Immunogenicity and Protective Efficacy of Influenza A DNA Vaccines Encoding Artificial Antigens Based on Conservative Hemagglutinin Stem Region and M2 Protein in Mice

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
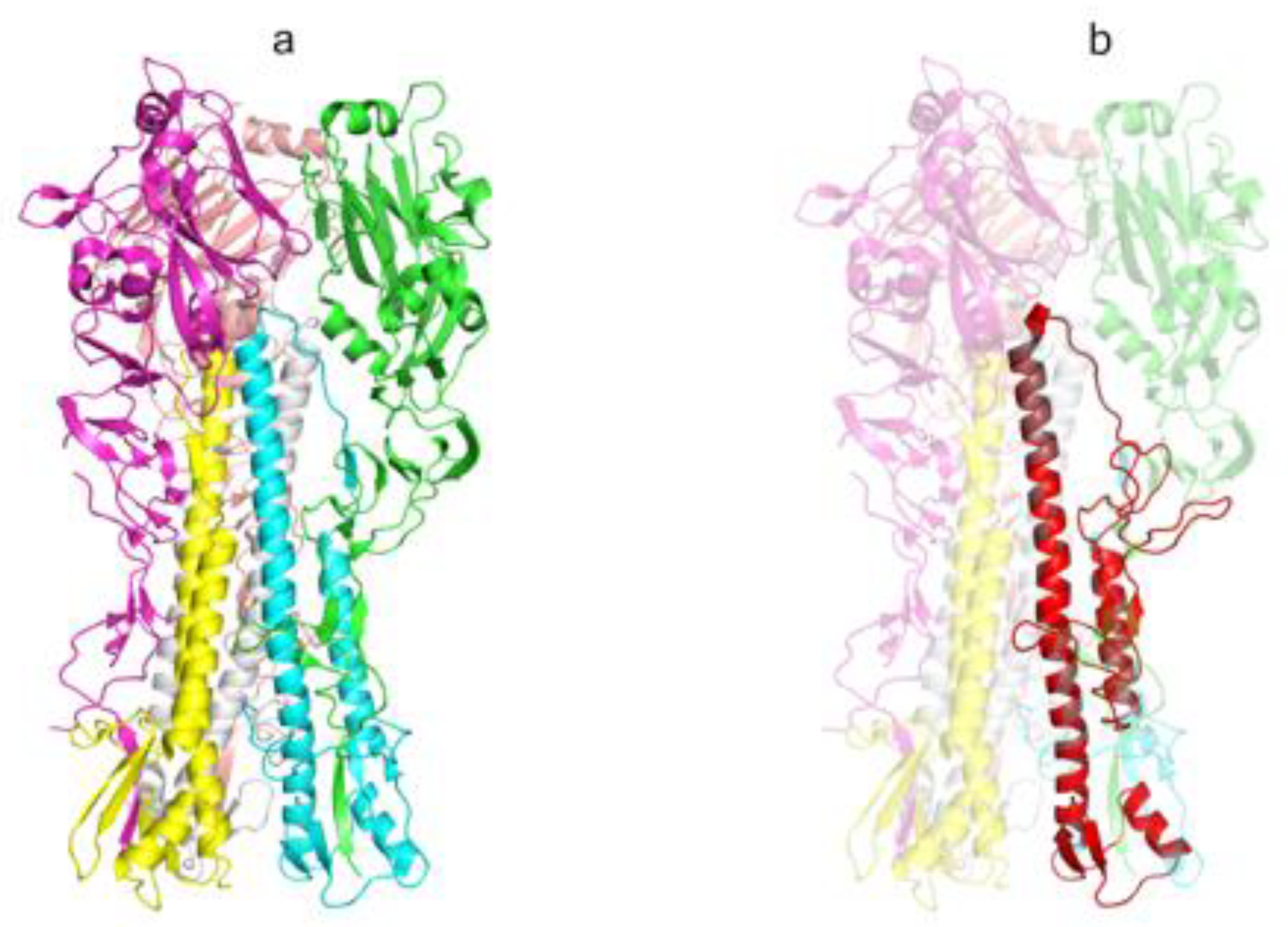
2.1. Design, Synthesis, and Cloning of the Target Genes
2.2. RT-PCR Detection of Target Gene Transcription
2.3. Ethics Statement
2.4. Immunization of Experimental Animals and Samples Collection
2.5. ELISpot Assay
2.6. Serum ELISA
2.7. Challenge Studies
2.8. Statistical Analysis and Software
3. Results
3.1. DNA Vaccine Constructions Encoding HA Stem and M2 Protein
3.2. Verification of Target Genes Transcription in Transfected Cells
3.3. Immunogenicity of DNA Vaccine Constructs Encoding the Target Antigens
- AgH1—mice were immunized with DNA-plasmid p-AgH1;
- AgH3—immunized with DNA-plasmid p-AgH3;
- AgM2—immunized with DNA-plasmid p-AgM2;
- Ag(H1 + H3 + M2)—immunized with a mixture of p-AgN1, p-AgN3, and p-AgM2 DNA-plasmids;
- Ag0—immunized with empty vector plasmid pcDNA3.1 (negative control);
- Intact—nonimmunized mice (negative control);
- H1N1pdm09—mice immunized with influenza A virus strain A/California/07/2009 (H1N1pdm09; positive control).
