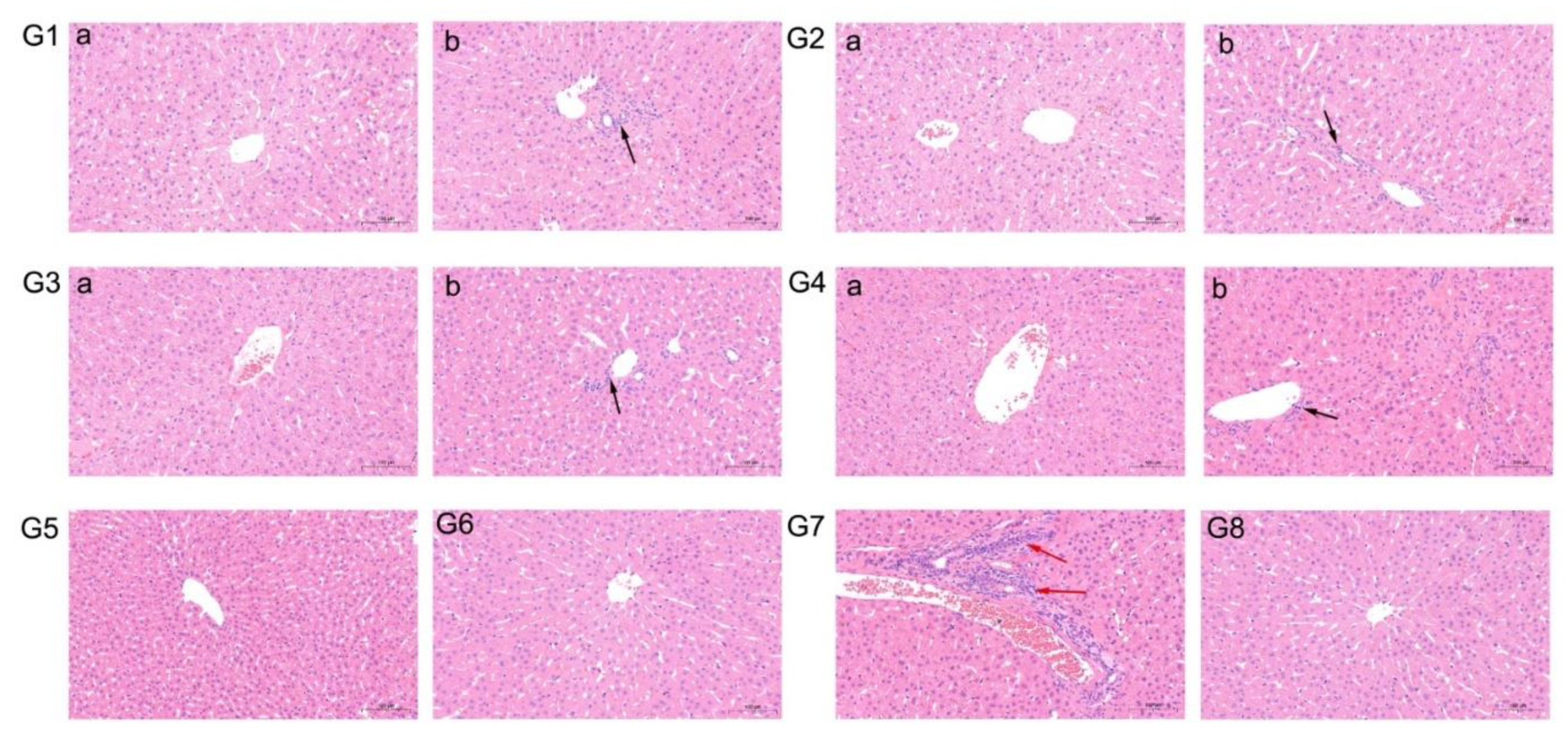
3.5. Gross and Microscopic Lesions
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Rein, D.B.; Stevens, G.A.; Theaker, J.; Wittenborn, J.S.; Wiersma, S.T. The global burden of hepatitis E virus genotypes 1 and 2 in 2005. Hepatology 2012, 55, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Labrique, A.B.; Sikder, S.S.; Krain, L.J.; West, K.P., Jr.; Christian, P.; Rashid, M.; Nelson, K.E. Hepatitis E, a vaccine-preventable cause of maternal deaths. Emerg. Infect. Dis. 2012, 18, 1401–1404. [Google Scholar] [CrossRef] [PubMed]
- Jameel, S. Molecular biology and pathogenesis of hepatitis E virus. Expert Rev. Mol. Med. 1999, 1999, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Tedder, R.S.; Ijaz, S. Persistent carriage of hepatitis E virus in patients with HIV infection. N. Engl. J. Med. 2009, 361, 1025–1027. [Google Scholar] [CrossRef] [PubMed]
- Ollier, L.; Tieulie, N.; Sanderson, F.; Heudier, P.; Giordanengo, V.; Fuzibet, J.G.; Nicand, E. Chronic hepatitis after hepatitis E virus infection in a patient with non-Hodgkin lymphoma taking rituxiMAb. Ann. Intern. Med. 2009, 150, 430–431. [Google Scholar] [CrossRef]
- Wedemeyer, H.; Pischke, S.; Manns, M.P. Pathogenesis and treatment of hepatitis e virus infection. Gastroenterology 2012, 142, 1388–1397. [Google Scholar] [CrossRef]
- Kamar, N.; Bendall, R.; Legrand-Abravanel, F.; Xia, N.S.; Ijaz, S.; Izopet, J.; Dalton, H.R. Hepatitis E. Lancet 2012, 379, 2477–2488. [Google Scholar] [CrossRef]
- Perez-Gracia, M.T.; Garcia, M.; Suay, B.; Mateos-Lindemann, M.L. Current Knowledge on Hepatitis E. J. Clin. Transl. Hepatol. 2015, 3, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Doceul, V.; Bagdassarian, E.; Demange, A.; Pavio, N. Zoonotic Hepatitis E Virus: Classi fi cation, Animal Reservoirs and Transmission Routes. Viruses 2016, 8, 270. [Google Scholar] [CrossRef]
- Liu, P.; Li, L.; Wang, L.; Bu, Q.; Fu, H.; Han, J.; Zhu, Y.; Lu, F.; Zhuang, H. Phylogenetic analysis of 626 hepatitis E virus (HEV) isolates from humans and animals in China (1986–2011) showing genotype diversity and zoonotic transmission. Infect. Genet. Evol. J. Mol. Epidemiol. Evolut. Genet. Infect. Dis. 2012, 12, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Lo, S.K.; Xing, F.; Yang, J.; Ye, H.; Chan, J.F.; Teng, J.L.; Huang, C.; Yip, C.C.; Lau, S.K.; et al. Clinical characteristics and molecular epidemiology of hepatitis E in Shenzhen, China: A shift toward foodborne transmission of hepatitis E virus infection. Emerg. Microbes Infect. 2017, 6, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 1997, 94, 9860–9865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arends, J.E.; Ghisetti, V.; Irving, W.; Dalton, H.R.; Izopet, J.; Hoepelman, A.I.; Salmon, D. Hepatitis E: An emerging infection in high income countries. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2014, 59, 81–88. [Google Scholar] [CrossRef]
