A DNA Vaccine Encoding Plasmodium falciparum PfRH5 in Cationic Liposomes for Dermal Tattooing Immunization

Abstract

1. Introduction
2. Materials and Methods
2.1. Cloning of Plasmid Constructs
2.2. Liposome Preparation and Entrapment of Plasmid DNA
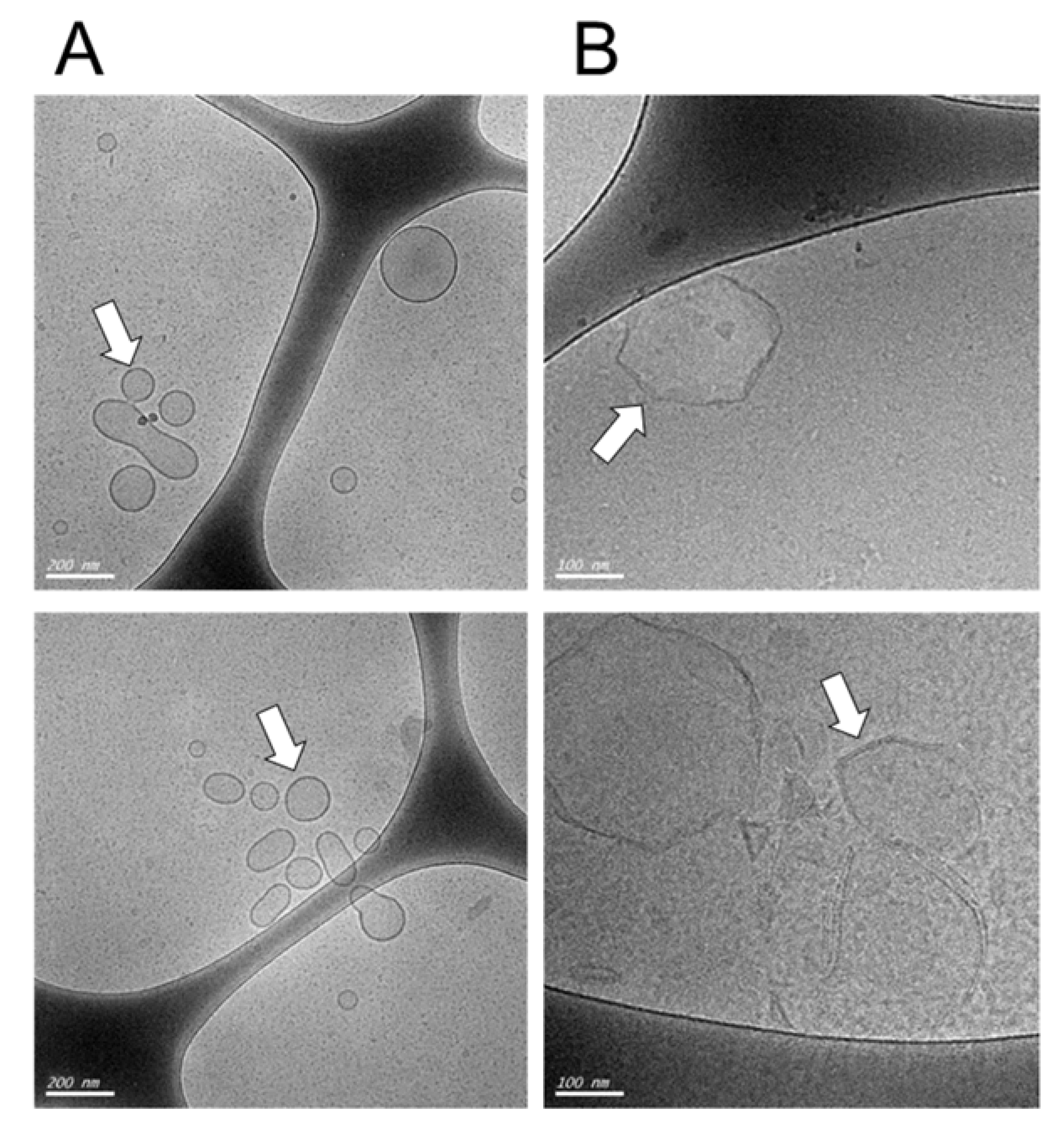
2.3. Cryoelectron Microscopy
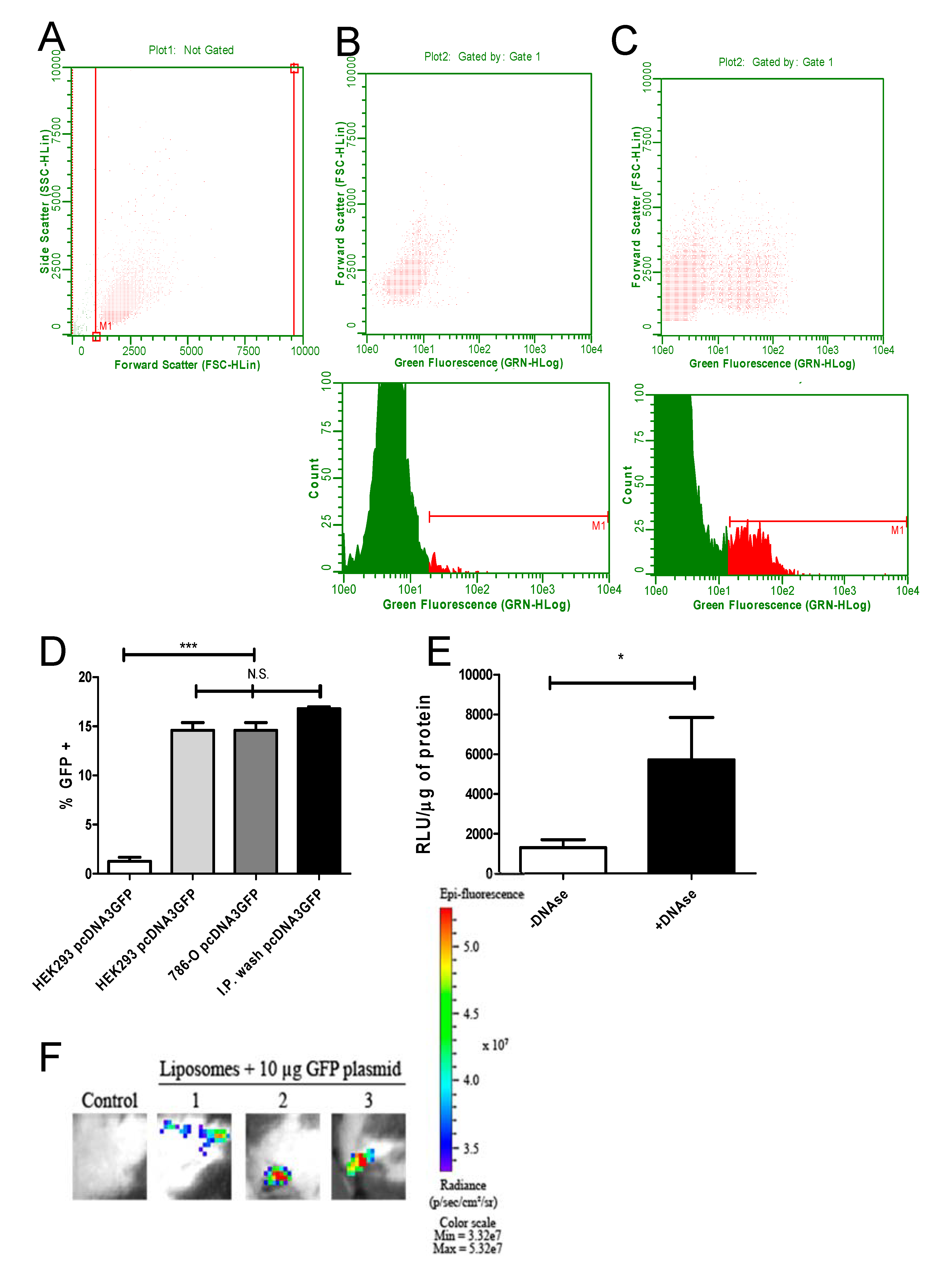
2.4. Liposome Transfection In Vitro and In Vivo Assay
2.5. Intradermal Immunization
2.6. P. falciparum Culture and Invasion Inhibition Assays
2.7. ELISA and Western Blots
2.8. Statistical Analyses
3. Results
3.1. Characteristics and Morphology of DDAB/DC-Chol Liposomes and Liposomes/DNA Complexes
3.2. Transfection In Vitro and In Vivo Reveals Efficient Antigen Production in Target Cells
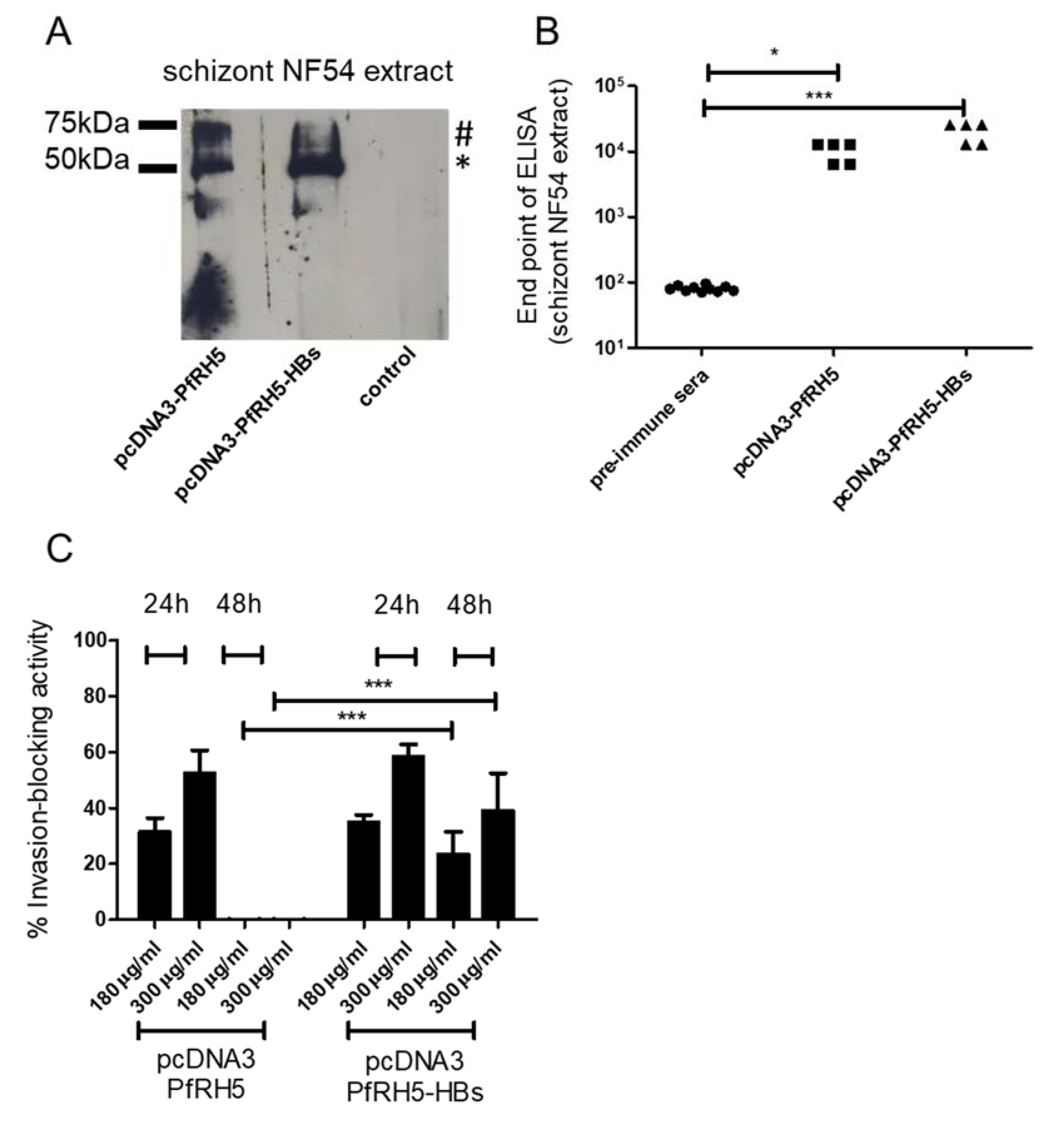
3.3. Intradermal Tattooing Immunization against PfRH5-HBs Generates Functional Antibodies against P. falciparum
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Torchilin, V.P. Recent advances with liposomes as pharmaceutical carriers. Nat. Rev. Drug Discov. 2005, 4, 145–160. [Google Scholar] [CrossRef]
- Singh, H.; Thompson, A.; Liu, L.; Corredig, M. Liposomes as food ingredients and nutraceutical delivery systems. In Encapsulation Technologies and Delivery Systems for Food Ingredients and Nutraceuticals; Woodhead Publishing: Sawston, UK, 2012; pp. 287–318. ISBN 9780857091246. [Google Scholar]
- Wesołowska, O.; Michalak, K.; Maniewska, J.; Hendrich, A.B. Giant unilamellar vesicles—A perfect tool to visualize phase separation and lipid rafts in model systems. Acta Biochim. Pol. 2009, 56, 33–39. [Google Scholar]
- Irvine, D.J.; Swartz, M.A.; Szeto, G.L. Engineering synthetic vaccines using cues from natural immunity. Nat. Mater. 2013, 12, 978–990. [Google Scholar] [CrossRef] [PubMed]
- Shim, G.; Kim, M.-G.; Park, J.Y.; Oh, Y.-K. Application of cationic liposomes for delivery of nucleic acids. Asian J. Pharm. Sci. 2013, 8, 72–80. [Google Scholar] [CrossRef]