3.4. Protective Efficacy of the Developed DNA Vaccine Constructs
- Ag(H1 + H3 + M2)—mice immunized with a combination of DNA-plasmids p-AgH1+p-AgH3+p-AgM2 (75 µg/mice for each plasmid);
- H1N1—mice immunized with A/California/07/2009 (H1N1pdm09) virus (positive control);
- H3N2—mice immunized with A/Aichi/2/68 (H3N2) virus strain (positive control);
- pDNA3.1—mice immunized with empty vector plasmid pDNA3.1 (negative control; 200 µg/mice).
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kumar, A.; Meldgaard, T.S.; Bertholet, S. Novel platforms for the development of a universal influenza vaccine. Front. Immunol. 2018, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Estrada, L.D.; Schultz-Cherry, S. Development of a universal influenza vaccine. J. Immunol. 2019, 202, 392–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, R.D.; Altenburg, A.F.; Rimmelzwaan, G.F. Universal influenza vaccines, science fiction or soon reality? Expert Rev. Vaccines 2015, 14, 1299–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacheck, V.; Egorov, A.; Groiss, F.; Pfeiffer, A.; Fuereder, T.; Hoeflmayer, D.; Kundi, M.; Popow-Kraupp, T.; Redlberger-Fritz, M.; Mueller, C.A.; et al. A novel type of influenza vaccine: Safety and immunogenicity of replication-deficient influenza virus created by deletion of the interferon antagonist NS1. J. Infect. Dis. 2010, 201, 354–362. [Google Scholar] [CrossRef]
- Wang, W.L.; Li, R.Q.; Deng, Y.; Lu, N.; Chen, H.; Meng, X.; Wang, W.; Wang, X.P.; Yan, K.X.; Qi, X.R.; et al. Protective Efficacy of the Conserved NP, PB1, and M1 Proteins as Immunogens in DNA- and Vaccinia Virus-Based Universal Influenza A Virus Vaccines in Mice. Clin. Vaccine Immunol. 2015, 22, 618–630. [Google Scholar] [CrossRef] [Green Version]
- López-Macías, C. Virus-like particle (VLP)-based vaccines for pandemic influenza: Performance of a VLP vaccine during the 2009 influenza pandemic. Hum. Vaccines Immunother 2012, 8, 402–405. [Google Scholar] [CrossRef]
- Ravin, N.V.; Blokhina, E.A.; Kuprianov, V.V.; Stepanova, L.A.; Shaldjan, A.A.; Kovaleva, A.A.; Tsybalova, L.M.; Skryabin, K.G. Development of a candidate influenza vaccine based on virus-like particles displaying influenza M2e peptide into the immunodominant loop region of hepatitis B core antigen: Insertion of multiple copies of M2e increases immunogenicity and protective efficiency. Vaccine 2015, 33, 3392–3397. [Google Scholar] [CrossRef]
- Lee, L.Y.Y.; Izzard, L.; Hurt, A.C. A review of DNA vaccines against influenza. Front. Immunol. 2018, 9, 1568. [Google Scholar] [CrossRef] [Green Version]
- Asthagiri Arunkumar, G.; McMahon, M.; Pavot, V.; Aramouni, M.; Ioannou, A.; Lambe, T.; Gilbert, S.; Krammer, F. Vaccination with viral vectors expressing NP, M1 and chimeric hemagglutinin induces broad protection against influenza virus challenge in mice. Vaccine 2019, 37, 5567–5577. [Google Scholar] [CrossRef]
- Krammer, F.; García-Sastre, A.; Palese, P. Is it possible to develop a “universal” influenza virus vaccine: Potential target antigens and critical aspects for a universal influenza vaccine. Cold Spring Harb. Perspect. Biol. 2018, 10. [Google Scholar] [CrossRef]
