Sendai Virus, a Strong Inducer of Anti-Lentiviral State in Ovine Cells

 , , , and
, , , and 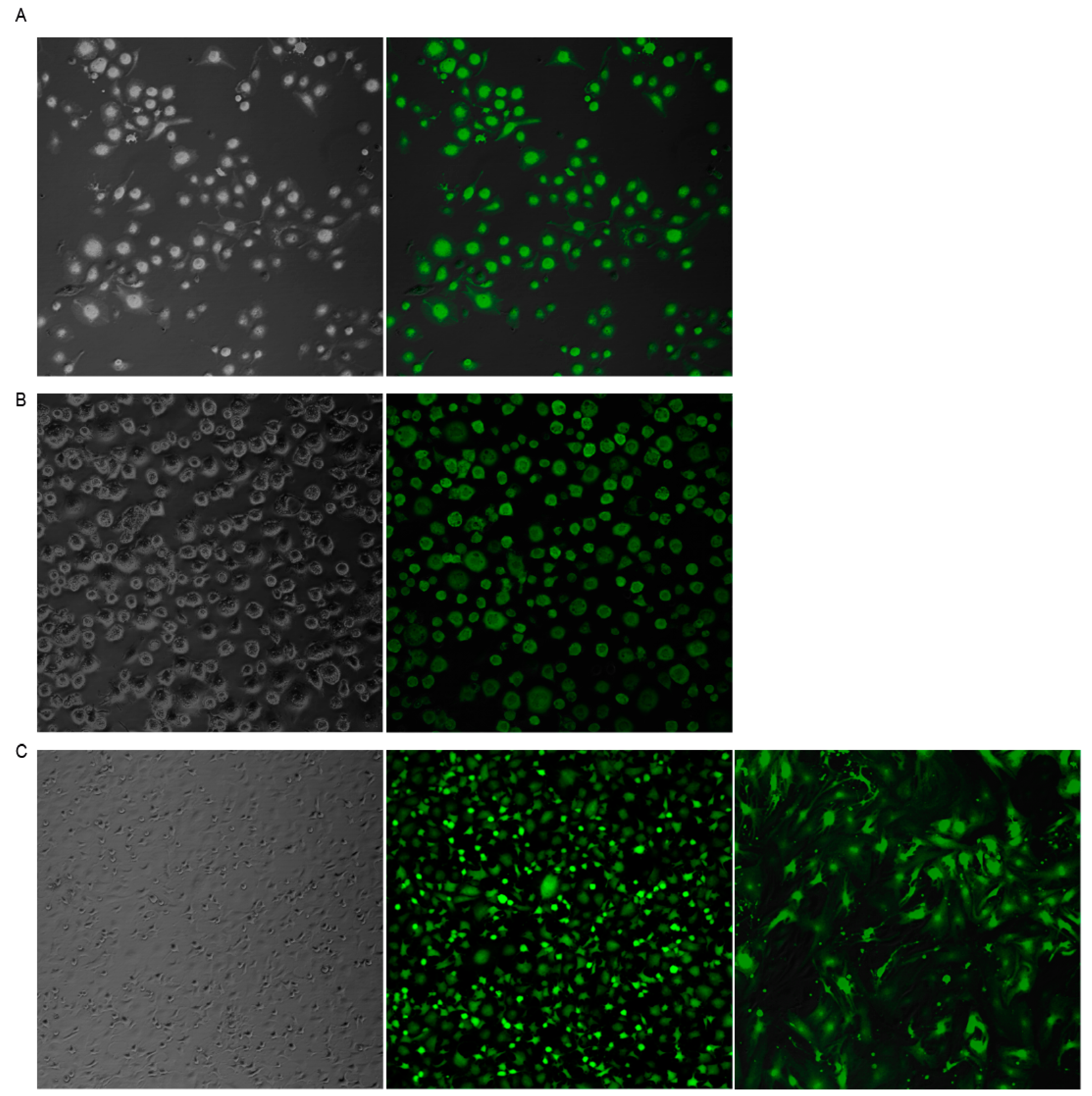{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Cell Infection and Virus Quantification
2.3. mRNA Relative Quantification
2.4. Type-I IFN Bioassay
2.5. Statistical Analysis
3. Results
3.1. SeV Infection Is Highly Efficient in Ovine Cells
3.2. SeV Infection Induced Stable GFP Expression in Ovine Cells
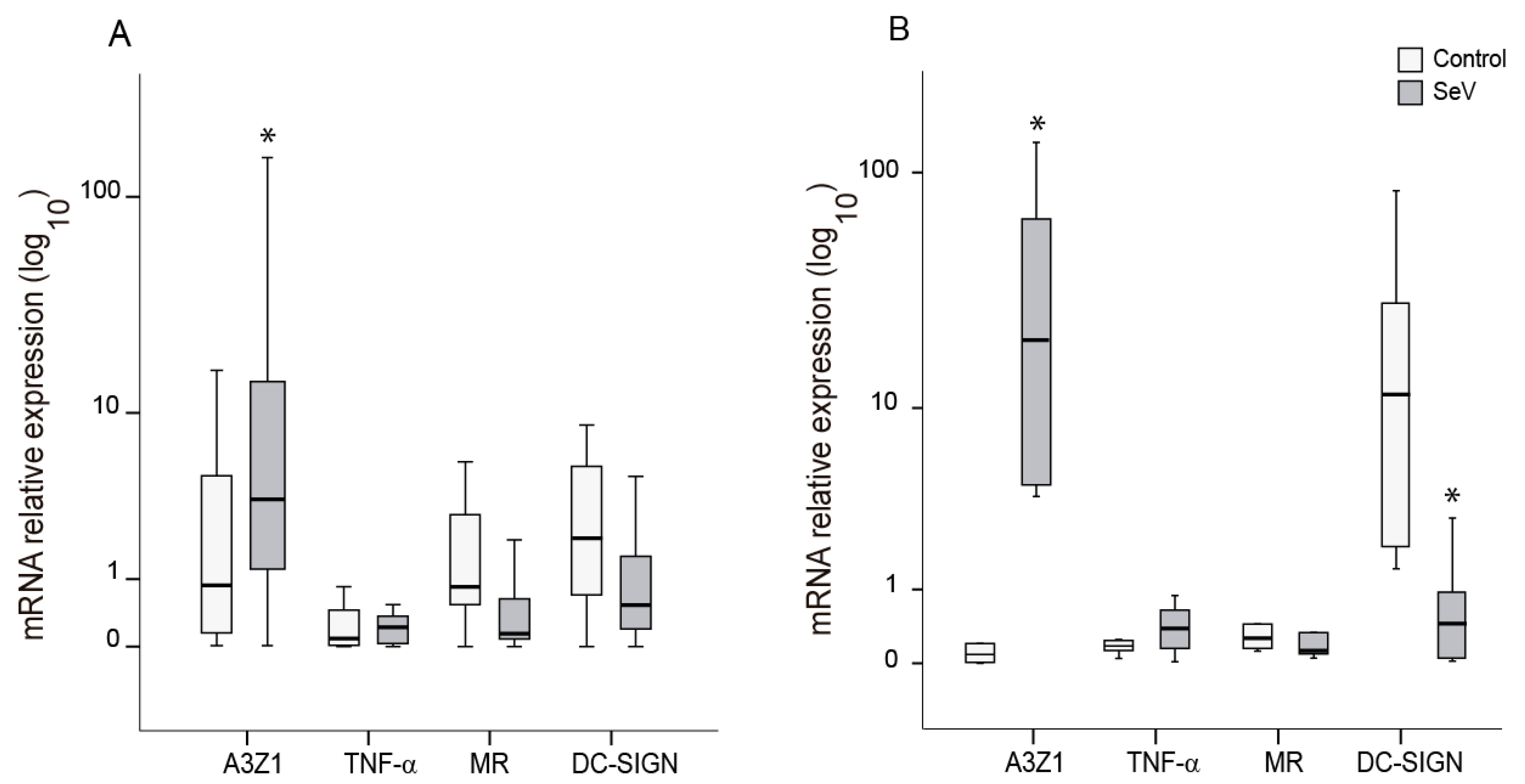
3.3. SeV Infection Induces Proinflammatory Responses in Ovine Cells
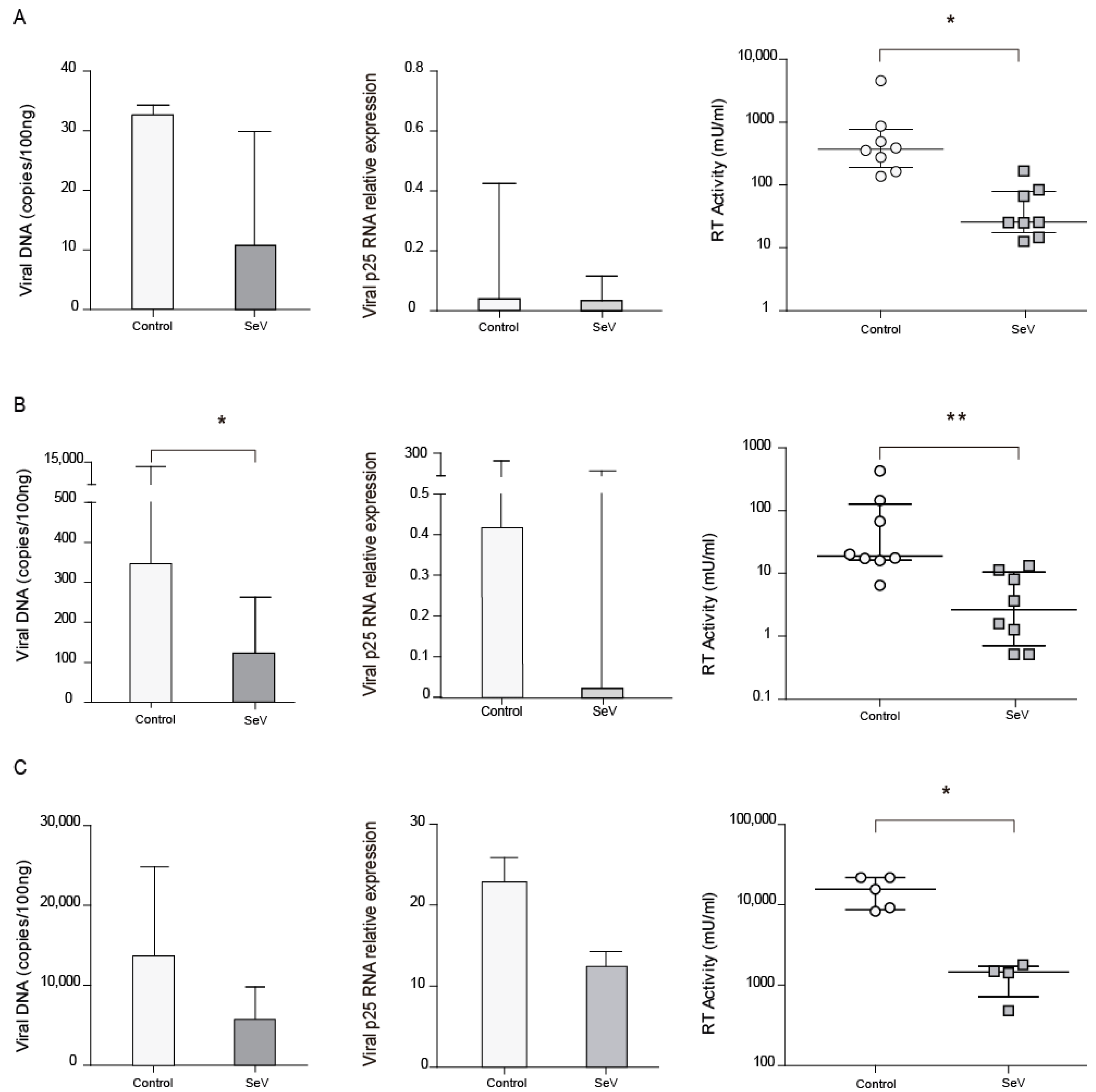
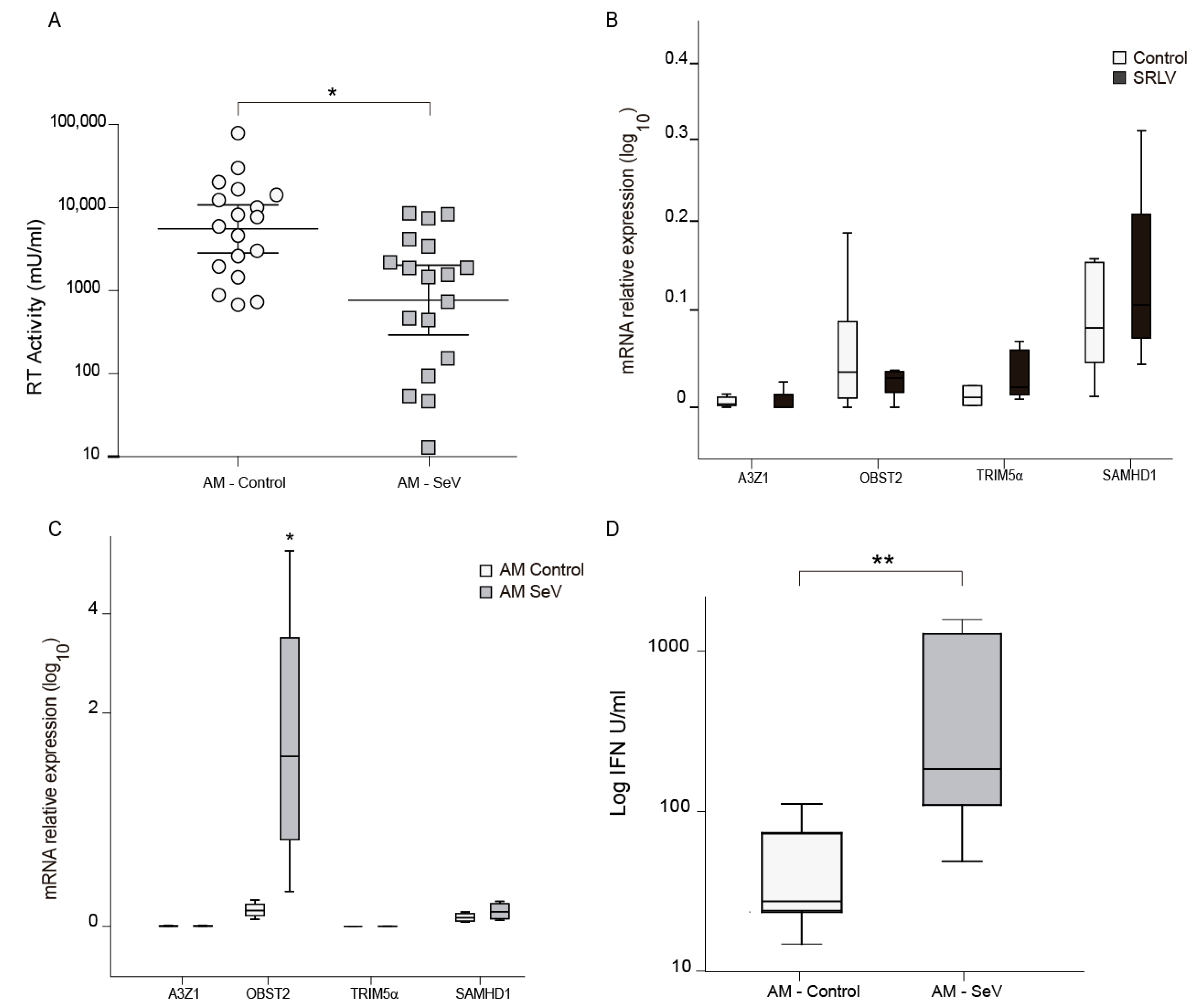
3.4. SeV-Infected Cells Reduced Permissibility to SRLV Infection
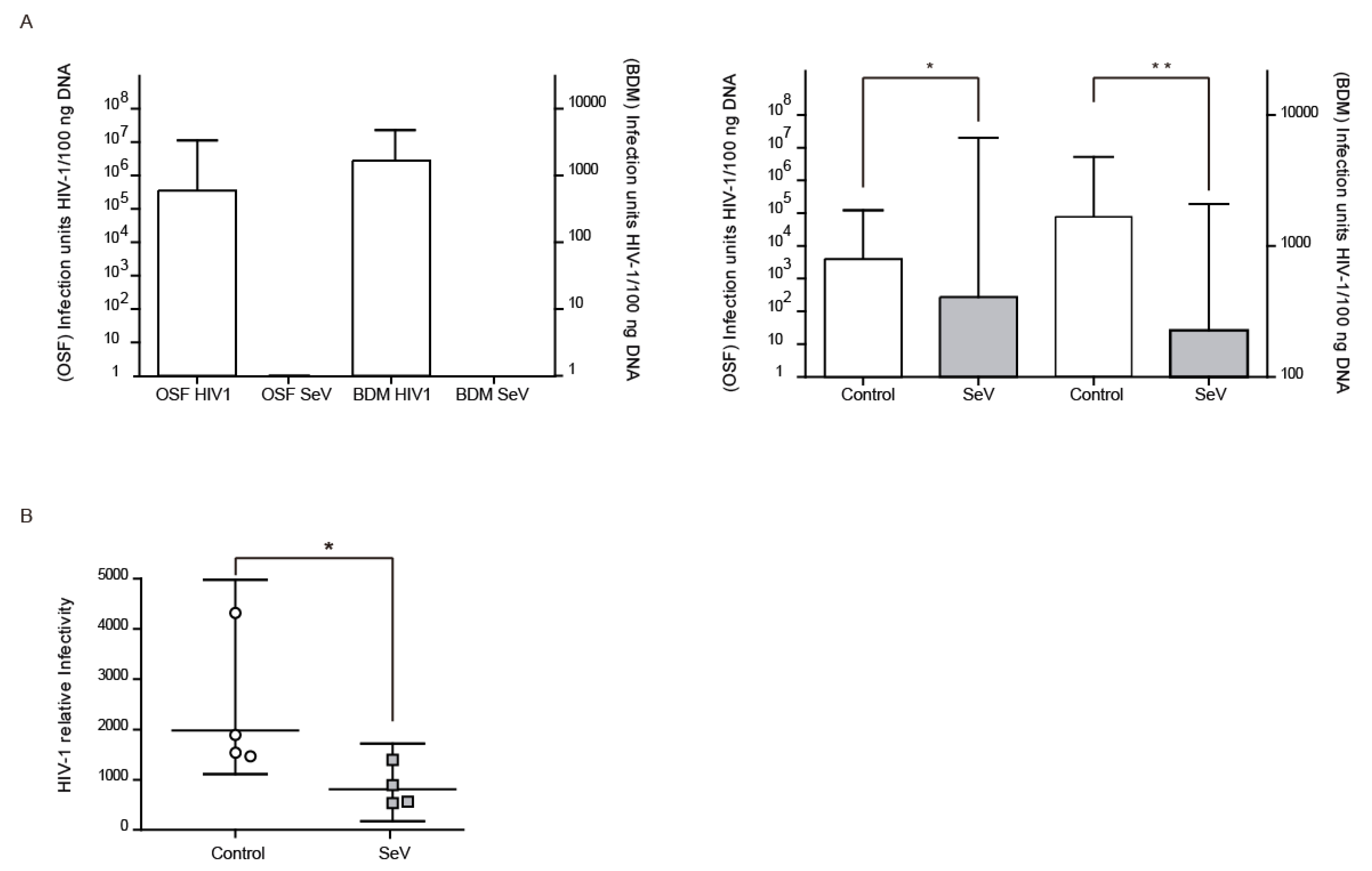
3.5. Ovine Cells Infected with SeV-GFP Inhibit HIV-1-GFP Vector Infectivity
3.6. Restriction Factors Induced after SeV Infection in Ovine Cells
3.7. SeV Infection May Induce Local Resistance to SRLV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ballandras-Colas, A.; Maskell, D.P.; Serrao, E.; Locke, J.; Swuec, P.; Jónsson, S.R.; Kotecha, A.; Cook, N.J.; Pye, V.E.; Taylor, I.A.; et al. A supramolecular assembly mediates lentiviral DNA integration. Science 2017, 355, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Mselli-Lakhal, L.; Guiguen, F.; Greenland, T.; Mornex, J.F.; Chebloune, Y. Gene transfer system derived from the caprine arthritis encephalitis lentivirus. J. Virol. Methods 2006, 136, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Reina, R.; de Andres, D.; Amorena, B. Immunization against small ruminant lentiviruses. Viruses 2013, 5, 1948–1963. [Google Scholar] [CrossRef] [PubMed]
- Minguijon, E.; Reina, R.; Perez, M.; Polledo, L.; Villoria, M.; Ramirez, H.; Leginagoikoa, I.; Badiola, J.J.; Garcia-Marin, J.F.; de Andres, D.; et al. Small ruminant lentivirus infections and diseases. Vet. Microbiol. 2015, 181, 75–89. [Google Scholar] [CrossRef]
- Ritchie, C.; Hosie, B. Increase in maedi-visna breakdowns. Vet. Rec. 2010, 167, 389. [Google Scholar]