- Wolff, J.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef]
- Tang, D.C.; DeVit, M.; Johnston, S.A. Genetic immunization is a simple method for eliciting an immune response. Nature 1992, 356, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.L.; Demeneix, B.A.; Quantin, B.; Coulombe, J.; Whalen, R.G. Plasmid DNA Is Superior to Viral Vectors for Direct Gene Transfer into Adult Mouse Skeletal Muscle. Hum. Gene Ther. 1993, 4, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Introduction of foreign genes into tissues of living mice by DNA-coated microprojectiles. Genetics 1991, 88, 2726–2730.
- Richie, T.L.; Charoenvit, Y.; Wang, R.; Epstein, J.E.; Hedstrom, R.C.; Kumar, S.; Luke, T.C.; Freilich, D.A.; Aguiar, J.C.; Sacci, J.B.; et al. Clinical trial in healthy malaria-naïve adults to evaluate the safety, tolerability, immunogenicity and efficacy of MuStDO5, a five-gene, sporozoite/hepatic stage Plasmodium falciparum DNA vaccine combined with escalating dose human GM-CSF DNA. Hum. Vaccines Immunother. 2012, 8, 1564–1584. [Google Scholar] [CrossRef] [PubMed]
- Pokorna, D.; Rubio, I.; Müller, M. DNA-vaccination via tattooing induces stronger humoral and cellular immune responses than intramuscular delivery supported by molecular adjuvants. Genet. Vaccines Ther. 2008, 6, 4. [Google Scholar] [CrossRef]
- Chiu, Y.-N.; Sampson, J.M.; Jiang, X.; Zolla-Pazner, S.B.; Kong, X.-P. Skin Tattooing as A Novel Approach for DNA Vaccine Delivery. J. Vis. Exp. 2012, 68, 50032. [Google Scholar] [CrossRef] [PubMed]
- Crosnier, C.; Bustamante, L.Y.; Bartholdson, S.J.; Bei, A.K.; Theron, M.; Uchikawa, M.; Mboup, S.; Ndir, O.; Kwiatkowski, D.P.; Duraisingh, M.T.; et al. Basigin is a receptor essential for erythrocyte invasion by Plasmodium falciparum. Nature 2011, 480, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Fotoran, W.L.; Santangelo, R.; de Miranda, B.N.M.; Irvine, D.J.; Wunderlich, G. DNA-Loaded Cationic Liposomes Efficiently Function as a Vaccine against Malarial Proteins. Mol. Ther. Methods Clin. Dev. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; ISBN 13: 978-0879693091. [Google Scholar]