- Kim, S.H.; Paldurai, A.; Samal, S.K. A novel chimeric Newcastle disease virus vectored vaccine against highly pathogenic avian influenza virus. Virology 2017, 503, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Xiang, K.; Ying, G.; Yan, Z.; Shanshan, Y.; Lei, Z.; Hongjun, L.; Maosheng, S. Progress on adenovirus-vectored universal influenza vaccines. Hum. Vaccines Immunother. 2015, 11, 1209–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffner, W.; van Buynder, P.; McNeil, S.; Osterhaus, A.D.M.E. Seasonal influenza immunisation: Strategies for older adults. Int. J. Clin. Pract. 2018, 72. [Google Scholar] [CrossRef] [PubMed]
- Koutsakos, M.; Nguyen, T.H.O.; Kedzierska, K. With a little help from T follicular helper friends: Humoral immunity to influenza vaccination. J. Immunol. 2019, 202, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Padilla-Quirarte, H.O.; Lopez-Guerrero, D.V.; Gutierrez-Xicotencatl, L.; Esquivel-Guadarrama, F. Protective antibodies against influenza proteins. Front. Immunol. 2019, 10, 1677. [Google Scholar] [CrossRef] [Green Version]
- Hashem, A.M. Prospects of HA-Based Universal Influenza Vaccine. Biomed Res. Int. 2015, 2015, 414637. [Google Scholar] [CrossRef]
- Van Der Lubbe, J.E.M.; Verspuij, J.W.A.; Huizingh, J.; Schmit-Tillemans, S.P.R.; Tolboom, J.T.B.M.; Dekking, L.E.H.A.; Kwaks, T.; Brandenburg, B.; Meijberg, W.; Zahn, R.C.; et al. Mini-HA is superior to full length hemagglutinin immunization in inducing stem-specific antibodies and protection against group 1 influenza virus challenges in mice. Front. Immunol. 2018, 9, 2350. [Google Scholar] [CrossRef]
- Yassine, H.M.; Boyington, J.C.; McTamney, P.M.; Wei, C.J.; Kanekiyo, M.; Kong, W.P.; Gallagher, J.R.; Wang, L.S.; Zhang, Y.; Joyce, M.G.; et al. Hemagglutinin-stem nanoparticles generate heterosubtypic influenza protection. Nat. Med. 2015, 21, 1065–1070. [Google Scholar] [CrossRef]
- Mallajosyula, V.V.A.; Citron, M.; Ferrara, F.; Lu, X.H.; Callahan, C.; Heidecker, G.J.; Sarma, S.P.; Flynn, J.A.; Temperton, N.J.; Liang, X.P.; et al. Influenza hemagglutinin stem-fragment immunogen elicits broadly neutralizing antibodies and confers heterologous protection. Proc. Natl. Acad. Sci. USA 2014, 111, E2514–E2523. [Google Scholar] [CrossRef] [Green Version]
- Tao, W.Q.; Gill, H.S. M2e-immobilized gold nanoparticles as influenza A vaccine: Role of soluble M2e and longevity of protection. Vaccine 2015, 33, 2307–2315. [Google Scholar] [CrossRef] [Green Version]
- Pizzolla, A.; Wakim, L.M. Memory T cell dynamics in the lung during influenza virus infection. J. Immunol. 2019, 202, 374–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutsakos, M.; Illing, P.T.; Nguyen, T.H.O.; Mifsud, N.A.; Crawford, J.C.; Rizzetto, S.; Eltahla, A.A.; Clemens, E.B.; Sant, S.; Chua, B.Y.; et al. Human CD8(+) T cell cross-reactivity across influenza A, B and C viruses. Nat. Immunol. 2019, 20, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Kreijtz, J.; Geelhoed-Mieras, M.M.; van Amerongen, G.; Verburgh, R.J.; van Trierum, S.E.; Kuiken, T.; Fouchier, R.A.M.; Osterhaus, A.; Rimmelzwaan, G.F. Vaccination against Seasonal Influenza A/H3N2 Virus Reduces the Induction of Heterosubtypic Immunity against Influenza A/H5N1 Virus Infection in Ferrets. J. Virol. 2011, 85, 2695–2702. [Google Scholar] [CrossRef] [Green Version]
- Holzer, B.; Martini, V.; Edmans, M.; Tchilian, E. T and B Cell Immune Responses to influenza viruses in pigs. Front. Immunol. 2019, 10, 98. [Google Scholar] [CrossRef]
- Thomas, P.G.; Keating, R.; Hulse-Post, D.J.; Doherty, P.C. Cell-mediated protection in influenza infection. Emerg. Infect. Dis. 2006, 12, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Hillaire, M.L.B.; Osterhaus, A.; Rimmelzwaan, G.F. Induction of Virus-Specific Cytotoxic T Lymphocytes as a Basis for the Development of Broadly Protective Influenza Vaccines. J. Biomed. Biotechnol. 2011, 2011, 939860. [Google Scholar] [CrossRef]
