Acellular Pertussis Vaccine Components: Today and Tomorrow



Abstract
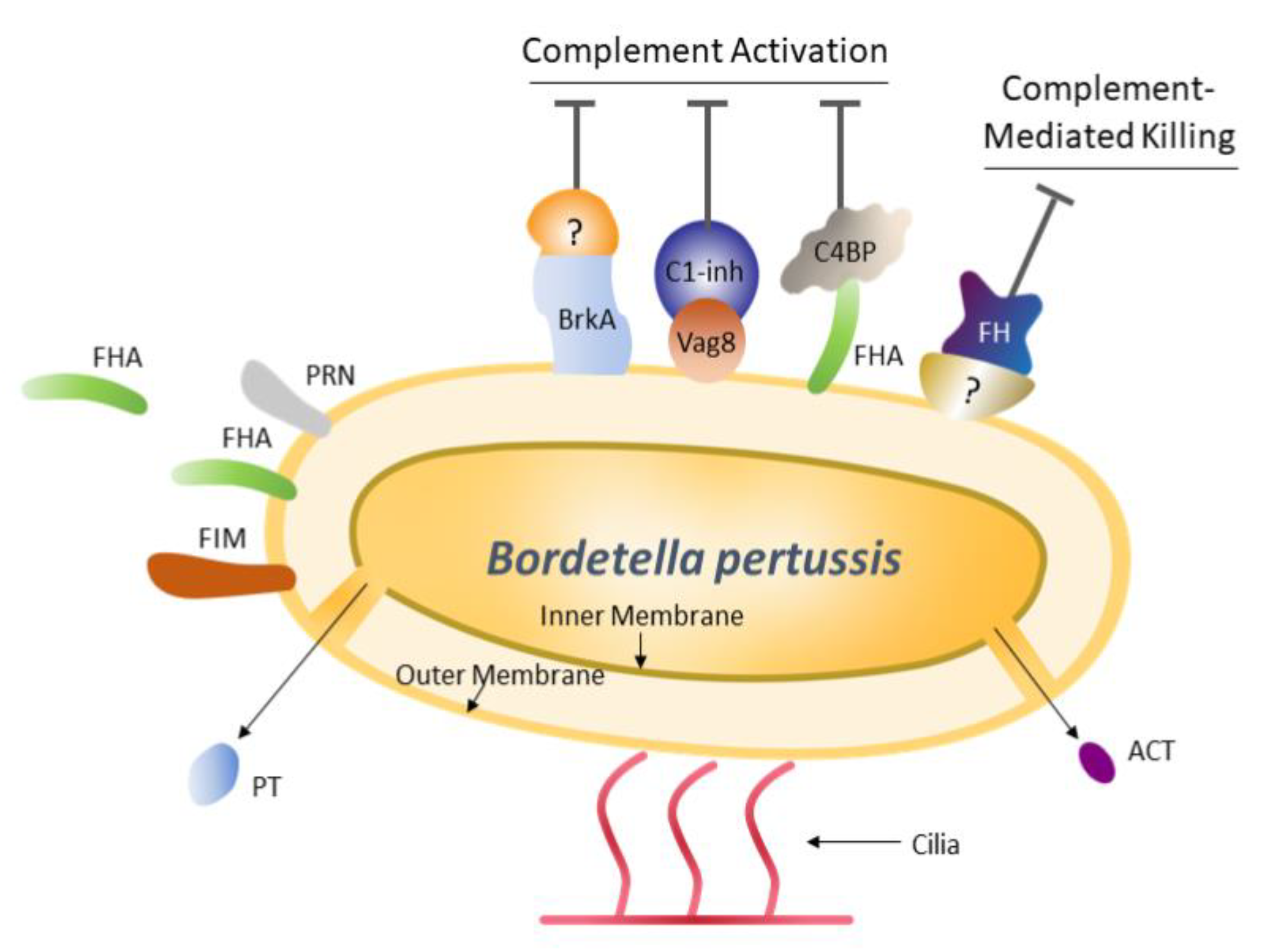
:1. Introduction
2. Pertussis Antigens in Acellular Vaccines
2.1. Pertussis Toxin
2.2. Filamentous Hemagglutinin
2.3. Pertactin
2.4. Fimbriae Types 2 and 3
3. Value of Multiple Antigens in Acellular Vaccines
4. Potential Correlates of Protection for Pertussis
5. Genetic Changes Attributed to Vaccines
5.1. Pertactin Loss
5.2. Pertussis Toxin Overexpression
6. Novel Vaccine Strategies
6.1. Adenylate Cyclase Toxin
6.2. Outer Membrane Vesicles
6.3. Novel Adjuvants
6.4. Live Attenuated, Nasally Administered Vaccines
7. Regulatory Considerations for Novel Acellular Vaccines
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Pertussis. In Epidemiology and Prevention of Vaccine-Preventable Diseases, 13th ed.; Hamborsky, J., Kroger, A., Eds.; Public Health Foundation: Washington, DC, USA, 2015. [Google Scholar]
- World Health Organization. Pertussis. Available online: https://www.who.int/immunization/monitoring_surveillance/burden/vpd/surveillance_type/passive/pertussis/en/ (accessed on 29 October 2019).