- Fotoran, W.L.; Santangelo, R.M.; Medeiros, M.M.; Colhone, M.; Ciancaglini, P.; Barboza, R.; Marinho, C.R.F.; Stábeli, R.G.; Wunderlich, G. Liposomes loaded with P. falciparum merozoite-derived proteins are highly immunogenic and produce invasion-inhibiting and anti-toxin antibodies. J. Control. Release 2015, 217, 121–127. [Google Scholar] [CrossRef]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Lelievre, J.; Berry, A. An alternative method for Plasmodium culture synchronization. Exp. Parasitol. 2005, 109, 195–197. [Google Scholar] [CrossRef]
- Lambros, C.; Vanderberg, J.P. Synchronization of Plasmodium falciparum erythrocytic stages in culture. J Parasitol. 1979, 65, 418–420. [Google Scholar] [CrossRef]
- Wilson, D.W.; Crabb, B.S.; Beeson, J.G. Development of fluorescent Plasmodium falciparum for in vitro growth inhibition assays. Malar. J. 2010, 9, 152. [Google Scholar] [CrossRef]
- Amanat, F.; Krammer, F. SARS-CoV-2 Vaccines: Status Report. Immunity 2020, 52, 583–589. [Google Scholar] [CrossRef]
- Beasley, D.W.C. New international guidance on quality, safety and efficacy of DNA vaccines. Npj. Vaccines 2020, 5, 53. [Google Scholar] [CrossRef]
- Lu, S. Timely development of vaccines against SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 542–544. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, J.J.; Wahren, B.; Liu, M.A. DNA Vaccines: Progress and Challenges. J. Immunol. 2005, 175, 633–639. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Jin, S.; Xu, J.; Wang, P.C.; Liang, X.J.; Zhang, X. Engineered biomaterials for development of nucleic acid vaccines. Biomater. Res. 2015, 19, 1–9. [Google Scholar] [CrossRef]