- Ahmad, I.; Holla, R.P.; Jameel, S. Molecular virology of hepatitis E virus. Virus Res. 2011, 161, 47–58. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Q.; Dang, L.; Sun, Y.; Gao, J.; Liu, B.; Syed, S.F.; Tao, H.; Zhang, G.; Luo, J.; et al. Characterization of Two Novel Linear B-Cell Epitopes in the Capsid Protein of Avian Hepatitis E Virus (HEV) That Are Common to Avian, Swine, and Human HEVs. J. Virol. 2015, 89, 5491–5501. [Google Scholar] [CrossRef] [Green Version]
- Emerson, S.U.; Nguyen, H.T.; Torian, U.; Mather, K.; Firth, A.E. An essential RNA element resides in a central region of hepatitis E virus ORF2. J. Gen. Virol. 2013, 94, 1468–1476. [Google Scholar] [CrossRef]
- Zhao, M.; Li, X.J.; Tang, Z.M.; Yang, F.; Wang, S.L.; Cai, W.; Zhang, K.; Xia, N.S.; Zheng, Z.Z. A Comprehensive Study of Neutralizing Antigenic Sites on the Hepatitis E Virus (HEV) Capsid by Constructing, Clustering, and Characterizing a Tool Box. J. Biol. Chem. 2015, 290, 19910–19922. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Tang, X.; Seetharaman, J.; Yang, C.; Gu, Y.; Zhang, J.; Du, H.; Shih, J.W.; Hew, C.L.; Sivaraman, J.; et al. Dimerization of hepatitis E virus capsid protein E2s domain is essential for virus-host interaction. PLoS Pathog. 2009, 5, e1000537. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Yang, C.; Gu, Y.; Song, C.; Zhang, X.; Wang, Y.; Zhang, J.; Hew, C.L.; Li, S.; Xia, N.; et al. Structural basis for the neutralization and genotype specificity of hepatitis E virus. Proc. Natl. Acad. Sci. USA 2011, 108, 10266–10271. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Tang, X.; Zhang, X.; Song, C.; Zheng, M.; Wang, K.; Zhang, J.; Ng, M.H.; Hew, C.L.; Li, S.; et al. Structural basis for the neutralization of hepatitis E virus by a cross-genotype antibody. Cell Res. 2015, 25, 604–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Liu, B.; Sun, Y.; Li, H.; Du, T.; Nan, Y.; Hiscox, J.A.; Zhou, E.M.; Zhao, Q. Characterization of Three Novel Linear Neutralizing B-Cell Epitopes in the Capsid Protein of Swine Hepatitis E Virus. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Lei, Y.; Liu, L.; Liu, P.; Xia, J.; Zhang, Y.; Zeng, H.; Wang, L.; Wang, L.; Zhuang, H. SPF rabbits infected with rabbit hepatitis E virus isolate experimentally showing the chronicity of hepatitis. PLoS ONE 2014, 9, e99861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; An, J.; She, R.; Shi, R.; Hao, W.; Soomro, M.; Yuan, X.; Yang, J.; Wang, J. Detection of Genotype 4 Swine Hepatitis E Virus in Systemic Tissues in Cross-Species Infected Rabbits. PLoS ONE 2017, 12, e0171277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Chen, Y.; Sun, Y.; Nan, Y.; Li, H.; Du, T.; Hiscox, J.A.; Zhao, Q.; Zhou, E.M. Experimental infection of rabbit with swine-derived hepatitis E virus genotype 4. Vet. Microbiol. 2019, 229, 168–175. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Wang, L. An overview: Rabbit hepatitis E virus (HEV) and rabbit providing an animal model for HEV study. Rev. Med. Virol. 2018, 28. [Google Scholar] [CrossRef]
- Zhang, J.; Shih, J.W.; Xia, N.S. Long-term efficacy of a hepatitis E vaccine. N. Engl. J. Med. 2015, 372, 2265–2266. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wei, M.; Pan, H.; Lin, Z.; Wang, K.; Weng, Z.; Zhu, Y.; Xin, L.; Zhang, J.; Li, S.; et al. Robust manufacturing and comprehensive characterization of recombinant hepatitis E virus-like particles in Hecolin((R)). Vaccine 2014, 32, 4039–4050. [Google Scholar] [CrossRef]
- Huang, F.F.; Haqshenas, G.; Guenette, D.K.; Halbur, P.G.; Schommer, S.K.; Pierson, F.W.; Toth, T.E.; Meng, X.J. Detection by reverse transcription-PCR and genetic characterization of field isolates of swine hepatitis E virus from pigs in different geographic regions of the United States. J. Clin. Microbiol. 2002, 40, 1326–1332. [Google Scholar] [CrossRef] [Green Version]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Ma, H.; Zheng, L.; Liu, Y.; Zhao, C.; Harrison, T.J.; Ma, Y.; Sun, S.; Zhang, J.; Wang, Y. Experimental infection of rabbits with rabbit and genotypes 1 and 4 hepatitis E viruses. PLoS ONE 2010, 5, e9160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Zhou, E.M.; Sun, Z.F.; Meng, X.J. Protection of chickens against avian hepatitis E virus (avian HEV) infection by immunization with recombinant avian HEV capsid protein. Vaccine 2007, 25, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J. Hepatitis E virus: Animal reservoirs and zoonotic risk. Vet. Microbiol. 2010, 140, 256–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, H. Hepatitis E virus cell culture models. Virus Res. 2011, 161, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Purdy, M.A.; McCaustland, K.A.; Krawczynski, K.; Spelbring, J.; Reyes, G.R.; Bradley, D.W. Preliminary evidence that a trpE-HEV fusion protein protects cynomolgus macaques against challenge with wild-type hepatitis E virus (HEV). J. Med. Virol. 