- Yeung, K.H.T.; Duclos, P.; Nelson, E.A.S.; Hutubessy, R.C.W. An update of the global burden of pertussis in children younger than 5 years: A modelling study. Lancet Infect. Dis. 2017, 17, 974–980. [Google Scholar] [CrossRef]
- Argondizo-Correia, C.; Rodrigues, A.K.S.; de Brito, C.A. Neonatal immunity to Bordetella pertussis infection and current prevention strategies. J. Immunol. Res. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Kuchar, E.; Karlikowska-Skwarnik, M.; Han, S.; Nitsch-Osuch, A. Pertussis: History of the disease and current prevention failure. Adv. Exp. Med. Biol. 2016, 934, 77–82. [Google Scholar] [CrossRef]
- Leslie, P.H.; Gardner, A.D. The phases of Haemophilus pertussis. J. Hyg. (Lond) 1931, 31, 423–434. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Pertussis vaccines: WHO position paper—September 2015. Wkly. Epidemiol. Rec. 2015, 90, 433–458. [Google Scholar]
- Hewlett, E.L.; Edwards, K.M. Clinical practice. Pertussis—not just for kids. N. Engl. J. Med. 2005, 352, 1215–1222. [Google Scholar] [CrossRef]
- Kilgore, P.E.; Salim, A.M.; Zervos, M.J.; Schmitt, H.J. Pertussis: Microbiology, disease, treatment, and prevention. Clin. Microbiol. Rev. 2016, 29, 449–486. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Sato, H. Development of acellular pertussis vaccines. Biologicals 1999, 27, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.K.; Verghese, V.P.; Ortiz, E. Primary vaccination of Indian infants at 6, 10, 14 weeks of age with a diphtheria, tetanus, acellular pertussis, inactivated poliovirus, Haemophilus influenzae type B conjugate vaccine (Pentaxim). Int. J. Infect. Dis. 2008, 12, e174. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.A.; Liese, J.; Madhi, S.A.; Ortiz, E. A DTaP-IPV//PRP∼T vaccine (Pentaxim): A review of 16 years’ clinical experience. Expert Rev. Vaccines 2011, 10, 981–1005. [Google Scholar] [CrossRef] [PubMed]
- Tregnaghi, M.; Zambrano, B.; Santos-Lima, E. Antibody persistence after a primary series of a new DTaP-IPV-Hep B-PRP-T combined vaccine or separate DTaP-IPV//PRP-T and hepatitis B vaccines at 2, 4, and 6 months of age and the effect of a subsequent DTaP-IPV//PRP-T booster vaccination at 18 months of age in healthy Argentinean infants. Pediatr. Infect. Dis. J. 2012, 31, e24–e30. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, L.; Hallander, H.O.; Olin, P.; Reizenstein, E.; Storsaeter, J. A controlled trial of a two-component acellular, a five-component acellular, and a whole-cell pertussis vaccine. N. Engl. J. Med. 1996, 334, 349–355. [Google Scholar] [CrossRef]
- Greco, D.; Salmaso, S.; Mastrantonio, P.; Giuliano, M.; Tozzi, A.E.; Anemona, A.; Ciofi degli Atti, M.L.; Giammanco, A.; Panei, P.; Blackwelder, W.C.; et al. A controlled trial of two acellular vaccines and one whole-cell vaccine against pertussis. Progetto Pertosse Working Group. N. Engl. J. Med. 1996, 334, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, D.W.; Rohani, P. Perplexities of pertussis: Recent global epidemiological trends and their potential causes. Epidemiol. Infect. 2014, 142, 672–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenech de Cellès, M.; Magpantay, F.M.; King, A.A.; Rohani, P. The pertussis enigma: Reconciling epidemiology, immunology and evolution. Proc. Biol. Sci. 2016, 283, 20152309. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.L.; Tiwari, T.; Moro, P.; Messonnier, N.E.; Reingold, A.; Sawyer, M.; Clark, T.A. Prevention of pertussis, tetanus, and diphtheria with vaccines in the United States: Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm. Rep. 2018, 67, 1–44. [Google Scholar] [CrossRef]