- Geall, A.J.; Verma, A.; Otten, G.R.; Shaw, C.A.; Hekele, A.; Banerjee, K.; Cu, Y.; Beard, C.W.; Brito, L.A.; Krucker, T.; et al. Nonviral delivery of self-amplifying RNA vaccines. Proc. Natl. Acad. Sci. USA 2012, 109, 14604–14609. [Google Scholar] [CrossRef]
- Rodriguez-Gascon, A.; del Pozo-Rodríguez, A.; Solinís, M.A. Development of nucleic acid vaccines: Use of self-amplifying RNA in lipid nanoparticles. Int. J. Nanomed. 2014, 9, 1833. [Google Scholar] [CrossRef] [PubMed]
- Oosterhuis, K.; van den Berg, J.H.; Schumacher, T.N.; Haanen, J.B.A.G. DNA Vaccines and Intradermal Vaccination by DNA Tattooing. In Current Topics in Microbiology and Immunology; Springer: Berlin, Germany, 2010; Volume 351, pp. 221–250. [Google Scholar]
- DeMuth, P.C.; Min, Y.; Huang, B.; Kramer, J.A.; Miller, A.D.; Barouch, D.H.; Hammond, P.T.; Irvine, D.J. Polymer multilayer tattooing for enhanced DNA vaccination. Nat. Mater. 2013, 12, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, J.H.; Nuijen, B.; Beijnen, J.H.; Vincent, A.; van Tinteren, H.; Kluge, J.; Woerdeman, L.A.E.; Hennink, W.E.; Storm, G.; Schumacher, T.; et al. Optimization of intradermal vaccination by DNA tattooing in human skin. Hum. Gene Ther. 2009, 20, 181–189. [Google Scholar] [CrossRef]
- Ragotte, R.J.; Higgins, M.K.; Draper, S.J. The RH5-CyRPA-Ripr Complex as a Malaria Vaccine Target. Trends Parasitol. 2020, 36, 545–559. [Google Scholar] [CrossRef]
- Bustamante, L.Y.; Bartholdson, S.J.; Crosnier, C.; Campos, M.G.; Wanaguru, M.; Nguon, C.; Kwiatkowski, D.P.; Wright, G.J.; Rayner, J.C. A full-length recombinant Plasmodium falciparum PfRH5 protein induces inhibitory antibodies that are effective across common PfRH5 genetic variants. Vaccine 2013, 31, 373–379. [Google Scholar] [CrossRef]
- Douglas, A.D.; Baldeviano, G.C.; Jing, J.; Miura, K.; Diouf, A.; Zenonos, Z.A.; Ventocilla, J.A.; Silk, S.E.; Marshall, J.M.; Alanine, D.G.W.; et al. A defined mechanistic correlate of protection against Plasmodium falciparum malaria in non-human primates. Nat. Comm. 2019, 10, 1953–1961. [Google Scholar] [CrossRef]