- Ritchie, C.; Hosie, B. Concern over maedi visna breakdowns. Vet. Rec. 2014, 175, 50–51. [Google Scholar] [CrossRef]
- Sattentau, Q.J.; Stevenson, M. Macrophages and HIV-1: An Unhealthy Constellation. Cell Host Microbe 2016, 19, 304–310. [Google Scholar] [CrossRef]
- Crespo, H.; Bertolotti, L.; Juganaru, M.; Glaria, I.; de Andres, D.; Amorena, B.; Rosati, S.; Reina, R. Small ruminant macrophage polarization may play a pivotal role on lentiviral infection. Vet. Res. 2013, 44, 83. [Google Scholar] [CrossRef]
- Cassetta, L.; Cassol, E.; Poli, G. Macrophage polarization in health and disease. Sci. World J. 2011, 11, 2391–2402. [Google Scholar] [CrossRef]
- Blacklaws, B.A. Small ruminant lentiviruses: Immunopathogenesis of visna-maedi and caprine arthritis and encephalitis virus. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 259–269. [Google Scholar] [CrossRef]
- Simon, V.; Bloch, N.; Landau, N.R. Intrinsic host restrictions to HIV-1 and mechanisms of viral escape. Nat. Immunol. 2015, 16, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Sironi, M.; Cagliani, R.; Forni, D.; Clerici, M. Evolutionary insights into host-pathogen interactions from mammalian sequence data. Nat. Rev. Genet. 2015, 16, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Ma, F.; Quinn, M.; Xiang, S.H. Genome-Wide Search for Host Association Factors during Ovine Progressive Pneumonia Virus Infection. PLoS ONE 2016, 11, e0150344. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Griesenbach, U.; Shiraki-Iida, T.; Shu, T.; Hironaka, T.; Hou, X.; Williams, J.; Zhu, J.; Jeffery, P.K.; Geddes, D.M.; et al. A defective nontransmissible recombinant Sendai virus mediates efficient gene transfer to airway epithelium in vivo. Gene Ther. 2004, 11, 1659–1664. [Google Scholar] [CrossRef]
- Ishii, K.J.; Koyama, S.; Nakagawa, A.; Coban, C.; Akira, S. Host innate immune receptors and beyond: Making sense of microbial infections. Cell Host Microbe 2008, 3, 352–363. [Google Scholar] [CrossRef]