- Preziosi, M.P.; Halloran, M.E. Effects of pertussis vaccination on transmission: Vaccine efficacy for infectiousness. Vaccine 2003, 21, 1853–1861. [Google Scholar] [CrossRef]
- Amirthalingam, G.; Campbell, H.; Ribeiro, S.; Fry, N.K.; Ramsay, M.; Miller, E.; Andrews, N. Sustained effectiveness of the maternal pertussis immunization program in England 3 years following introduction. Clin. Infect. Dis. 2016, 63, S236–S243. [Google Scholar] [CrossRef]
- Carbonetti, N.H. Pertussis toxin and adenylate cyclase toxin: Key virulence factors of Bordetella pertussis and cell biology tools. Future Microbiol. 2010, 5, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Higgs, R.; Higgins, S.C.; Ross, P.J.; Mills, K.H. Immunity to the respiratory pathogen Bordetella pertussis. Mucosal Immunol. 2012, 5, 485–500. [Google Scholar] [CrossRef] [Green Version]
- Freyberg, Z.; Harvill, E.T. Pathogen manipulation of host metabolism: A common strategy for immune evasion. PLoS Pathog. 2017, 13, e1006669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirimanjeswara, G.S.; Agosto, L.M.; Kennett, M.J.; Bjornstad, O.N.; Harvill, E.T. Pertussis toxin inhibits neutrophil recruitment to delay antibody-mediated clearance of Bordetella pertussis. J. Clin. Invest. 2005, 115, 3594–3601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shumilla, J.A.; Lacaille, V.; Hornell, T.M.; Huang, J.; Narasimhan, S.; Relman, D.A.; Mellins, E.D. Bordetella pertussis infection of primary human monocytes alters HLA-DR expression. Infect. Immun. 2004, 72, 1450–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, A.; Volpe, E.; Auricchio, G.; Colizzi, V.; Baldini, P.M. Influence of pertussis toxin on CD1a isoform expression in human dendritic cells. J. Clin. Immunol. 2006, 26, 153–159. [Google Scholar] [CrossRef]
- Esposito, S.; Stefanelli, P.; Fry, N.K.; Fedele, G.; He, Q.; Paterson, P.; Tan, T.; Knuf, M.; Rodrigo, C.; Weil Olivier, C.; et al. Pertussis prevention: Reasons for resurgence, and differences in the current acellular pertussis vaccines. Front. Immunol. 2019, 10, 1344. [Google Scholar] [CrossRef] [Green Version]
- Thierry-Carstensen, B.; Dalby, T.; Stevner, M.A.; Robbins, J.B.; Schneerson, R.; Trollfors, B. Experience with monocomponent acellular pertussis combination vaccines for infants, children, adolescents and adults—a review of safety, immunogenicity, efficacy and effectiveness studies and 15 years of field experience. Vaccine 2013, 31, 5178–5191. [Google Scholar] [CrossRef]
- Trollfors, B.; Taranger, J.; Lagergard, T.; Lind, L.; Sundh, V.; Zackrisson, G.; Lowe, C.U.; Blackwelder, W.; Robbins, J.B. A placebo-controlled trial of a pertussis-toxoid vaccine. N. Engl. J. Med. 1995, 333, 1045–1050. [Google Scholar] [CrossRef]
- Mattoo, S.; Cherry, J.D. Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin. Microbiol. Rev. 2005, 18, 326–382. [Google Scholar] [CrossRef] [Green Version]
- McGuirk, P.; Mills, K.H. Direct anti-inflammatory effect of a bacterial virulence factor: IL-10-dependent suppression of IL-12 production by filamentous hemagglutinin from Bordetella pertussis. Eur. J. Immunol. 2000, 30, 415–422. [Google Scholar] [CrossRef]
- Mills, K.H.; Ryan, M.; Ryan, E.; Mahon, B.P. A murine model in which protection correlates with pertussis vaccine efficacy in children reveals complementary roles for humoral and cell-mediated immunity in protection against Bordetella pertussis. Infect. Immun. 1998, 66, 594–602. [Google Scholar] [CrossRef] [Green Version]