- Krishnaswamy, J.K.; Gowthaman, U.; Zhang, B.; Mattsson, J.; Szeponik, L.; Liu, D.; Wu, R.; White, T.; Calabro, S.; Xu, L.; et al. Migratory CD11b+ conventional dendritic cells induce T follicular helper cell–dependent antibody responses. Sci. Immunol. 2017, 2, eaam9169. [Google Scholar] [CrossRef] [PubMed]
- Boopathy, A.V.; Mandal, A.; Kulp, D.W.; Menis, S.; Bennett, N.R.; Watkins, H.C.; Wang, W.; Martin, J.T.; Thai, N.T.; He, Y.; et al. Enhancing humoral immunity via sustained-release implantable microneedle patch vaccination. Proc. Natl. Acad. Sci. USA 2019, 116, 16473–16478. [Google Scholar] [CrossRef]
- Douglas, A.D.; Williams, A.R.; Illingworth, J.J.; Kamuyu, G.; Biswas, S.; Goodman, A.L.; Wyllie, D.H.; Crosnier, C.; Miura, K.; Wright, G.J.; et al. The blood-stage malaria antigen PfRH5 is susceptible to vaccine-inducible cross-strain neutralizing antibody. Nat. Commun. 2011, 2, 601. [Google Scholar] [CrossRef] [PubMed]
- Obaldia, N., 3rd; Stockelman, M.G.; Otero, W.; Cockrill, J.A.; Ganeshan, H.; Abot, E.N.; Zhang, J.; Limbach, K.; Charoenvit, Y.; Doolan, D.L.; et al. Heterologous Immunization Regimen Partially Protects Aotus Monkeys against Blood-Stage Challenge. Clin Vaccine Immunol. 2017, 24, e00539-16. [Google Scholar] [CrossRef]
- Stoute, J.A.; Slaoui, M.; Heppner, D.G.; Momin, P.; Kester, K.E.; Desmons, P.; Wellde, B.T.; Garçon, N.; Krzych, U.; Marchand, M.; et al. A preliminary evaluation of a recombinant circumsporozoite protein vaccine against Plasmodium falciparum malaria. N. Engl. J. Med. 1997, 336, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, G.; del Portillo, H.A. Biochemical and immunological properties of a viral hybrid particle expressing the Plasmodium vivax merozoite surface protein 1 C-terminal region. Mol. Med. 2000, 6, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, G.; Moura, I.C.; del Portillo, H.A.A. Genetic immunization of BALB/c mice with a plasmid bearing the gene coding for a hybrid merozoite surface protein 1-hepatitis B virus surface protein fusion protects mice against lethal Plasmodium chabaudi chabaudi PC1 infection. Infect. Immun. 2000, 68, 5839–5845. [Google Scholar] [CrossRef]