- Seki, S.; Matano, T. Development of a Sendai virus vector-based AIDS vaccine inducing T cell responses. Expert Rev. Vaccines 2016, 15, 119–127. [Google Scholar] [CrossRef]
- Kamga, I.; Kahi, S.; Develioglu, L.; Lichtner, M.; Maranon, C.; Deveau, C.; Meyer, L.; Goujard, C.; Lebon, P.; Sinet, M.; et al. Type I interferon production is profoundly and transiently impaired in primary HIV-1 infection. J. Infect. Dis. 2005, 192, 303–310. [Google Scholar] [CrossRef]
- Nyombayire, J.; Anzala, O.; Gazzard, B.; Karita, E.; Bergin, P.; Hayes, P.; Kopycinski, J.; Omosa-Manyonyi, G.; Jackson, A.; Bizimana, J.; et al. First-in-Human Evaluation of the Safety and Immunogenicity of an Intranasally Administered Replication-Competent Sendai Virus-Vectored HIV Type 1 Gag Vaccine: Induction of Potent T-Cell or Antibody Responses in Prime-Boost Regimens. J. Infect. Dis. 2017, 215, 95–104. [Google Scholar] [CrossRef]
- Griesenbach, U.; McLachlan, G.; Owaki, T.; Somerton, L.; Shu, T.; Baker, A.; Tennant, P.; Gordon, C.; Vrettou, C.; Baker, E.; et al. Validation of recombinant Sendai virus in a non-natural host model. Gene Ther. 2011, 18, 182–188. [Google Scholar] [CrossRef]
- Glaria, I.; Reina, R.; Ramirez, H.; de Andres, X.; Crespo, H.; Jauregui, P.; Salazar, E.; Lujan, L.; Perez, M.M.; Benavides, J.; et al. Visna/Maedi virus genetic characterization and serological diagnosis of infection in sheep from a neurological outbreak. Vet. Microbiol. 2012, 155, 137–146. [Google Scholar] [CrossRef]
- Gonzalez, B.; Reina, R.; Garcia, I.; Andres, S.; Glaria, I.; Alzueta, M.; Mora, M.I.; Jugo, B.M.; Arrieta-Aguirre, I.; de la Lastra, J.M.; et al. Mucosal immunization of sheep with a Maedi-Visna virus (MVV) env DNA vaccine protects against early MVV productive infection. Vaccine 2005, 23, 4342–4352. [Google Scholar] [CrossRef] [PubMed]
- Adler, H.; Frech, B.; Thony, M.; Pfister, H.; Peterhans, E.; Jungi, T.W. Inducible nitric oxide synthase in cattle. Differential cytokine regulation of nitric oxide synthase in bovine and murine macrophages. J. Immunol. 1995, 154, 4710–4718. [Google Scholar]