- Ad Hoc Group for the Study of Pertussis Vaccines. Placebo-controlled trial of two acellular pertussis vaccines in Sweden--protective efficacy and adverse events. Lancet 1988, 1, 955–960. [Google Scholar]
- Marzouqi, I.; Richmond, P.; Fry, S.; Wetherall, J.; Mukkur, T. Development of improved vaccines against whooping cough: Current status. Hum. Vaccin. 2010, 6, 543–553. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.B.; Huang, Y.Y.; Halperin, S.A.; Anderson, R.; Morris, A.; Macmillan, A.; Jones, T.; Burt, D.S.; Van Nest, G.; Lee, S.F. Immunogenicity and protective efficacy of a recombinant filamentous haemagglutinin from Bordetella pertussis. Clin. Exp. Immunol. 2006, 144, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Storsaeter, J.; Olin, P. Relative efficacy of two acellular pertussis vaccines during three years of passive surveillance. Vaccine 1992, 10, 142–144. [Google Scholar] [CrossRef]
- Everest, P.; Li, J.; Douce, G.; Charles, I.; De Azavedo, J.; Chatfield, S.; Dougan, G.; Roberts, M. Role of the Bordetella pertussis P.69/pertactin protein and the P.69/pertactin RGD motif in the adherence to and invasion of mammalian cells. Microbiolgy 1996, 142, 3261–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.W.; Pawloski, L.; Williams, M.; Weening, K.; DeBolt, C.; Qin, X.; Reynolds, L.; Kenyon, C.; Giambrone, G.; Kudish, K.; et al. Pertactin-negative Bordetella pertussis strains: Evidence for a possible selective advantage. Clin. Infect. Dis. 2015, 60, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Inatsuka, C.S.; Xu, Q.; Vujkovic-Cvijin, I.; Wong, S.; Stibitz, S.; Miller, J.F.; Cotter, P.A. Pertactin is required for Bordetella species to resist neutrophil-mediated clearance. Infect. Immun. 2010, 78, 2901–2909. [Google Scholar] [CrossRef] [Green Version]
- Carbonetti, N.H.; Wirsing von Konig, C.H.; Lan, R.; Jacob-Dubuisson, F.; Cotter, P.A.; Deora, R.; Merkel, T.J.; van Els, C.A.; Locht, C.; Hozbor, D.; et al. Highlights of the 11th International Bordetella Symposium: From basic biology to vaccine development. Clin. Vaccine Immunol. 2016, 23, 842–850. [Google Scholar] [CrossRef] [Green Version]
- Olin, P.; Rasmussen, F.; Gustafsson, L.; Hallander, H.O.; Heijbel, H. Randomised controlled trial of two-component, three-component, and five-component acellular pertussis vaccines compared with whole-cell pertussis vaccine. Ad Hoc Group for the Study of Pertussis Vaccines. Lancet 1997, 350, 1569–1577. [Google Scholar] [CrossRef]
- Zhang, L.; Prietsch, S.O.; Axelsson, I.; Halperin, S.A. Acellular vaccines for preventing whooping cough in children. Cochrane Database Syst. Rev. 2014. [Google Scholar] [CrossRef]
- Jefferson, T.; Rudin, M.; DiPietrantonj, C. Systematic review of the effects of pertussis vaccines in children. Vaccine 2003, 21, 2003–2014. [Google Scholar] [CrossRef]
- Desauziers, E.; Hessel, L.; Decker, M.D.; Caro, J.J.; Liese, J.G. Systematic review of the effects of pertussis vaccines in children. Vaccine 2004, 22, 2681–2684. [Google Scholar] [CrossRef] [PubMed]
- Gorringe, A.R.; Vaughan, T.E. Bordetella pertussis fimbriae (Fim): Relevance for vaccines. Expert Rev. Vaccines 2014, 13, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Decker, K.B.; Boucher, P.E.; Hinton, D.; Stibitz, S. Novel architectural features of Bordetella pertussis fimbrial subunit promoters and their activation by the global virulence regulator BvgA. Mol. Microbiol. 2010, 77, 1326–1340. [Google Scholar] [CrossRef] [Green Version]
