Sex-Differential Impact of Human Cytomegalovirus Infection on In Vitro Reactivity to Toll-Like Receptor 2, 4 and 7/8 Stimulation in Gambian Infants

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Ethics Statement
2.3. Diagnosis of HCMV Infection
2.4. Whole Blood Cultures and Cytokine Multiplex Analysis
2.5. Statistical Analysis
3. Results
3.1. Infant Characteristics
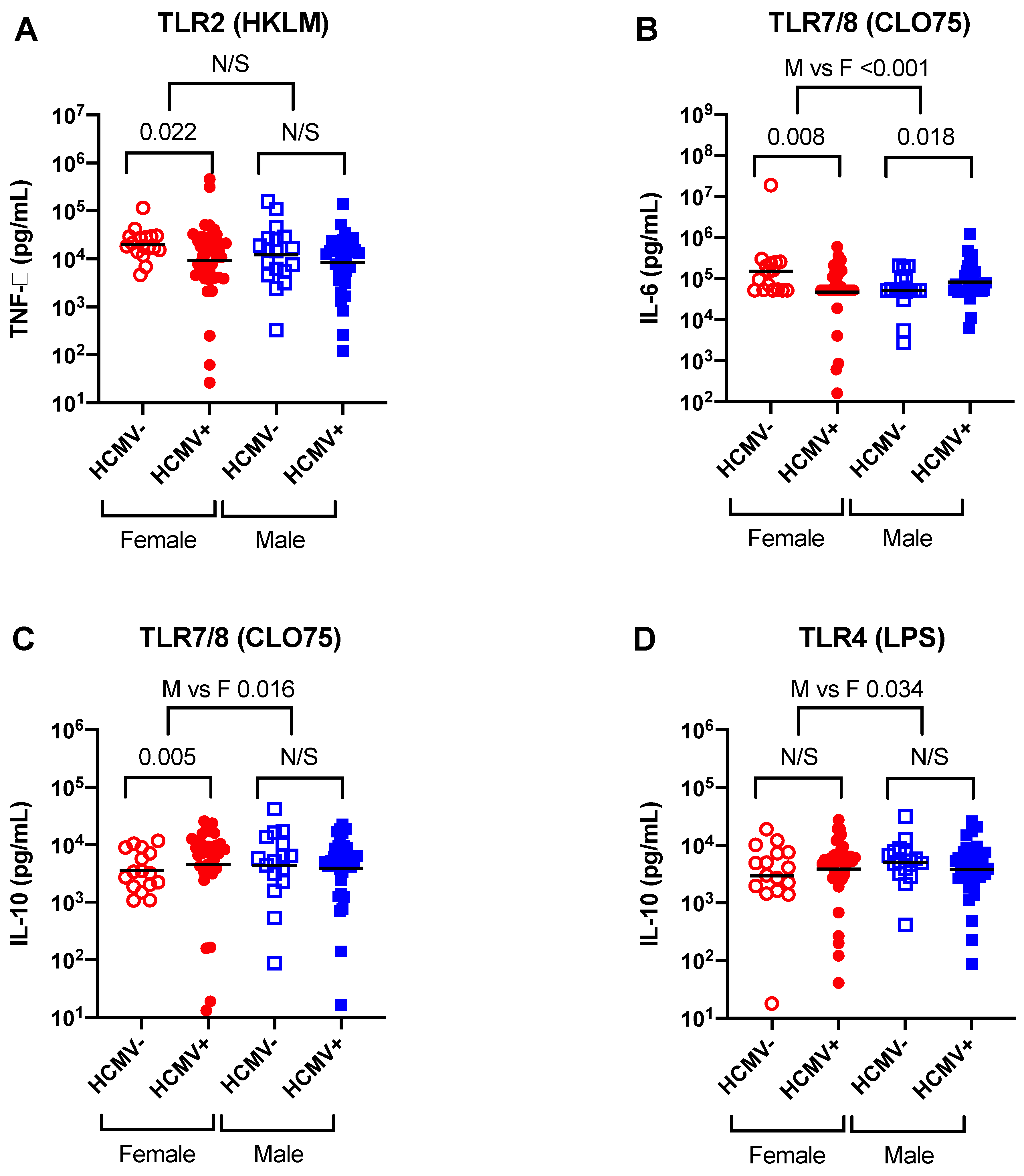
3.2. Minimal Effects of HCMV Infection on TLR Ligand Responses When Males and Females Are Analysed Together
3.3. Evidence for Effects of HCMV Infection on TLR Ligand Responses in Females but Not Males
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miles, D.J.; van der Sande, M.; Jeffries, D.; Kaye, S.; Ismaili, J.; Ojuola, O.; Sanneh, M.; Touray, E.S.; Waight, P.; Rowland-Jones, S.; et al. Cytomegalovirus infection in gambian infants leads to profound CD8 T-Cell differentiation. J. Virol. 2007, 81, 5766–5776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, E.; Zhang, Z.; Thorp, E.B.; Hummel, M. Cytomegalovirus latency and reactivation: An intricate interplay with the host immune response. Front. Cell Infect. Microbiol. 2020, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Cheeran, M.C.; James, R.; Lokensgard, J.R.; Schleiss, M.R. Neuropathogenesis of congenital cytomegalovirus infection: Disease mechanisms and prospects for intervention. Clin. Microbiol. Rev. 2009, 22, 99–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Berg, P.J.; Heutinck, K.M.; Raabe, R.; Minnee, R.C.; La Young, S.; van Donselaar-van der Pant, K.A.; Bemelman, F.J.; van Lier, R.A.; ten Berge, I.J. Human cytomegalovirus induces systemic immune activation characterized by a Type 1 Cytokine Signature. J. Infect. Dis. 2010, 202, 690–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compton, T.; Kurt-Jones, E.A.; Boehme, K.W.; Belko, J.; Latz, E.; Golenbock, D.T.; Finberg, R.W. Human cytomegalovirus activates inflammatory cytokine responses via CD14 and toll-like receptor 2. J. Virol. 2003, 77, 4588–4596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juckem, L.K.; Boehme, K.W.; Feire, A.L.; Compton, T. Differential initiation of innate immune responses induced by human cytomegalovirus entry into fibroblast cells. J. Immunol. 2008, 180, 4965–4977. [Google Scholar] [CrossRef] [Green Version]
- Loewendorf, A.; Benedict, C.A. Modulation of host innate and adaptive immune defenses by Cytomegalovirus: Timing is everything. J. Intern. Med. 2010, 267, 483–501. [Google Scholar] [CrossRef]
- Jun Cui, J.; Chen, Y.; Wang, H.Y.; Wang, R.F. Mechanisms and pathways of innate immune activation and regulation in health and cancer. Hum. Vaccin. Immunother. 2014, 10, 3270–3285. [Google Scholar] [CrossRef] [Green Version]
- Terrazzini, N.; Kern, F. Cell-mediated immunity to human CMV infection: A brief overview. F1000Prime Rep. 2014, 6, 28. [Google Scholar] [CrossRef]
- Camille Khairallah, C.; Julie Déchanet-Merville, J.; Capone, M. γδ T Cell-mediated immunity to cytomegalovirus infection. Front. Immunol. 2017, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Seckert, C.K.; Griessl, M.; Büttner, J.K.; Scheller, S.; Simon, C.O.; Kropp, K.A.; Renzaho, A.; Kühnapfel, B.; Grzimek, N.K.; Reddehase, M.J. Viral latency drives ‘Memory inflation’: A unifying hypothesis linking two hallmarks of Cytomegalovirus infection. Med. Microbiol. Immunol. 2012, 201, 551–566. [Google Scholar] [CrossRef] [PubMed]
