A Herpesvirus of Turkey-Based Vector Vaccine Reduces Transmission of Newcastle Disease Virus in Commercial Broiler Chickens with Maternally Derived Antibodies

 ,
,  ,
,  , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
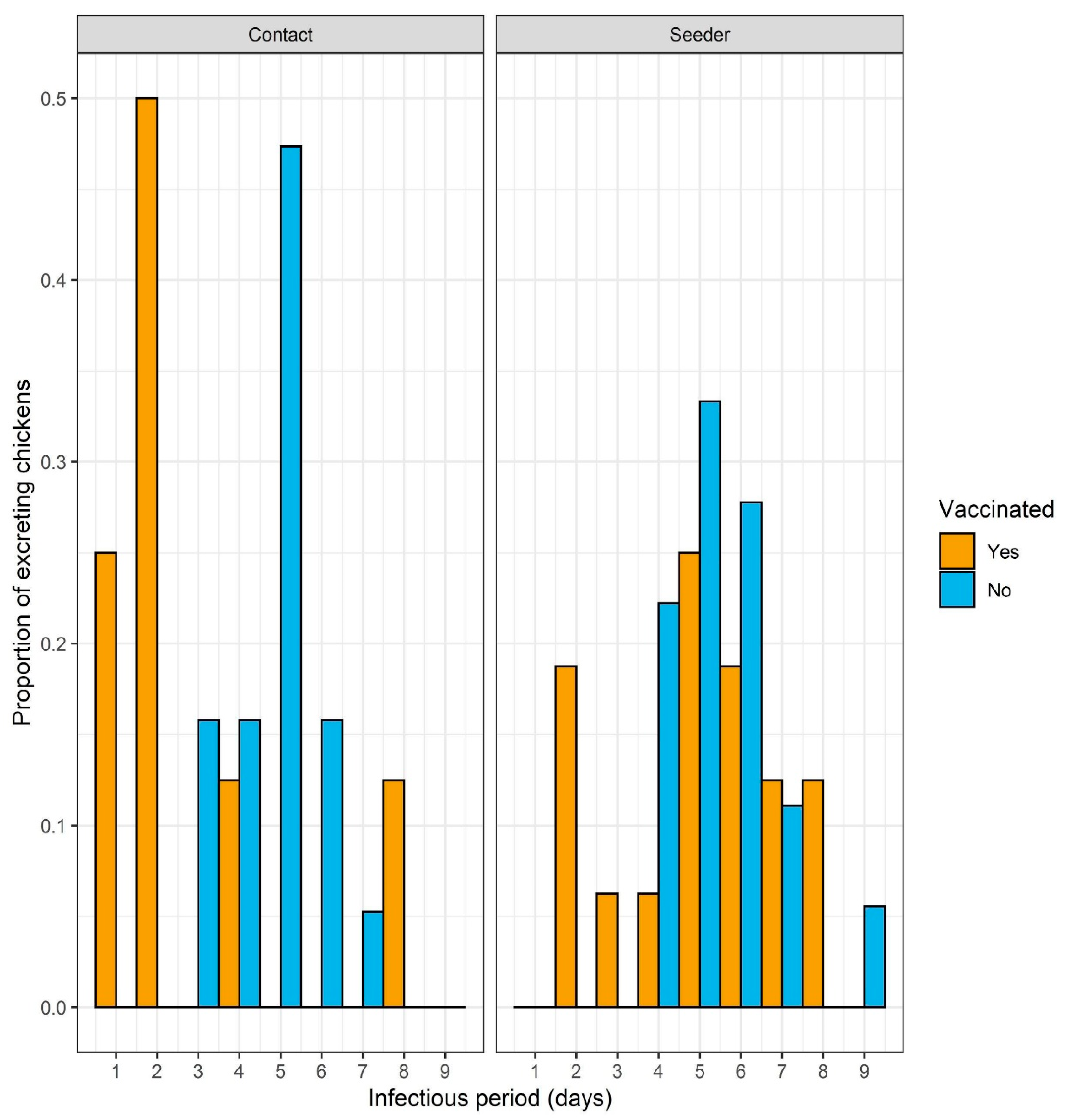{kind=link}
Abstract
Share and Cite
Tatár-Kis, T.; Fischer, E.A.J.; Cazaban, C.; Walkó-Kovács, E.; Homonnay, Z.G.; Velkers, F.C.; Palya, V.; Stegeman, J.A. A Herpesvirus of Turkey-Based Vector Vaccine Reduces Transmission of Newcastle Disease Virus in Commercial Broiler Chickens with Maternally Derived Antibodies. Vaccines 2020, 8, 614. https://doi.org/10.3390/vaccines8040614
Tatár-Kis T, Fischer EAJ, Cazaban C, Walkó-Kovács E, Homonnay ZG, Velkers FC, Palya V, Stegeman JA. A Herpesvirus of Turkey-Based Vector Vaccine Reduces Transmission of Newcastle Disease Virus in Commercial Broiler Chickens with Maternally Derived Antibodies. Vaccines. 2020; 8(4):614. https://doi.org/10.3390/vaccines8040614
Chicago/Turabian StyleTatár-Kis, Timea, Egil A.J. Fischer, Christophe Cazaban, Edit Walkó-Kovács, Zalan G. Homonnay, Francisca C. Velkers, Vilmos Palya, and J. Arjan Stegeman. 2020. "A Herpesvirus of Turkey-Based Vector Vaccine Reduces Transmission of Newcastle Disease Virus in Commercial Broiler Chickens with Maternally Derived Antibodies" Vaccines 8, no. 4: 614. https://doi.org/10.3390/vaccines8040614
APA StyleTatár-Kis, T., Fischer, E. A. J., Cazaban, C., Walkó-Kovács, E., Homonnay, Z. G., Velkers, F. C., Palya, V., & Stegeman, J. A. (2020). A Herpesvirus of Turkey-Based Vector Vaccine Reduces Transmission of Newcastle Disease Virus in Commercial Broiler Chickens with Maternally Derived Antibodies. Vaccines, 8(4), 614. https://doi.org/10.3390/vaccines8040614