- Heikkinen, E.; Xing, D.K.; Olander, R.M.; Hytonen, J.; Viljanen, M.K.; Mertsola, J.; He, Q. Bordetella pertussis isolates in Finland: Serotype and fimbrial expression. BMC Microbiol. 2008, 8, 162. [Google Scholar] [CrossRef] [Green Version]
- Funnell, S.G.; Robinson, A. A novel adherence assay for Bordetella pertussis using tracheal organ cultures. FEMS Microbiol. Lett. 1993, 110, 197–203. [Google Scholar] [CrossRef]
- Rodriguez, M.E.; Hellwig, S.M.; Perez Vidakovics, M.L.; Berbers, G.A.; van de Winkel, J.G. Bordetella pertussis attachment to respiratory epithelial cells can be impaired by fimbriae-specific antibodies. FEMS Immunol. Med. Microbiol. 2006, 46, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Queenan, A.M.; Dowling, D.J.; Cheng, W.K.; Fae, K.; Fernandez, J.; Flynn, P.J.; Joshi, S.; Brightman, S.E.; Ramirez, J.; Serroyen, J.; et al. Increasing FIM2/3 antigen-content improves efficacy of Bordetella pertussis vaccines in mice in vivo without altering vaccine-induced human reactogenicity biomarkers in vitro. Vaccine 2019, 37, 80–89. [Google Scholar] [CrossRef]
- Hallander, H.; Advani, A.; Alexander, F.; Gustafsson, L.; Ljungman, M.; Pratt, C.; Hall, I.; Gorringe, A.R. Antibody responses to Bordetella pertussis Fim2 or Fim3 following immunization with a whole-cell, two-component, or five-component acellular pertussis vaccine and following pertussis disease in children in Sweden in 1997 and 2007. Clin. Vaccine Immunol. 2014, 21, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Storsaeter, J.; Hallander, H.O.; Gustafsson, L.; Olin, P. Levels of anti-pertussis antibodies related to protection after household exposure to Bordetella pertussis. Vaccine 1998, 16, 1907–1916. [Google Scholar] [CrossRef]
- Vesikari, T.; Xu, J.; Johnson, D.R.; Hall, J.; Marček, T.; Goveia, M.G.; Acosta, C.J.; Lee, A.W. Hepatitis B and pertussis antibodies in 4- to 5-year-old children previously vaccinated with different hexavalent vaccines. Hum. Vaccine Immunother. 2020, 16, 867–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, D.P.; Doemland, M.; Bettinger, J.A.; Scheifele, D.W.; Halperin, S.A.; IMPACT Investigators; Waters, V.; Kandola, K. Epidemiology of pertussis and Haemophilus influenzae type b disease in Canada with exclusive use of a diphtheria-tetanus-acellular pertussis-inactivated poliovirus-Haemophilus influenzae type b pediatric combination vaccine and an adolescent-adult tetanus-diphtheria-acellular pertussis vaccine: Implications for disease prevention in the United States. Pediatr. Infect. Dis. J. 2009, 28, 521–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, C.F.A.; Andrews, N.; Dabrera, G.; Ribeiro, S.; Stowe, J.; Ramsay, M.; Amirthalingam, G. National outbreak of pertussis in England, 2011–2012: A case-control study comparing 3-component and 5-component acellular vaccines with whole-cell pertussis vaccines. Clin. Infect. Dis. 2020, 70, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Hovingh, E.S.; Kuipers, B.; Bonacic Marinovic, A.A.; Jan Hamstra, H.; Hijdra, D.; Mughini Gras, L.; van Twillert, I.; Jongerius, I.; van Els, C.; Pinelli, E. Detection of opsonizing antibodies directed against a recently circulating Bordetella pertussis strain in paired plasma samples from symptomatic and recovered pertussis patients. Sci. Rep. 2018, 8, 12039. [Google Scholar] [CrossRef] [PubMed]