- Bird, P.; Blacklaws, B.; Reyburn, H.T.; Allen, D.; Hopkins, J.; Sargan, D.; McConnell, I. Early events in immune evasion by the lentivirus maedi-visna occurring within infected lymphoid tissue. J. Virol. 1993, 67, 5187–5197. [Google Scholar] [CrossRef] [PubMed]
- Sargan, D.R.; Bennet, I.D.; Cousens, C.; Roy, D.J.; Blacklaws, B.A.; Dalziel, R.G.; Watt, N.J.; McConnell, I. Nucleotide sequence of EV1, a British isolate of maedi-visna virus. J. Gen. Virol. 1991, 72 Pt 8, 1893–1903. [Google Scholar] [CrossRef]
- Glaria, I.; Reina, R.; Crespo, H.; de Andres, X.; Ramirez, H.; Biescas, E.; Perez, M.M.; Badiola, J.; Lujan, L.; Amorena, B.; et al. Phylogenetic analysis of SRLV sequences from an arthritic sheep outbreak demonstrates the introduction of CAEV-like viruses among Spanish sheep. Vet. Microbiol. 2009, 138, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Per Cent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Strahle, L.; Garcin, D.; Kolakofsky, D. Sendai virus defective-interfering genomes and the activation of interferon-beta. Virology 2006, 351, 101–111. [Google Scholar] [CrossRef]
- Jauregui, P.; Crespo, H.; Glaria, I.; Lujan, L.; Contreras, A.; Rosati, S.; de Andres, D.; Amorena, B.; Towers, G.J.; Reina, R. Ovine TRIM5alpha can restrict visna/maedi virus. J. Virol. 2012, 86, 9504–9509. [Google Scholar] [CrossRef]
- Jacques, D.A.; McEwan, W.A.; Hilditch, L.; Price, A.J.; Towers, G.J.; James, L.C. HIV-1 uses dynamic capsid pores to import nucleotides and fuel encapsidated DNA synthesis. Nature 2016, 536, 349–353. [Google Scholar] [CrossRef]
- De Pablo-Maiso, L.; Glaria, I.; Crespo, H.; Nistal-Villan, E.; Andresdottir, V.; de Andres, D.; Amorena, B.; Reina, R. Characterization of Ovine A3Z1 Restriction Properties against Small Ruminant Lentiviruses (SRLVs). Viruses 2017, 9, 345. [Google Scholar] [CrossRef]