- Slobedman, B.; Mocarski, E.S. Mechanisms modulating immune clearance during human cytomegalovirus latency. Proc. Natl. Acad. Sci. USA 2012, 109, 14291–14292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddehase, M.J. ‘Checks and balances’ in cytomegalovirus-host cohabitation. Med. Microbiol. Immunol. 2019, 208, 259–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller-Kittrell, M.; Sparer, T.E. Feeling manipulated: Cytomegalovirus immune manipulation. Virol. J. 2009, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Paulus, C.; Krauss, S.; Nevels, M. A human cytomegalovirus antagonist of type I IFN-dependent signal transducer and activator of transcription signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 3840–3845. [Google Scholar] [CrossRef] [Green Version]
- Goodier, M.R.; Jonjić, S.; Eleanor, M.; Riley, E.M.; Juranić Lisnić, V. CMV and Natural killer cells: Shaping the response to vaccination. Eur. J. Immunol. 2018, 48, 50–65. [Google Scholar] [CrossRef]
- Christina Paulus, C.; Nevels, M. The human cytomegalovirus major immediate-early proteins as antagonists of intrinsic and innate antiviral host responses. Viruses 2009, 1, 760–779. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.K.; Gottlieb, D.J.; Plachter, B.; Pepperl-Klindworth, S.; Avdic, S.; Cunningham, A.L.; Abendroth, A.; Slobedman, B. The role of the human Cytomegalovirus UL111A gene in Down-Regulating CD4+ T-cell recognition of latently infected cells: Implications for virus elimination during latency. Blood 2009, 114, 4128–4137. [Google Scholar] [CrossRef] [Green Version]
- Roark, H.K.; Jenks, J.A.; Permar, S.R.; Schleiss, M.R. Animal models of congenital Cytomegalovirus transmission: Implications for vaccine development. J. Infect. Dis. 2020, 221, S60–S73. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Joosten, L.A.; Latz, E.; Mills, K.H.; Natoli, G.; Stunnenberg, H.G.; O‘Neill, L.A.; Xavier, R.J. Trained immunity: A program of innate immune memory in health and disease. Science 2016, 352. [Google Scholar] [CrossRef] [Green Version]
- Luetke-Eversloh, M.; Quirin Hammer, Q.; Pawel Durek, P.; Karl Nordström, K.; Gasparoni, G.; Pink, M.; Hamann, A.; Walter, J.; Chang, H.D.; Dong, J.; et al. Human Cytomegalovirus drives epigenetic imprinting of the IFNG locus in NKG2Chi natural killer cells. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Q.; Romagnani, C. About training and memory: NK-Cell adaptation to viral infections. Adv. Immunol. 2017, 133, 171–207. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.J.; Schmid, S.D.; Hyde, T.B. Review of Cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, S.; Derhovanessian, E.; Steinhagen-Thiessen, E.; Goldeck, D.; Müller, L.; Pawelec, G. Impact of age, sex and CMV-infection on peripheral T cell phenotypes: Results from the Berlin BASE-II Study. Biogerontology 2015, 16, 631–643. [Google Scholar] [CrossRef]
- Villacres, M.C.; Longmate, J.; Auge, C.; Diamond, D.J. Predominant Type 1 CMV-specific memory T-helper response in humans: Evidence for gender differences in cytokine secretion. Hum. Immunol. 2004, 65, 476–485. [Google Scholar] [CrossRef]
- Reed, R.G.; Al-Attar, A.; Presnell, S.R.; Lutz, C.T.; Suzanne, C.; Segerstrom, S.C. A longitudinal study of the stability, variability, and interdependencies among late-differentiated T and NK cell subsets in older adults. Exp. Gerontol. 2019, 121, 46–54. [Google Scholar] [CrossRef]