- Jongerius, I.; Schuijt, T.J.; Mooi, F.R.; Pinelli, E. Complement evasion by Bordetella pertussis: Implications for improving current vaccines. J. Mol. Med. 2015, 93, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berggard, K.; Johnsson, E.; Morfeldt, E.; Persson, J.; Stalhammar-Carlemalm, M.; Lindahl, G. Binding of human C4BP to the hypervariable region of M protein: A molecular mechanism of phagocytosis resistance in Streptococcus pyogenes. Mol. Microbiol. 2001, 42, 539–551. [Google Scholar] [CrossRef] [Green Version]
- Kohberger, R.C.; Jemiolo, D.; Noriega, F. Prediction of pertussis vaccine efficacy using a correlates of protection model. Vaccine 2008, 26, 3516–3521. [Google Scholar] [CrossRef]
- Taranger, J.; Trollfors, B.; Lagergard, T.; Sundh, V.; Bryla, D.A.; Schneerson, R.; Robbins, J.B. Correlation between pertussis toxin IgG antibodies in postvaccination sera and subsequent protection against pertussis. J. Infect. Dis. 2000, 181, 1010–1013. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.A. Complex correlates of protection after vaccination. Clin. Infect. Dis. 2013, 56, 1458–1465. [Google Scholar] [CrossRef] [PubMed]
- Warfel, J.M.; Zimmerman, L.I.; Merkel, T.J. Acellular pertussis vaccines protect against disease but fail to prevent infection and transmission in a nonhuman primate model. Proc. Natl. Acad. Sci. USA 2014, 111, 787–792. [Google Scholar] [CrossRef] [Green Version]
- Barkoff, A.M.; Mertsola, J.; Pierard, D.; Dalby, T.; Hoegh, S.V.; Guillot, S.; Stefanelli, P.; van Gent, M.; Berbers, G.; Vestrheim, D.; et al. Pertactin-deficient Bordetella pertussis isolates: Evidence of increased circulation in Europe, 1998 to 2015. Euro. Surveill. 2019, 24, 1700832. [Google Scholar] [CrossRef] [PubMed]
- Weigand, M.R.; Williams, M.M.; Peng, Y.; Kania, D.; Pawloski, L.C.; Tondella, M.L.; CDC Pertussis Working Group. Genomic survey of Bordetella pertussis diversity, United States, 2000–2013. Emerg. Infect. Dis. 2019, 25, 780–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breakwell, L.; Kelso, P.; Finley, C.; Schoenfeld, S.; Goode, B.; Misegades, L.K.; Martin, S.W.; Acosta, A.M. Pertussis vaccine effectiveness in the setting of pertactin-deficient pertussis. Pediatrics 2016, 137, e20153973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooi, F.R.; van Loo, I.H.; van Gent, M.; He, Q.; Bart, M.J.; Heuvelman, K.J.; de Greeff, S.C.; Diavatopoulos, D.; Teunis, P.; Nagelkerke, N.; et al. Bordetella pertussis strains with increased toxin production associated with pertussis resurgence. Emerg. Infect. Dis. 2009, 15, 1206–1213. [Google Scholar] [CrossRef]
- van Gent, M.; Heuvelman, C.J.; van der Heide, H.G.; Hallander, H.O.; Advani, A.; Guiso, N.; Wirsing von Konig, C.H.; Vestrheim, D.F.; Dalby, T.; Fry, N.K.; et al. Analysis of Bordetella pertussis clinical isolates circulating in European countries during the period 1998–2012. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 821–830. [Google Scholar] [CrossRef]
- de Gouw, D.; Hermans, P.W.; Bootsma, H.J.; Zomer, A.; Heuvelman, K.; Diavatopoulos, D.A.; Mooi, F.R. Differentially expressed genes in Bordetella pertussis strains belonging to a lineage which recently spread globally. PLoS ONE 2014, 9, e84523. [Google Scholar] [CrossRef]
- Misiak, A.; Leuzzi, R.; Allen, A.C.; Galletti, B.; Baudner, B.C.; D’Oro, U.; O’Hagan, D.T.; Pizza, M.; Seubert, A.; Mills, K.H.G. Addition of a TLR7 agonist to an acellular pertussis vaccine enhances Th1 and Th17 responses and protective immunity in a mouse model. Vaccine 2017, 35, 5256–5263. [Google Scholar] [CrossRef]