- Toledo-Arana, A.; Dussurget, O.; Nikitas, G.; Sesto, N.; Guet-Revillet, H.; Balestrino, D.; Loh, E.; Gripenland, J.; Tiensuu, T.; Vaitkevicius, K.; et al. The Listeria transcriptional landscape from saprophytism to virulence. Nature 2009, 459, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Vermeire, J.; Naessens, E.; Vanderstraeten, H.; Landi, A.; Iannucci, V.; Van Nuffel, A.; Taghon, T.; Pizzato, M.; Verhasselt, B. Quantification of reverse transcriptase activity by real-time PCR as a fast and accurate method for titration of HIV, lenti- and retroviral vectors. PLoS ONE 2012, 7, e50859. [Google Scholar] [CrossRef] [PubMed]
- Crespo, H.; Jauregui, P.; Glaria, I.; Sanjose, L.; Polledo, L.; Garcia-Marin, J.F.; Lujan, L.; de Andres, D.; Amorena, B.; Reina, R. Mannose receptor may be involved in small ruminant lentivirus pathogenesis. Vet. Res. 2012, 43, 43. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Rius-Rocabert, S.; Presa, J.L.; Esteban-Rubio, S.; Ayuso-Sacido, A.; Nistal-Villan, E. A Digital Method to Quantify Type I Interferon. J. Interferon Cytokine Res. 2019, 39, 711–719. [Google Scholar] [CrossRef]
- Olech, M.; Valas, S.; Kuzmak, J. Epidemiological survey in single-species flocks from Poland reveals expanded genetic and antigenic diversity of small ruminant lentiviruses. PLoS ONE 2018, 13, e0193892. [Google Scholar] [CrossRef]
- Reina, R.; Juganaru, M.M.; Profiti, M.; Cascio, P.; Cerruti, F.; Bertolotti, L.; De Meneghi, D.; Amorena, B.; Rosati, S. Immunological parameters in goats experimentally infected with SRLV genotype E, strain Roccaverano. Vet. Immunol. Immunopathol. 2011, 139, 237–244. [Google Scholar] [CrossRef]
- Skraban, R.; Matthiasdottir, S.; Torsteinsdottir, S.; Agnarsdottir, G.; Gudmundsson, B.; Georgsson, G.; Meloen, R.H.; Andresson, O.S.; Staskus, K.A.; Thormar, H.; et al. Naturally occurring mutations within 39 amino acids in the envelope glycoprotein of maedi-visna virus alter the neutralization phenotype. J. Virol. 1999, 73, 8064–8072. [Google Scholar] [CrossRef]