- Noho-Konteh, F.; Adetifa, J.U.; Cox, M.; Hossin, S.; Reynolds, J.; Le, M.T.; Sanyang, L.C.; Drammeh, A.; Plebanski, M.; Forster, T.; et al. Sex-differential non-vaccine-specific immunological effects of diphtheria-tetanus-pertussis and measles vaccination. Clin. Infect. Dis. 2016, 63, 1213–1226. [Google Scholar] [CrossRef] [Green Version]
- Dowling, D.J.; Tan, Z.; Prokopowicz, Z.M.; Palmer, C.D.; Maura-Ann, H.; Matthews, M.A.H.; Dietsch, G.N.; Hershberg, R.M.; Levy, O. The ultra-potent and selective TLR8 agonist VTX-294 activates human newborn and adult leukocytes. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Ndure, J.; Noho-Konteh, F.; Adetifa, J.U.; Cox, M.; Barker, F.; Le, M.T.; Sanyang, L.C.; Drammeh, A.; Whittle, H.C.; Clarke, E.; et al. Negative correlation between circulating CD4(+)FOXP3(+)CD127(-) regulatory T cells and subsequent antibody responses to infant measles vaccine but not diphtheria-tetanus-pertussis vaccine implies a regulatory role. Front. Immunol. 2017, 8, 921. [Google Scholar] [CrossRef] [Green Version]
- Preiser, W.; Brauninger, S.; Schwerdtfeger, R.; Ayliffe, U.; Garson, J.A.; Brink, N.S.; Franck, S.; Doerr, H.W.; Rabenau, H.F. Evaluation of diagnostic methods for the detection of cytomegalovirus in recipients of allogeneic stem cell transplants. J. Clin. Virol. 2001, 20, 59–70. [Google Scholar] [CrossRef]
- Marchant, A.; Appay, V.; van der Sande, M.; Dulphy, N.; Liesnard, C.; Kidd, M.; Kaye, S.; Ojuola, O.; Gillespie, G.M.; Vargas Cuero, A.L.; et al. Mature CD8(+) T lymphocyte response to viral infection during fetal life. J. Clin. Investig. 2003, 111, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Burl, S.; Townend, J.; Njie-Jobe, J.; Cox, M.; Adetifa, U.J.; Touray, E.; Philbin, V.J.; Mancuso, C.; Kampmann, B.; Whittle, H.; et al. Age-dependent maturation of Toll-like receptor-mediated cytokine responses in Gambian infants. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flanagan, K.L.; Fink, A.L.; Plebanski, M.; Klein, S.L. Sex and gender differences in the outcomes of vaccination over the life course. Ann. Rev. Cell. Dev. Biol. 2017, 33, 577–599. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.; Flanagan, K.L. Effect of sex on vaccination outcomes: Important but frequently overlooked. Curr. Opin. Pharmacol. 2018, 41, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Feghali, C.A.; Wright, T.M. Cytokines in acute and chronic inflammation. Front. Biosci. 1997, 2, d12–d26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O‘Garra, A. Interleukin-10 and the interleukin-10 Receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like receptors and the control of immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Cox, M.; Adetifa, J.U.; Noho-Konteh, F.; Njie-Jobe, J.; Sanyang, L.C.; Drammeh, A.; Plebanski, M.; Whittle, H.C.; Rowland-Jones, S.L.; Robertson, I.; et al. Limited impact of human cytomegalovirus infection in African infants on vaccine-specific responses following diphtheria-tetanus-pertussis and measles vaccination. Front. Immunol. 2020, 11, 1083. [Google Scholar] [CrossRef] [PubMed]