- Allen, A.C.; Wilk, M.M.; Misiak, A.; Borkner, L.; Murphy, D.; Mills, K.H.G. Sustained protective immunity against Bordetella pertussis nasal colonization by intranasal immunization with a vaccine-adjuvant combination that induces IL-17-secreting TRM cells. Mucosal Immunol. 2018, 11, 1763–1776. [Google Scholar] [CrossRef] [Green Version]
- Jennings-Gee, J.; Quataert, S.; Ganguly, T.; D’Agostino, R., Jr.; Deora, R.; Dubey, P. The adjuvant Bordetella colonization factor A attenuates alum-induced Th2 responses and enhances Bordetella pertussis clearance from mouse lungs. Infect. Immun. 2018, 86, e0093517. [Google Scholar] [CrossRef] [Green Version]
- Boehm, D.T.; Wolf, M.A.; Hall, J.M.; Wong, T.Y.; Sen-Kilic, E.; Basinger, H.D.; Dziadowicz, S.A.; Gutierrez, M.P.; Blackwood, C.B.; Bradford, S.D.; et al. Intranasal acellular pertussis vaccine provides mucosal immunity and protects mice from Bordetella pertussis. NPJ Vaccines 2019, 4, 40. [Google Scholar] [CrossRef]
- Locht, C.; Papin, J.F.; Lecher, S.; Debrie, A.S.; Thalen, M.; Solovay, K.; Rubin, K.; Mielcarek, N. Live attenuated pertussis vaccine BPZE1 protects baboons against Bordetella pertussis disease and infection. J. Infect. Dis. 2017, 216, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Thorstensson, R.; Trollfors, B.; Al-Tawil, N.; Jahnmatz, M.; Bergstrom, J.; Ljungman, M.; Torner, A.; Wehlin, L.; Van Broekhoven, A.; Bosman, F.; et al. A phase I clinical study of a live attenuated Bordetella pertussis vaccine--BPZE1; a single centre, double-blind, placebo-controlled, dose-escalating study of BPZE1 given intranasally to healthy adult male volunteers. PLoS ONE 2014, 9, e83449. [Google Scholar] [CrossRef]
- Cheung, G.Y.; Xing, D.; Prior, S.; Corbel, M.J.; Parton, R.; Coote, J.G. Effect of different forms of adenylate cyclase toxin of Bordetella pertussis on protection afforded by an acellular pertussis vaccine in a murine model. Infect. Immun. 2006, 74, 6797–6805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurita, M.E.; Wilk, M.M.; Carriquiriborde, F.; Bartel, E.; Moreno, G.; Misiak, A.; Mills, K.H.G.; Hozbor, D. A pertussis outer membrane vesicle-based vaccine induces lung-resident memory CD4 T cells and protection against Bordetella pertussis, including pertactin deficient strains. Front. Cell. Infect. Microbiol. 2019, 9, 125. [Google Scholar] [CrossRef]
- Gestal, M.C.; Howard, L.K.; Dewan, K.; Johnson, H.M.; Barbier, M.; Bryant, C.; Soumana, I.H.; Rivera, I.; Linz, B.; Blas-Machado, U.; et al. Enhancement of immune response against Bordetella spp. by disrupting immunomodulation. Sci. Rep. 2019, 9, 20261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherry, J.D.; Xing, D.X.; Newland, P.; Patel, K.; Heininger, U.; Corbel, M.J. Determination of serum antibody to Bordetella pertussis adenylate cyclase toxin in vaccinated and unvaccinated children and in children and adults with pertussis. Clin. Infect. Dis. 2004, 38, 502–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo, R.; Fernandez, S.; Zayas, C.; Acosta, A.; Sarmiento, M.E.; Ferro, V.A.; Rosenqvist, E.; Campa, C.; Cardoso, D.; Garcia, L.; et al. Bacterial outer membrane vesicles and vaccine applications. Front. Immunol. 2014, 5, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerritzen, M.J.H.; Martens, D.E.; Wijffels, R.H.; van der Pol, L.; Stork, M. Bioengineering bacterial outer membrane vesicles as vaccine platform. Biotechnol. Adv. 2017, 35, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Cecil, J.D.; Sirisaengtaksin, N.; O’Brien-Simpson, N.M.; Krachler, A.M. Outer membrane vesicle-host cell interactions. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Kanzawa, Y.; Harada, T.; Koreeda, A.; Harada, A. Curdlan gel formed by neutralizing its alkaline solution. Agric. Biol. Chem. 1987, 51, 1839–1843. [Google Scholar]