- Torsteinsdottir, S.; Andresdottir, V.; Arnarson, H.; Petursson, G. Immune response to maedi-visna virus. Front. Biosci. 2007, 12, 1532–1543. [Google Scholar] [CrossRef]
- Reina, R.; Berriatua, E.; Lujan, L.; Juste, R.; Sanchez, A.; de Andres, D.; Amorena, B. Prevention strategies against small ruminant lentiviruses: An update. Vet. J. 2009, 182, 31–37. [Google Scholar] [CrossRef]
- Zhang, X.; Edwards, J.P.; Mosser, D.M. The expression of exogenous genes in macrophages: Obstacles and opportunities. Methods Mol. Biol. 2009, 531, 123–143. [Google Scholar] [PubMed]
- Villar, E.; Barroso, I.M. Role of sialic acid-containing molecules in paramyxovirus entry into the host cell: A minireview. Glycoconj. J. 2006, 23, 5–17. [Google Scholar] [CrossRef]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Eltahir, Y.M.; Dovas, C.I.; Papanastassopoulou, M.; Koumbati, M.; Giadinis, N.; Verghese-Nikolakaki, S.; Koptopoulos, G. Development of a semi-nested PCR using degenerate primers for the generic detection of small ruminant lentivirus proviral DNA. J. Virol. Methods 2006, 135, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Stavrou, S.; Crawford, D.; Blouch, K.; Browne, E.P.; Kohli, R.M.; Ross, S.R. Different modes of retrovirus restriction by human APOBEC3A and APOBEC3G in vivo. PLoS Pathog. 2014, 10, e1004145. [Google Scholar] [CrossRef]
- Franzolin, E.; Pontarin, G.; Rampazzo, C.; Miazzi, C.; Ferraro, P.; Palumbo, E.; Reichard, P.; Bianchi, V. The deoxynucleotide triphosphohydrolase SAMHD1 is a major regulator of DNA precursor pools in mammalian cells. Proc. Natl. Acad. Sci. USA 2013, 110, 14272–14277. [Google Scholar] [CrossRef]
- Chen, S.; Bonifati, S.; Qin, Z.; St Gelais, C.; Kodigepalli, K.M.; Barrett, B.S.; Kim, S.H.; Antonucci, J.M.; Ladner, K.J.; Buzovetsky, O.; et al. SAMHD1 suppresses innate immune responses to viral infections and inflammatory stimuli by inhibiting the NF-kappaB and interferon pathways. Proc. Natl. Acad. Sci. USA 2018, 115, E3798–E3807. [Google Scholar] [CrossRef]