- Bello, C.; Whittle, H. Cytomegalovirus infection in Gambian mothers and their babies. J. Clin. Pathol. 1991, 44, 366–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reikie, B.A.; Adams, R.C.; Ruck, C.E.; Ho, K.; Leligdowicz, A.; Pillay, S.; Naidoo, S.; Fortuno, E.S., 3rd; de Beer, C.; Preiser, W.; et al. Ontogeny of toll-like receptor mediated cytokine responses of South African infants throughout the first year of life. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, D.J.; Scott, E.A.; Scheid, A.; Bergelson, I.; Joshi, S.; Pietrasanta, C.; Brightman, S.; Sanchez-Schmitz, G.; van Haren, S.D.; Ninković, J.; et al. Toll-like receptor 8 agonist nanoparticles mimic immunomodulating effects of the live BCG vaccine and enhance neonatal innate and adaptive immune responses. J. Allergy Clin. Immunol. 2017, 140, 1339–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, D.J.; van Haren, S.D.; Scheid, A.; Bergelson, I.; Kim, D.; Mancuso, C.J.; Foppen, W.; Ozonoff, A.; Fresh, L.; Theriot, T.B. TLR7/8 adjuvant overcomes newborn hyporesponsiveness to pneumococcal conjugate vaccine at birth. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Group | HCMV+ | HCMV- | Total |
|---|---|---|---|
| All Infants | 73 (67.6%) | 35 (32.4%) | 108 |
| Males | 35 (66.0%) | 18 (34.0%) | 53 |
| Females | 38 (69.1%) | 17 (30.9%) | 55 |
| TNF-α | HCMV− | HCMV+ | Comparison (Unadjusted) | Comparison (Adjusted) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Geo Mean (GSD) | N | Geo Mean (GSD) | Δ | 95% CI | p-Value | Δ | 95% CI | p-Value | |
| Unstimulated | ||||||||||
| ALL | 34 | 73 (9.0) | 73 | 97 (9.5) | 25 | (−33 to 165) | 0.52 | 426 | (−88 to 1144) | 0.12 |
| Male | 17 | 139 (9.7) | 35 | 111 (9.5) | −27 | (−109 to 278) | 0.75 | 588 | (−56 to 1581) | 0.08 |
| Female | 17 | 38 (7.2) | 38 | 89 (9.8) | 51 | (−10 to 238) | 0.15 | 272 | (−1874 to 1503) | 0.54 |
| M vs. F | −78 | (−170 to 446) | 0.23 | 316 | (−585 to 2092) | 0.51 | ||||
| TLR2 (HKLM) | ||||||||||
| ALL | 35 | 15,529 (3.2) | 73 | 8818 (5.3) | −6711 | (−10,386 to −409) | 0.040 | −1369 | (−2414 to 107) | 0.066 |
| Male | 18 | 12,050 (4.2) | 35 | 7843 (4.4) | −4207 | (−8630 to 5937) | 0.31 | −338 | (−1712 to 1966) | 0.72 |
| Female | 17 | 20,311 (2.0) | 38 | 9614 (6.1) | −10,698 | (−15,214 to −2178) | 0.021 | −2738 | (−7615 to −497) | 0.022 |
| M vs. F | 6490 | (−1406 to 20,944) | 0.55 | 2400 | (−679 to 5880) | 0.21 | ||||
| TLR4 (LPS) | ||||||||||
| ALL | 32 | 6349 (5.0) | 72 | 4661 (5.4) | −1688 | (−3978 to 2816) | 0.37 | −181 | (−1108 to 1254) | 0.76 |
| Male | 16 | 6115 (4.2) | 34 | 5027 (4.5) | −1088 | (−4019 to 5941) | 0.66 | 27 | (−1150 to 2222) | 0.97 |
| Female | 16 | 6591 (6.2) | 38 | 4429 (6.0) | −2161 | (−4996 to 5709) | 0.45 | −396 | (−3837 to 1839) | 0.66 |
| M vs. F | 1073 | (−2644 to 15,336) | 0.77 | 422 | (−1047 to 3909) | 0.74 | ||||
| TLR5 (Flagellin) | ||||||||||
| ALL | 33 | 5245 (4.3) | 69 | 4405 (6.1) | −840 | (−2948 to 3202) | 0.60 | −239 | (−945 to 875) | 0.62 |
| Male | 17 | 5137 (4.6) | 34 | 4326 (5.4) | −811 | (−3429 to 5824) | 0.72 | 151 | (−749 to 1848) | 0.80 |
| Female | 16 | 5362 (4.2) | 35 | 4462 (6.8) | −900 | (−3553 to 5645) | 0.69 | −690 | (−3257 to 895) | 0.32 |
| M vs. F | 89 | (−3053 to 11,572) | 0.99 | 842 | (−247 to 3360) | 0.37 | ||||
| TLR7/8 (CLO75) | ||||||||||
| ALL | 32 | 3960 (3.5) | 68 | 3523 (6.7) | −438 | (−2072 to 2610) | 0.71 | −130 | (−798 to 830) | 0.76 |
| Male | 16 | 3420 (4.6) | 34 | 2678 (4.8) | −742 | (−2358 to 3331) | 0.60 | −164 | (−916 to 1123) | 0.75 |
| Female | 16 | 4586 (2.5) | 34 | 4319 (8.3) | −267 | (−2628 to 4943) | 0.88 | −74 | (−2341 to 1649) | 0.91 |