- McKinstry, K.K.; Strutt, T.M.; Kuang, Y.; Brown, D.M.; Sell, S.; Dutton, R.W.; Swain, S.L. Memory CD4 + T cells protect against influenza through multiple synergizing mechanisms. J. Clin. Investig. 2012, 122, 2847–2856. [Google Scholar] [CrossRef] [Green Version]
- Altenburg, A.F.; Rimmelzwaan, G.F.; de Vries, R.D. Virus-specific T cells as correlate of (cross-)protective immunity against influenza. Vaccine 2015, 33, 500–506. [Google Scholar] [CrossRef]
- Stachyra, A.; Gora-Sochacka, A.; Sirko, A. DNA vaccines against influenza. Acta Biochim. Pol. 2014, 61, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Ulmer, J.B. Influenza DNA vaccines. Vaccine 2002, 20, S74–S76. [Google Scholar] [CrossRef]
- Tompkins, S.M.; Zhao, Z.S.; Lo, C.Y.; Misplon, J.A.; Liu, T.; Ye, Z.P.; Hogan, R.J.; Wu, Z.Q.; Benton, K.A.; Tumpey, T.M.; et al. Matrix protein 2 vaccination and protection against influenza viruses, including subtype H5N1. Emerg. Infect. Dis. 2007, 13, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Bragstad, K.; Vinner, L.; Hansen, M.S.; Nielsen, J.; Fomsgaard, A. A polyvalent influenza A DNA vaccine induces heterologous immunity and protects pigs against pandemic A(H1N1)pdm09 virus infection. Vaccine 2013, 31, 2281–2288. [Google Scholar] [CrossRef] [PubMed]
- Price, G.E.; Soboleski, M.R.; Lo, C.Y.; Misplon, J.A.; Pappas, C.; Houser, K.V.; Tumpey, T.M.; Epstein, S.L. Vaccination focusing immunity on conserved antigens protects mice and ferrets against virulent H1N1 and H5N1 influenza A viruses. Vaccine 2009, 27, 6512–6521. [Google Scholar] [CrossRef] [PubMed]
- Koday, M.T.; Leonard, J.A.; Munson, P.; Forero, A.; Koday, M.; Bratt, D.L.; Fuller, J.T.; Murnane, R.; Qin, S.L.; Reinhart, T.A.; et al. Multigenic DNA vaccine induces protective cross-reactive T cell responses against heterologous influenza virus in nonhuman primates. PLoS ONE 2017, 12, e0189780. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yu, H.; Yang, F.R.; Huang, M.; Ma, J.H.; Tong, G.Z. Protective efficacy of a broadly cross-reactive swine influenza DNA vaccine encoding M2e, cytotoxic T lymphocyte epitope and consensus H3 hemagglutinin. Virology J. 2012, 9, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Morrow, M.P.; Chu, J.S.; Racine, T.; Reed, C.C.; Khan, A.S.; Broderick, K.E.; Kim, J.J.; Kobinger, G.P.; Sardesai, N.Y.; et al. Broad cross-protective anti-hemagglutination responses elicited by influenza microconsensus DNA vaccine. Vaccine 2018, 36, 3079–3089. [Google Scholar] [CrossRef]
- Villalobos, A.; Welch, M.; Minshull, J. In silico design of functional DNA constructs. In Methods in Molecular Biology; Springer Protocols: Berlin/Heidelberg, Germany, 2012; Volume 852, pp. 197–213. [Google Scholar]
- Zhang, Y.; Aevermann, B.D.; Anderson, T.K.; Burke, D.F.; Dauphin, G.; Gu, Z.P.; He, S.; Kumar, S.; Larsen, C.N.; Lee, A.J.; et al. Influenza Research Database: An integrated bioinformatics resource for influenza virus research. Nucleic Acids Res. 2017, 45, D466–D474. [Google Scholar] [CrossRef] [Green Version]
- Pages, H.; Aboyoun, P.; Gentleman, R.; DebRoy, S. Biostrings: String Objects Representing Biological Sequences, and Matching Algorithms. R Package Version 2.42.1. Available online: http://www.citeulike.org/user/heliopais/article/11644278 (accessed on 23 April 2016).