- Dufrasne, F.E.; Lucchetti, M.; Martin, A.; Andre, E.; Dessilly, G.; Kabamba, B.; Goubau, P.; Ruelle, J. Modulation of the NF-kappaB signaling pathway by the HIV-2 envelope glycoprotein and its incomplete BST-2 antagonism. Virology 2018, 513, 11–16. [Google Scholar] [CrossRef]
- Crespo, H.; Bertolotti, L.; Proffiti, M.; Cascio, P.; Cerruti, F.; Acutis, P.L.; de Andres, D.; Reina, R.; Rosati, S. Low proviral small ruminant lentivirus load as biomarker of natural restriction in goats. Vet. Microbiol. 2016, 192, 152–162. [Google Scholar] [CrossRef]
- Murphy, B.; McElliott, V.; Vapniarsky, N.; Oliver, A.; Rowe, J. Tissue tropism and promoter sequence variation in caprine arthritis encephalitis virus infected goats. Virus Res. 2010, 151, 177–184. [Google Scholar] [CrossRef]
- Herrmann-Hoesing, L.M.; Noh, S.M.; Snekvik, K.R.; White, S.N.; Schneider, D.A.; Truscott, T.; Knowles, D.P. Ovine progressive pneumonia virus capsid antigen as found in CD163- and CD172a-positive alveolar macrophages of persistently infected sheep. Vet. Pathol. 2010, 47, 518–528. [Google Scholar] [CrossRef] [PubMed]
- White, S.N.; Mousel, M.R.; Reynolds, J.O.; Lewis, G.S.; Herrmann-Hoesing, L.M. Common promoter deletion is associated with 3.9-fold differential transcription of ovine CCR5 and reduced proviral level of ovine progressive pneumonia virus. Anim. Genet. 2009, 40, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, J.; Holic, N.; Martinez, K.; Danos, O.; Perea, J. A high throughput method for genome-wide analysis of retroviral integration. Nucleic Acids Res. 2006, 34, e134. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fournier, P.; Zeng, J.; Schirrmacher, V. Two ways to induce innate immune responses in human PBMCs: Paracrine stimulation of IFN-alpha responses by viral protein or dsRNA. Int. J. Oncol. 2003, 23, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Pollara, G.; Jones, M.; Handley, M.E.; Rajpopat, M.; Kwan, A.; Coffin, R.S.; Foster, G.; Chain, B.; Katz, D.R. Herpes simplex virus type-1-induced activation of myeloid dendritic cells: The roles of virus cell interaction and paracrine type I IFN secretion. J. Immunol. 2004, 173, 4108–4119. [Google Scholar] [CrossRef] [PubMed]