| M vs. F | −475 | (−2360 to 5890) | 0.77 | −91 | (−1054 to 1978) | 0.88 | ||||
| IL-1β | HCMV− | HCMV+ | Comparison (Unadjusted) | Comparison (Adjusted) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Geo Mean (GSD) | N | Geo Mean (GSD) | Δ | 95%CI | p-Value | Δ | 95%CI | p-Value | |
| Unstimulated | ||||||||||
| ALL | 34 | 83 (13.2) | 73 | 73 (11.8) | −10 | (−57 to 120) | 0.81 | −1223 | (−1939 to −212) | 0.022 |
| Male | 17 | 212 (14.4) | 35 | 86 (14.4) | −126 | (−194 to 194) | 0.25 | −1460 | (−2463 to 329) | 0.094 |
| Female | 17 | 32 (8.7) | 38 | 64 (10.4) | 32 | (−13 to 187) | 0.27 | −971 | (−4292 to 415) | 0.15 |
| M vs. F | −159 | (−232 to 376) | 0.11 | −489 | (−1693 to 2056) | 0.65 | ||||
| TLR2 (HKLM) | ||||||||||
| ALL | 35 | 14,290 (3.4) | 73 | 10,137 (7.0) | −4154 | (−8757 to 4280) | 0.27 | 97 | (−1256 to 2168) | 0.91 |
| Male | 18 | 13,556 (3.7) | 35 | 8436 (4.9) | −5120 | (−9800 to 5390) | 0.25 | −379 | (−1729 to 1869) | 0.68 |
| Female | 17 | 15,111 (3.3) | 38 | 11,646 (8.9) | −3464 | (−10,213 to 12,584) | 0.56 | 630 | (−5053 to 4807) | 0.65 |
| M vs. F | −1655 | (−7513 to 17,510) | 0.72 | −1009 | (−2943 to 3511) | 0.54 | ||||
| TLR4 (LPS) | ||||||||||
| ALL | 32 | 5668 (5.7) | 72 | 4038 (6.7) | −1630 | (−3744 to 2809) | 0.37 | −275 | (−1063 to 1021) | 0.61 |
| Male | 16 | 5280 (3.2) | 34 | 4519 (4.9) | −761 | (−3252 to 4789) | 0.70 | 137 | (−830 to 1985) | 0.83 |
| Female | 16 | 6084 (9.0) | 38 | 3737 (8.3) | −2348 | (−4987 to 6637) | 0.44 | −717 | (−4625 to 1314) | 0.39 |
| M vs. F | 1587 | (−1887 to 16,599) | 0.66 | 854 | (−370 to 3944) | 0.43 | ||||
| TLR5 (Flagellin) | ||||||||||
| ALL | 33 | 5063 (4.1) | 69 | 4024 (4.4) | −1039 | (−2832 to 2194) | 0.45 | −491 | (−1146 to 468) | 0.27 |
| Male | 17 | 6032 (4.0) | 34 | 3994 (3.4) | −2037 | (−4206 to 2706) | 0.30 | −897 | (−1665 to 345) | 0.13 |
| Female | 16 | 4204 (4.2) | 35 | 4046 (5.2) | −158 | (−2487 to 5332) | 0.93 | −132 | (−2266 to 1409) | 0.83 |
| M vs. F | −1880 | (−4623 to 6879) | 0.53 | −765 | (−1768 to 1240) | 0.38 | ||||
| TLR7/8 (CLO75) | ||||||||||
| ALL | 32 | 3940 (4.2) | 68 | 3014 (6.5) | −926 | (−2389 to 1917) | 0.43 | −348 | (−812 to 307) | 0.26 |
| Male | 16 | 3536 (6.0) | 34 | 2898 (4.0) | −638 | (−2456 to 4246) | 0.69 | −228 | (−904 to 884) | 0.63 |
| Female | 16 | 4390 (2.8) | 34 | 3105 (8.9) | −1284 | (−3053 to 2823) | 0.42 | −468 | (−2054 to 481) | 0.28 |
| M vs. F | 647 | (−1460 to 8365) | 0.82 | 240 | (−638 to 2029) | 0.69 | ||||
| IL-6 | HCMV− | HCMV+ | Comparison (Unadjusted) | Comparison (Adjusted) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Geo Mean (GSD) | N | Geo Mean (GSD) | Δ | 95%CI | p-Value | Δ | 95%CI | p-Value | |
| Unstimulated | ||||||||||
| ALL | 34 | 2188 (10.0) | 73 | 2480 (7.2) | 293 | (−1167 to 3839) | 0.78 | 6906 | (−2431 to 21,524) | 0.17 |
| Male | 17 | 5791 (8.5) | 35 | 3312 (7.3) | −2480 | (−4810 to 5388) | 0.37 | 2717 | (−11,543 to 30,714) | 0.78 |
| Female | 17 | 826 (8.0) | 38 | 2027 (7.0) | 1201 | (−166 to 5401) | 0.12 | 8854 | (−11,251 to 26,477) | 0.091 |
| M vs. F | −3681 | (−6360 to 10,339) | 0.085 | −6137 | (−23,124 to 34,746) | 0.37 | ||||
| TLR2 (HKLM) | ||||||||||
| ALL | 35 | 99,711 (2.6) | 73 | 73,393 (3.2) | −26,317 | (−51,057 to 11,003) | 0.14 | −8868 | (−26,981 to 17,855) | 0.46 |
| Male | 18 | 93,368 (2.9) | 35 | 88,146 (3.1) | −5221 | (−45,992 to 70,635) | 0.86 | 6634 | (−22,721 to 58,030) | 0.72 |