- R Development Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: the R Foundation for Statistical Computing. Available online: https://www.r-project.org (accessed on 22 June 2020).
- Therneau, T.M. A Package for Survival Analysis in S. version 2.39-5. Available online: https://CRAN.R-project.org/package=survival (accessed on 10 April 2019).
- Kassambara, A.; Kosinski, M.; Biecek, P.; Fabian, S. Survminer: Drawing Survival Curves Using “ggplot2”. R Package Version 0.3.1. Available online: https://CRAN.R-project.org/package=survminer (accessed on 10 April 2019).
- Squires, R.B.; Noronha, J.; Hunt, V.; García-Sastre, A.; Macken, C.; Baumgarth, N.; Suarez, D.; Pickett, B.E.; Zhang, Y.; Larsen, C.N.; et al. Influenza Research Database: An integrated bioinformatics resource for influenza research and surveillance. Influ. Other Respir. Viruses 2012, 6, 404–416. [Google Scholar] [CrossRef]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- DeLano, W.L.; Bromberg, S. PyMOL User’s Guide; DeLano Scientific LLC: San Francisco, CA, USA, 2004. [Google Scholar]
- Lundegaard, C.; Lamberth, K.; Harndahl, M.; Buus, S.; Lund, O.; Nielsen, M. NetMHC-3.0: accurate web accessible predictions of human, mouse and monkey MHC class I affinities for peptides of length 8-11. Nucleic Acids Res. 2008, 36, W509–W512. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.; Lundegaard, C.; Blicher, T.; Peters, B.; Sette, A.; Justesen, S.; Buus, S.; Lund, O. Quantitative predictions of peptide binding to any HLA-DR molecule of known sequence: NetMHCIIpan. PLoS Comput. Biol. 2008, 4, e1000107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Cho, K.J.; Fiers, W.; Saelens, X. M2e-based universal influenza a vaccines. Vaccines 2015, 3, 105–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottlieb, T.; Ben-Yedidia, T. Epitope-based approaches to a universal influenza vaccine. J. Autoimmun. 2014, 54, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Van Drunen Littel-Van Den Hurk, S.; Hannaman, D. Electroporation for DNA immunization: Clinical application. Expert Rev. Vaccines 2010, 9, 503–517. [Google Scholar] [CrossRef]
- Sardesai, N.Y.; Weiner, D.B. Electroporation delivery of DNA vaccines: prospects for success. Current Opin. Immunol. 2011, 23, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Carragher, D.M.; Kaminski, D.A.; Moquin, A.; Hartson, L.; Randall, T.D. A novel role for non-neutralizing antibodies against nucleoprotein in facilitating resistance to influenza virus. J. Immunol. 2008, 181, 4168–4176. [Google Scholar] [CrossRef] [Green Version]
- Varečková, E.; Mucha, V.; Wharton, S.A.; Kostolanský, F. Inhibition of fusion activity of influenza A haemagglutinin mediated by HA2-specific monoclonal antibodies. Arch. Virol. 2003, 148, 469–486. [Google Scholar] [CrossRef]
- Terajima, M.; Co, M.D.T.; Cruz, J.; Ennis, F.A. High Antibody-Dependent Cellular Cytotoxicity Antibody Titers to H5N1 and H7N9 Avian Influenza A Viruses in Healthy US Adults and Older Children. J. Infect. Dis. 2015, 212, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Wren, L.; Kent, S.J. HIV vaccine efficacy trial: Glimmers of hope and the potential role of antibody-dependent cellular cytotoxicity. Hum. Vaccines 2011, 7, 466–473. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Annotation |