- Sahin, U.; Karikó, K.; Türeci, Ö. mRNA-based therapeutics — developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Pelc, R.S.; Muramatsu, H.; Andersen, H.; DeMaso, C.R.; Dowd, K.A.; Sutherland, L.L.; Scearce, R.M.; Parks, R.; et al. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature 2017, 543, 248–251. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Liposomes (DDAB+DC Cholesterin) | 8 nM of Lipid/µg of DNA Content |
|---|---|---|
| Zeta Potential (mV) Average ± SD | 14.15 ± 1.22 | −16.95 ± 1.55 |
| Polydispersity index Average ± SD | 0.225 ± 0.02 | 0.250 ± 0.01 |
| Diameter (nm) Average ± SD | 57.94± 0.85 | 110.54 ± 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fotoran, W.L.; Kleiber, N.; Glitz, C.; Wunderlich, G. A DNA Vaccine Encoding Plasmodium falciparum PfRH5 in Cationic Liposomes for Dermal Tattooing Immunization. Vaccines 2020, 8, 619. https://doi.org/10.3390/vaccines8040619
Fotoran WL, Kleiber N, Glitz C, Wunderlich G. A DNA Vaccine Encoding Plasmodium falciparum PfRH5 in Cationic Liposomes for Dermal Tattooing Immunization. Vaccines. 2020; 8(4):619. https://doi.org/10.3390/vaccines8040619
Chicago/Turabian StyleFotoran, Wesley Luzetti, Nicole Kleiber, Christiane Glitz, and Gerhard Wunderlich. 2020. "A DNA Vaccine Encoding Plasmodium falciparum PfRH5 in Cationic Liposomes for Dermal Tattooing Immunization" Vaccines 8, no. 4: 619. https://doi.org/10.3390/vaccines8040619
APA StyleFotoran, W. L., Kleiber, N., Glitz, C., & Wunderlich, G. (2020). A DNA Vaccine Encoding Plasmodium falciparum PfRH5 in Cationic Liposomes for Dermal Tattooing Immunization. Vaccines, 8(4), 619. https://doi.org/10.3390/vaccines8040619