| Female | 17 | 10,6898 (2.3) | 38 | 64,113 (3.3) | −42,785 | (−69,160 to 2024) | 0.06 | −23,230 | (−96,836 to 10,025) | 0.14 |
| M vs. F | 37,564 | (−11,617 to 14,8820) | 0.28 | 29,863 | (−6245 to 10,6282) | 0.20 | ||||
| TLR4 (LPS) | ||||||||||
| ALL | 32 | 98,179 (4.4) | 72 | 61,846 (2.9) | −36,334 | (−62,872 to 10,152) | 0.11 | −17,692 | (−35,345 to 10,443) | 0.18 |
| Male | 16 | 11,4889 (2.7) | 34 | 75,326 (2.4) | −39,563 | (−72,343 to 18,475) | 0.15 | −18,329 | (−42,013 to 23,017) | 0.32 |
| Female | 16 | 83,899 (6.3) | 38 | 54,144 (3.2) | −29,755 | (−62,884 to 55,601) | 0.36 | −17,274 | (−10,0509 to 25,908) | 0.34 |
| M vs. F | −9808 | (−60,196 to 14,2396) | 0.98 | −1055 | (−33,561 to 77,520) | 0.89 | ||||
| TLR5 (Flagellin) | ||||||||||
| ALL | 33 | 71,286 (3.6) | 69 | 69,451 (2.8) | −1836 | (−28,974 to 42,709) | 0.92 | 861 | (−16,017 to 27,511) | 0.94 |
| Male | 17 | 77,027 (3.1) | 34 | 72,256 (2.7) | −4771 | (−38,656 to 59,038) | 0.84 | −73 | (−20,553 to 35,652) | 1.00 |
| Female | 16 | 65,655 (4.3) | 35 | 67,485 (2.9) | 1830 | (−34,427 to 80,182) | 0.94 | 1670 | (−58,715 to 47,904) | 0.92 |
| M vs. F | −6602 | (−52,201 to 11,7002) | 0.86 | −1743 | (−30,217 to 68,278) | 0.93 | ||||
| TLR7/8 (CLO75) | ||||||||||
| ALL | 32 | 87,048 (4.1) | 68 | 61,996 (4.3) | −25,052 | (−52,802 to 25,183) | 0.26 | −8595 | (−26,326 to 20,587) | 0.49 |
| Male | 16 | 50,778 (3.2) | 34 | 82,605 (2.8) | 31,827 | (−8476 to 11,0530) | 0.15 | 28,847 | (3689 to 72,477) | 0.018 |
| Female | 16 | 14,9225 (4.3) | 34 | 50,082 (5.3) | −99,143 | (−12,8262 to −29,577) | 0.014 | −57,211 | (−15,7017 to −20,296) | 0.008 |
| M vs. F | 13,0971 | (75,908 to 29,6114) | 0.005 | 86,058 | (50,886 to 17,2231) | <0.001 | ||||
| IL-12 | HCMV− | HCMV+ | Comparison (Unadjusted) | Comparison (Adjusted) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Geo Mean (GSD) | N | Geo Mean (GSD) | Δ | 95% CI | p-Value | Δ | 95% CI | p-Value | |
| Unstimulated | ||||||||||
| ALL | 34 | 25 (3.9) | 73 | 25 (6.1) | 0 | (−11 to 22) | 0.95 | −19 | (−60 to 50) | 0.52 |
| Male | 17 | 40 (3.1) | 35 | 27 (6.5) | −13 | (−28 to 22) | 0.35 | −51 | (−101 to 48) | 0.24 |
| Female | 17 | 16 (4.3) | 38 | 25 (6.0) | 9 | (−5 to 42) | 0.30 | 5 | (−166 to 113) | 0.89 |
| M vs. F | −22 | (−41 to 40) | 0.17 | −56 | (−120 to 114) | 0.36 | ||||
| TLR2 (HKLM) | ||||||||||
| ALL | 35 | 117 (2.1) | 73 | 116 (2.7) | −1 | (−35 to 45) | 0.95 | 7 | (−9 to 28) | 0.41 |
| Male | 18 | 121 (2.2) | 35 | 115 (2.1) | −6 | (−47 to 56) | 0.82 | 11 | (−14 to 45) | 0.44 |
| Female | 17 | 113 (2.1) | 38 | 117 (3.2) | 3 | (−42 to 78) | 0.91 | 4 | (−71 to 35) | 0.74 |
| M vs. F | −9 | (−65 to 98) | 0.81 | 6 | (−26 to 59) | 0.75 | ||||
| TLR4 (LPS) | ||||||||||
| ALL | 32 | 105 (2.2) | 72 | 108 (3.0) | 3 | (−31 to 51) | 0.90 | 5 | (−15 to 32) | 0.66 |
| Male | 16 | 100 (2.2) | 34 | 115 (2.5) | 14 | (−30 to 87) | 0.59 | 17 | (−9 to 54) | 0.22 |
| Female | 16 | 110 (2.3) | 38 | 103 (3.3) | −7 | (−50 to 66) | 0.81 | −7 | (−77 to 34) | 0.70 |
| M vs. F | 21 | (−38 to 144) | 0.59 | 24 | (−12 to 86) | 0.27 | ||||
| TLR5 (Flagellin) | ||||||||||
| ALL | 33 | 92 (2.5) | 69 | 112 (2.6) | 19 | (−17 to 72) | 0.34 | 7 | (−9 to 28) | 0.44 |
| Male | 17 | 105 (2.5) | 34 | 110 (3.2) | 5 | (−45 to 95) | 0.88 | 2 | (−21 to 38) | 0.88 |
| Female | 16 | 81 (2.5) | 35 | 113 (2.3) | 32 | (−13 to 107) | 0.20 | 11 | (−25 to 37) | 0.28 |