|---|---|
| F: 5′-ACTGTTGACACAGTGCTGGAAAAGAAT-3′ R: 5′-TTTTCATCTTTGTTGAATTCTTTTCC-3′ | Primers pair to detect mRNA encoding antigen AgH1 |
| F: 5′-GCTTTGAGCTACATTCTATGTCTGG-3′ R: 5′-GGTCCTGAATTCTCCCTTCATCTTC-3′ | Primers for detection of AgH3 mRNA |
| F: 5′-GAATGGGGGTGCAGATGCAACGATTC-3′ R: 5′-CAACTCTATGCTGACAAAATGACTGTC-3′ | Primers for AgM2 mRNA detection |
| Peptides in Composition of the Designed Antigens | ||
|---|---|---|
| AgH1 | AgH3 | AgM2 |
| KYVRSAKLR | LFERTKKQL | RGPSTEGVP |
| LYEKVKSQL | HDVYRDEAL | ETPIRNEWG |
| FYHKADNEA | RYVKQNTLK | LLTEVETPI |
| AKLRMVTGL | KPFQNVNRI | |
| SHGSANSSL | LENQHTIDL | |
| KSGYKDWIL | ||
| IEVTNATEL | ||
| Antigen/Number of Mice/Reverse Titer in ELISA against A/California/07/09 (H1N1) | ||||||||||||
| Ag(H1 + H3 + M2) | H1N1 | H3N2 | pcDNA | |||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 1 | 1 | 1 | 2 | 3 |
| 1600 | 1600 | 1600 | 1600 | 3200 | 3200 | 3200 | 3200 | 12,800 | 3200 | 200 | 200 | 200 |
| Antigen/Number of mice/Reverse Titer in ELISA against A/Aichi/02/68 (H3N2) | ||||||||||||
| Ag(H1 + H3 + M2) | H1N1 | H3N2 | pcDNA | |||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 1 | 1 | 1 | 2 | 3 |
| 1600 | 3200 | 1600 | 3200 | 6400 | 6400 | 3200 | 6400 | 6400 | 12800 | 200 | 400 | 400 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazhan, S.; Antonets, D.; Starostina, E.; Ilyicheva, T.; Kaplina, O.; Marchenko, V.; Durymanov, A.; Oreshkova, S.; Karpenko, L. Immunogenicity and Protective Efficacy of Influenza A DNA Vaccines Encoding Artificial Antigens Based on Conservative Hemagglutinin Stem Region and M2 Protein in Mice. Vaccines 2020, 8, 448. https://doi.org/10.3390/vaccines8030448
Bazhan S, Antonets D, Starostina E, Ilyicheva T, Kaplina O, Marchenko V, Durymanov A, Oreshkova S, Karpenko L. Immunogenicity and Protective Efficacy of Influenza A DNA Vaccines Encoding Artificial Antigens Based on Conservative Hemagglutinin Stem Region and M2 Protein in Mice. Vaccines. 2020; 8(3):448. https://doi.org/10.3390/vaccines8030448
Chicago/Turabian StyleBazhan, Sergei, Denis Antonets, Ekaterina Starostina, Tatyana Ilyicheva, Olga Kaplina, Vasiliy Marchenko, Alexander Durymanov, Svetlana Oreshkova, and Larisa Karpenko. 2020. "Immunogenicity and Protective Efficacy of Influenza A DNA Vaccines Encoding Artificial Antigens Based on Conservative Hemagglutinin Stem Region and M2 Protein in Mice" Vaccines 8, no. 3: 448. https://doi.org/10.3390/vaccines8030448
APA StyleBazhan, S., Antonets, D., Starostina, E., Ilyicheva, T., Kaplina, O., Marchenko, V., Durymanov, A., Oreshkova, S., & Karpenko, L. (2020). Immunogenicity and Protective Efficacy of Influenza A DNA Vaccines Encoding Artificial Antigens Based on Conservative Hemagglutinin Stem Region and M2 Protein in Mice. Vaccines, 8(3), 448. https://doi.org/10.3390/vaccines8030448