- Debrie, A.S.; Coutte, L.; Raze, D.; Mooi, F.; Alexander, F.; Gorringe, A.; Mielcarek, N.; Locht, C. Construction and evaluation of Bordetella pertussis live attenuated vaccine strain BPZE1 producing Fim3. Vaccine 2018, 36, 1345–1352. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| 2-Component aP a | 5-Component aP | wP | ||||
|---|---|---|---|---|---|---|
| VE, % | 95% CI | VE, % | 95% CI | VE, % | 95% CI | |
| Culture-confirmed pertussis with ≥21 days of paroxysmal cough | ||||||
| Storsaeter 1998 [52] | 42.4 | 19.9, 58.5 | 75.4 | 59.2, 85.2 | 28.5 | 1.6, 48.0 |
| Gustafsson 1996 [14] | 58.9 | 50.9, 65.9 | 85.2 | 80.6, 88.8 | 48.3 | 37.0, 57.6 |
| ≥1 day of paroxysmal cough and positive laboratory criteria | ||||||
| Storsaeter 1998 [52] | 5.7 | −9.1, 19.6 | 61.8 | 47.4, 72.2 | 3.1 | −12.9, 16.8 |
| Novel Adjuvants | |||
| Studied In | Adjuvant | Effect | Immune Response |
| Mice [69] | Alum-absorbed TLR7a agonist | Higher PT neutralizing antibodies and increased inhibition of FHA binding to lung epithelium | Induced TH1/TH17 response and IgG2a/b |
| Mice [70] | Cyclic dimeric guanosine monophosphate with a B. pertussis TLR2 agonist | Combined effect of intracellular induction of interferon genes and broader TLR stimulation | Induced IFN-β, IL-12 and IL-23 and maturation of dendritic cells |
| Mice [71] | B. bronchiseptica colonization factor added to alum | Improved clearance of B. pertussis | Induced TH1/TH17 response and IL-17 |
| Mice [72] | Curdlan (1,3-β-glucan) | “Sticky” properties promote vaccine localization; binds to dendritic cells and induces NF-κB | Increased IL-17; intranasal mucosal IgA and serum IgG response |
| Novel Vaccines | |||
| Studied In | Target/Strategy | Effect | Immune Response |
| Non-human primates; humans [73,74] | Live attenuated intranasal B. pertussis BPZE1 a | Reduced nasopharyngeal colonization | Induced serum IgA and systemic IgG response |
| Mice [75] | Detoxified ACT combined with 3-component aP vaccine | Reduced bacterial counts in lungs postchallenge | Induced IgG2a response and stronger TH1 and TH2 response |
| Mice [76] | B. pertussis OMV | Induced lung tissue-resident memory cells and reduced bacterial counts in lungs | Increased IL-17 levels |
| Mice [77] | btrS-deficient strain of B. bronchiseptica | Induced broad cross-species protection against B. pertussis, B. parapertussis and B. bronchiseptica | Increased immunity by disrupting bacterial suppression of host immune responses |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dewan, K.K.; Linz, B.; DeRocco, S.E.; Harvill, E.T. Acellular Pertussis Vaccine Components: Today and Tomorrow. Vaccines 2020, 8, 217. https://doi.org/10.3390/vaccines8020217
Dewan KK, Linz B, DeRocco SE, Harvill ET. Acellular Pertussis Vaccine Components: Today and Tomorrow. Vaccines. 2020; 8(2):217. https://doi.org/10.3390/vaccines8020217
Chicago/Turabian StyleDewan, Kalyan K., Bodo Linz, Susan E. DeRocco, and Eric T. Harvill. 2020. "Acellular Pertussis Vaccine Components: Today and Tomorrow" Vaccines 8, no. 2: 217. https://doi.org/10.3390/vaccines8020217
APA StyleDewan, K. K., Linz, B., DeRocco, S. E., & Harvill, E. T. (2020). Acellular Pertussis Vaccine Components: Today and Tomorrow. Vaccines, 8(2), 217. https://doi.org/10.3390/vaccines8020217