| M vs. F | −27 | (−87 to 104) | 0.47 | −9 | (−36 to 38) | 0.58 | ||||
| TLR7/8 (CLO75) | ||||||||||
| ALL | 32 | 95 (3.4) | 68 | 123 (3.5) | 28 | (−21 to 111) | 0.32 | 13 | (−11 to 47) | 0.32 |
| Male | 16 | 100 (3.4) | 34 | 112 (2.6) | 12 | (−44 to 120) | 0.75 | 28 | (−6 to 82) | 0.12 |
| Female | 16 | 89 (3.5) | 34 | 132 (4.3) | 42 | (−27 to 189) | 0.31 | −3 | (−78 to 44) | 0.89 |
| M vs. F | −31 | (−102 to 165) | 0.59 | 31 | (−13 to 113) | 0.22 | ||||
| IL-10 | HCMV− | HCMV+ | Comparison (Unadjusted) | Comparison (Adjusted) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Geo Mean (GSD) | N | Geo Mean (GSD) | Δ | 95%CI | p-Value | Δ | 95%CI | p-Value | |
| Unstimulated | ||||||||||
| ALL | 34 | 76 (8.0) | 73 | 96 (8.4) | 20 | (−35 to 148) | 0.59 | 136 | (−350 to 875) | 0.64 |
| Male | 17 | 149 (9.1) | 35 | 108 (9.2) | −40 | (−119 to 245) | 0.63 | 32 | (−596 to 1172) | 0.94 |
| Female | 17 | 39 (5.8) | 38 | 89 (8.1) | 49 | (−7 to 208) | 0.12 | 221 | (−1363 to 1368) | 0.58 |
| M vs. F | −90 | (−177 to 365) | 0.18 | −189 | (−988 to 1675) | 0.73 | ||||
| TLR2 (HKLM) | ||||||||||
| ALL | 35 | 4295 (3.3) | 73 | 3409 (4.3) | −886 | (−2258 to 1409) | 0.38 | 98 | (−243 to 563) | 0.62 |
| Male | 18 | 4786 (3.8) | 35 | 3366 (4.6) | −1420 | (−3273 to 2699) | 0.39 | −134 | (−530 to 437) | 0.60 |
| Female | 17 | 3830 (2.8) | 38 | 3441 (4.3) | −389 | (−2035 to 2768) | 0.75 | 283 | (−902 to 1120) | 0.33 |
| M vs. F | −1031 | (−3197 to 5038) | 0.64 | −418 | (−1024 to 728) | 0.27 | ||||
| TLR4 (LPS) | ||||||||||
| ALL | 32 | 3820 (3.7) | 72 | 3593 (4.0) | −227 | (−1744 to 2398) | 0.83 | 299 | (−348 to 1199) | 0.41 |
| Male | 16 | 5016 (2.5) | 34 | 3775 (3.6) | −1241 | (−3007 to 2078) | 0.38 | −533 | (−1260 to 553) | 0.29 |
| Female | 16 | 2910 (4.8) | 38 | 3475 (4.5) | 566 | (−1455 to 5393) | 0.69 | 860 | (−859 to 2297) | 0.062 |
| M vs. F | −1807 | (−4294 to 5484) | 0.40 | −1393 | (−2409 to −495) | 0.034 | ||||
| TLR5 (Flagellin) | ||||||||||
| ALL | 33 | 4409 (3.4) | 69 | 5412 (2.6) | 1003 | (−1040 to 4285) | 0.40 | 627 | (−226 to 1753) | 0.17 |
| Male | 17 | 4901 (3.4) | 34 | 5327 (2.5) | 425 | (−2125 to 5319) | 0.80 | 590 | (−637 to 2483) | 0.40 |
| Female | 16 | 3940 (3.6) | 35 | 5474 (2.8) | 1534 | (−1180 to 6917) | 0.35 | 661 | (−1569 to 2214) | 0.27 |
| M vs. F | −1109 | (−4366 to 7274) | 0.61 | −71 | (−1593 to 2630) | 0.94 | ||||
| TLR7/8 (CLO75) | ||||||||||
| ALL | 32 | 3941 (3.2) | 68 | 4198 (5.1) | 257 | (−1537 to 3391) | 0.82 | 350 | (−266 to 1148) | 0.29 |
| Male | 16 | 4382 (4.3) | 34 | 3786 (4.6) | −596 | (−2826 to 4825) | 0.75 | −580 | (−1416 to 684) | 0.32 |
| Female | 16 | 3544 (2.2) | 34 | 4534 (5.6) | 990 | (−1201 to 5231) | 0.47 | 1069 | (337 to 2195) | 0.005 |
| M vs. F | −1586 | (−4121 to 6083) | 0.49 | −1649 | (−2704 to −194) | 0.016 | ||||
| TNF:IL-10 | HCMV− | HCMV+ | Comparison (Unadjusted) | Comparison (Adjusted) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Geo Mean (GSD) | N | Geo Mean (GSD) | Δ | 95% CI | p-Value | Δ | 95% CI | p-Value | |
| Unstimulated | ||||||||||
| ALL | 34 | 0.95 (3.0) | 73 | 0.97 (3.0) | 0.02 | (−0.33 to 0.56) | 0.94 | 0.18 | (−0.08 to 0.54) | 0.19 |
| Male | 17 | 0.93 (2.8) | 35 | 1.03 (2.7) | 0.10 | (−0.36 to 0.92) | 0.75 | 0.33 | (−0.02 to 0.86) | 0.064 |
| Female | 17 | 0.97 (3.3) | 38 | 0.93 (3.3) | −0.04 | (−0.49 to 0.82) | 0.89 | 0.04 | (−1.08 to 0.60) | 0.86 |