- Griesenbach, U.; Boyton, R.J.; Somerton, L.; Garcia, S.E.; Ferrari, S.; Owaki, T.; Ya-Fen, Z.; Geddes, D.M.; Hasegawa, M.; Altmann, D.M.; et al. Effect of tolerance induction to immunodominant T-cell epitopes of Sendai virus on gene expression following repeat administration to lung. Gene Ther. 2006, 13, 449–456. [Google Scholar] [CrossRef]
- Komaru, A.; Ueda, Y.; Furuya, A.; Tanaka, S.; Yoshida, K.; Kato, T.; Kinoh, H.; Harada, Y.; Suzuki, H.; Inoue, M.; et al. Sustained and NK/CD4+ T cell-dependent efficient prevention of lung metastasis induced by dendritic cells harboring recombinant Sendai virus. J. Immunol. 2009, 183, 4211–4219. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Pablo-Maiso, L.; Echeverría, I.; Rius-Rocabert, S.; Luján, L.; Garcin, D.; de Andrés, D.; Nistal-Villán, E.; Reina, R. Sendai Virus, a Strong Inducer of Anti-Lentiviral State in Ovine Cells. Vaccines 2020, 8, 206. https://doi.org/10.3390/vaccines8020206
de Pablo-Maiso L, Echeverría I, Rius-Rocabert S, Luján L, Garcin D, de Andrés D, Nistal-Villán E, Reina R. Sendai Virus, a Strong Inducer of Anti-Lentiviral State in Ovine Cells. Vaccines. 2020; 8(2):206. https://doi.org/10.3390/vaccines8020206
Chicago/Turabian Stylede Pablo-Maiso, Lorena, Irache Echeverría, Sergio Rius-Rocabert, Lluís Luján, Dominique Garcin, Damián de Andrés, Estanislao Nistal-Villán, and Ramsés Reina. 2020. "Sendai Virus, a Strong Inducer of Anti-Lentiviral State in Ovine Cells" Vaccines 8, no. 2: 206. https://doi.org/10.3390/vaccines8020206
APA Stylede Pablo-Maiso, L., Echeverría, I., Rius-Rocabert, S., Luján, L., Garcin, D., de Andrés, D., Nistal-Villán, E., & Reina, R. (2020). Sendai Virus, a Strong Inducer of Anti-Lentiviral State in Ovine Cells. Vaccines, 8(2), 206. https://doi.org/10.3390/vaccines8020206