1993, 41, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Dai, X.; Chang, J.C.; Lopareva, E.; Pillot, J.; Fields, H.A.; Khudyakov, Y.E. Identification and characterization of the neutralization epitope(s) of the hepatitis E virus. Virology 2001, 288, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.M.; Tang, M.; Zhao, M.; Wen, G.P.; Yang, F.; Cai, W.; Wang, S.L.; Zheng, Z.Z.; Xia, N.S. A novel linear neutralizing epitope of hepatitis E virus. Vaccine 2015, 33, 3504–3511. [Google Scholar] [CrossRef]
- Sun, Y.; Lu, Q.; Liu, B.; Sheng, Y.; Du, T.; Hiscox, J.A.; Zhou, E.M.; Zhao, Q. Cross-species infection of mice by rabbit hepatitis E virus. Vet. Microbiol. 2018, 225, 48–52. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Immunization | Seroconversion to Rabbit HEV | Fecal Virus Shedding | Viremia | ALT Increased | Microscopic Lesions |
|---|---|---|---|---|---|---|
| 1 | VKLYTS | 4/5 | 2/5 | 2/5 | 2/5 | 2/5 |
| 2 | PSRPF | 3/5 | 3/5 | 0/5 | 2/5 | 2/5 |
| 3 | EPTV | 3/5 | 1/5 | 0/5 | 1/5 | 1/5 |
| 4 | EPTVKLYTS | 1/5 | 1/5 | 1/5 | 1/5 | 1/5 |
| 5 | EPTVKLYTSPSRPF | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 |
| 6 | Sp239 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 |
| 7 | PBS (NC) | 5/5 | 5/5 | 5/5 | 5/5 | 5/5 |
| 8 | PBS (Normal) | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Chen, T.; Luo, Y.; Fan, J.; Zhang, M.; Zhao, Q.; Nan, Y.; Liu, B.; Zhou, E.-M. Synthetic Peptides Containing Three Neutralizing Epitopes of Genotype 4 Swine Hepatitis E Virus ORF2 induced Protection against Swine HEV Infection in Rabbit. Vaccines 2020, 8, 178. https://doi.org/10.3390/vaccines8020178
Chen Y, Chen T, Luo Y, Fan J, Zhang M, Zhao Q, Nan Y, Liu B, Zhou E-M. Synthetic Peptides Containing Three Neutralizing Epitopes of Genotype 4 Swine Hepatitis E Virus ORF2 induced Protection against Swine HEV Infection in Rabbit. Vaccines. 2020; 8(2):178. https://doi.org/10.3390/vaccines8020178
Chicago/Turabian StyleChen, Yiyang, Tianxiang Chen, Yuhang Luo, Jie Fan, Meimei Zhang, Qin Zhao, Yuchen Nan, Baoyuan Liu, and En-Min Zhou. 2020. "Synthetic Peptides Containing Three Neutralizing Epitopes of Genotype 4 Swine Hepatitis E Virus ORF2 induced Protection against Swine HEV Infection in Rabbit" Vaccines 8, no. 2: 178. https://doi.org/10.3390/vaccines8020178
APA StyleChen, Y., Chen, T., Luo, Y., Fan, J., Zhang, M., Zhao, Q., Nan, Y., Liu, B., & Zhou, E.-M. (2020). Synthetic Peptides Containing Three Neutralizing Epitopes of Genotype 4 Swine Hepatitis E Virus ORF2 induced Protection against Swine HEV Infection in Rabbit. Vaccines, 8(2), 178. https://doi.org/10.3390/vaccines8020178