| M vs. F | 0.14 | (−0.46 to 1.59) | 0.75 | 0.29 | (−0.18 to 1.18) | 0.26 | ||||
| TLR2 (HKLM) | ||||||||||
| ALL | 35 | 3.62 (2.8) | 73 | 2.57 (3.1) | −1.04 | (−1.93 to 0.32) | 0.12 | −0.74 | (−1.29 to 0.05) | 0.063 |
| Male | 18 | 2.52 (2.5) | 35 | 2.33 (2.5) | −0.19 | (−1.13 to 1.39) | 0.77 | −0.18 | (−0.90 to 1.04) | 0.72 |
| Female | 17 | 5.30 (2.8) | 38 | 2.77 (3.6) | −2.53 | (−3.80 to −0.19) | 0.038 | −1.48 | (−4.07 to −0.27) | 0.021 |
| M vs. F | 2.34 | (−1.06 to 5.20) | 0.16 | 1.30 | (−0.38 to 3.17) | 0.20 | ||||
| TLR4 (LPS) | ||||||||||
| ALL | 32 | 1.66 (3.3) | 72 | 1.29 (3.4) | −0.37 | (−0.88 to 0.46) | 0.32 | −0.09 | (−0.58 to 0.66) | 0.77 |
| Male | 16 | 1.22 (3.0) | 34 | 1.33 (3.0) | 0.11 | (−0.53 to 1.35) | 0.79 | 0.03 | (−0.58 to 1.17) | 0.95 |
| Female | 16 | 2.27 (3.4) | 38 | 1.26 (3.8) | −1.00 | (−1.64 to 0.29) | 0.10 | −0.22 | (−2.03 to 0.95) | 0.64 |
| M vs. F | 1.12 | (−0.29 to 3.27) | 0.17 | 0.25 | (−0.52 to 2.07) | 0.70 | ||||
| TLR5 (Flagellin) | ||||||||||
| ALL | 33 | 1.19 (3.3) | 69 | 0.82 (3.9) | −0.37 | (−0.70 to 0.18) | 0.16 | −0.12 | (−0.50 to 0.47) | 0.63 |
| Male | 17 | 1.05 (4.2) | 34 | 0.81 (3.7) | −0.24 | (−0.69 to 0.78) | 0.54 | 0.08 | (−0.39 to 0.98) | 0.79 |
| Female | 16 | 1.36 (2.5) | 35 | 0.83 (4.1) | −0.53 | (−0.91 to 0.16) | 0.11 | −0.36 | (−1.74 to 0.50) | 0.34 |
| M vs. F | 0.30 | (−0.22 to 1.73) | 0.64 | 0.44 | (−0.14 to 1.78) | 0.38 | ||||
| TLR7/8 (CLO75) | ||||||||||
| ALL | 32 | 1.00 (2.7) | 68 | 0.84 (3.2) | −0.16 | (−0.46 to 0.30) | 0.43 | −0.06 | (−0.42 to 0.45) | 0.78 |
| Male | 16 | 0.78 (2.9) | 34 | 0.71 (2.8) | −0.07 | (−0.40 to 0.55) | 0.76 | −0.09 | (−0.48 to 0.60) | 0.75 |
| Female | 16 | 1.29 (2.4) | 34 | 0.96 (3.5) | −0.33 | (−0.75 to 0.41) | 0.31 | −0.02 | (−1.24 to 0.92) | 0.95 |
| M vs. F | 0.26 | (−0.15 to 1.20) | 0.65 | −0.06 | (−0.58 to 1.05) | 0.86 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cox, M.; Adetifa, J.U.; Noho-Konteh, F.; Sanyang, L.C.; Drammeh, A.; Plebanski, M.; Whittle, H.C.; Rowland-Jones, S.L.; Robertson, I.; Flanagan, K.L. Sex-Differential Impact of Human Cytomegalovirus Infection on In Vitro Reactivity to Toll-Like Receptor 2, 4 and 7/8 Stimulation in Gambian Infants. Vaccines 2020, 8, 407. https://doi.org/10.3390/vaccines8030407
Cox M, Adetifa JU, Noho-Konteh F, Sanyang LC, Drammeh A, Plebanski M, Whittle HC, Rowland-Jones SL, Robertson I, Flanagan KL. Sex-Differential Impact of Human Cytomegalovirus Infection on In Vitro Reactivity to Toll-Like Receptor 2, 4 and 7/8 Stimulation in Gambian Infants. Vaccines. 2020; 8(3):407. https://doi.org/10.3390/vaccines8030407
Chicago/Turabian StyleCox, Momodou, Jane U. Adetifa, Fatou Noho-Konteh, Lady C. Sanyang, Abdoulie Drammeh, Magdalena Plebanski, Hilton C. Whittle, Sarah L. Rowland-Jones, Iain Robertson, and Katie L. Flanagan. 2020. "Sex-Differential Impact of Human Cytomegalovirus Infection on In Vitro Reactivity to Toll-Like Receptor 2, 4 and 7/8 Stimulation in Gambian Infants" Vaccines 8, no. 3: 407. https://doi.org/10.3390/vaccines8030407
APA StyleCox, M., Adetifa, J. U., Noho-Konteh, F., Sanyang, L. C., Drammeh, A., Plebanski, M., Whittle, H. C., Rowland-Jones, S. L., Robertson, I., & Flanagan, K. L. (2020). Sex-Differential Impact of Human Cytomegalovirus Infection on In Vitro Reactivity to Toll-Like Receptor 2, 4 and 7/8 Stimulation in Gambian Infants. Vaccines, 8(3), 407. https://doi.org/10.3390/vaccines8030407