Combined Exercise Training and l-Glutamine Supplementation Enhances Both Humoral and Cellular Immune Responses after Influenza Virus Vaccination in Elderly Subjects

 , , , , ,
, , , , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
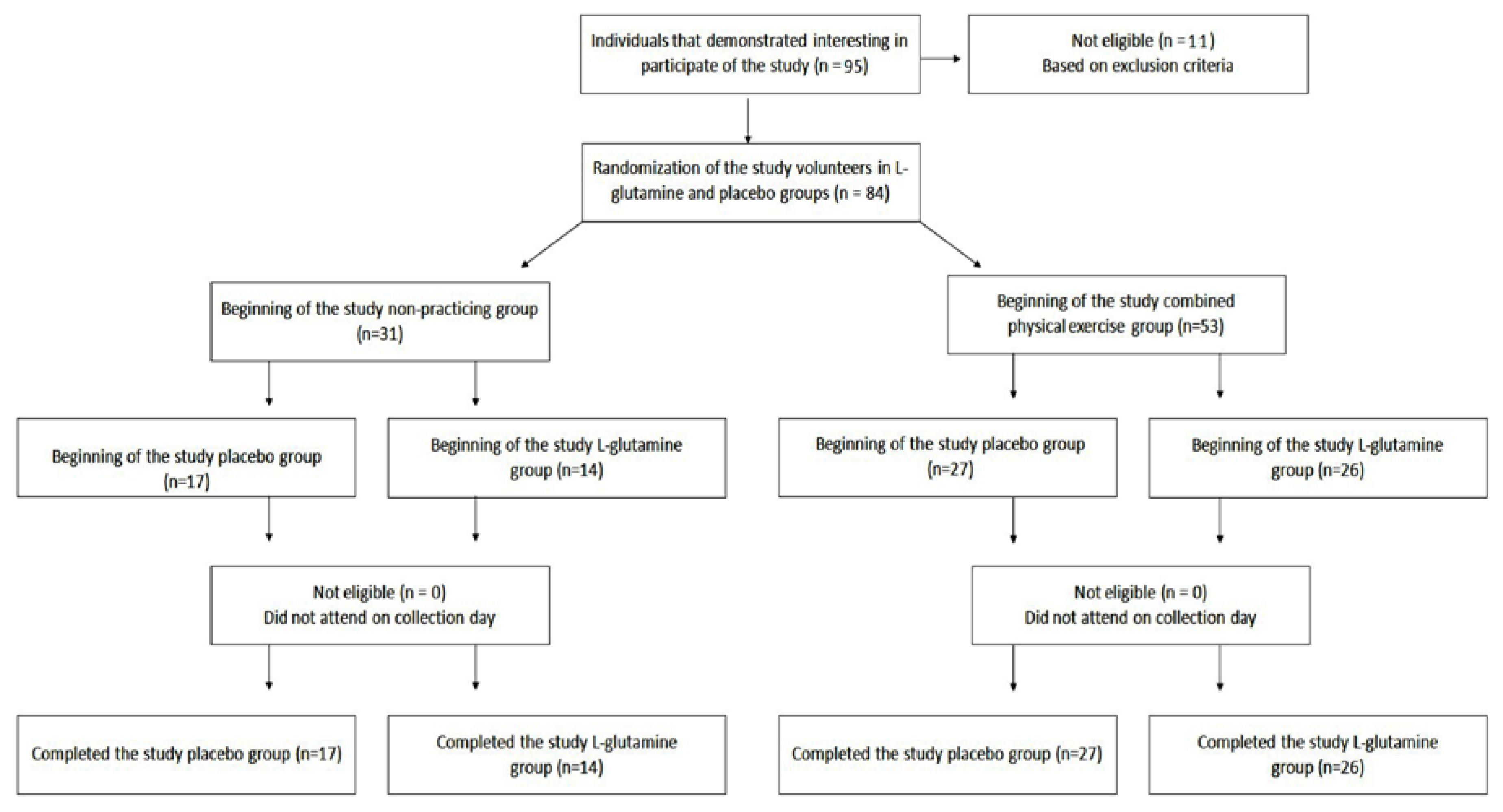
2.1. Subjects and Study Design
2.2. Combined-Exercise Training (CET)
2.3. Non-Practicing Group
2.4. l-Glutamine or Placebo Supplementation
2.5. Influenza Vaccine
2.6. Collection of the Samples
2.7. Immunophenotypic Profile
2.8. Determination of Specific Antibodies (IgA, IgM, IgG) for the Flu Vaccine (Influenza Virus)
2.9. Hemagglutination-Inhibition (HI) Assay
2.10. Statistical Analysis
3. Results
3.1. Combined Exercise Training and l-Glutamine Supplementation Improves the Specific Antibodies Response for Influenza Vaccine
3.2. l-Glutamine Supplementation Improved the Neutralizing Antibody Capacity
3.3. l-Glutamine Supplementation Increases the Absolute Number of Effector CD4+ T Cells, Whereas the CET Increase the Activated CD4+ T Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. World Report on Ageing and Health; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Sgarbieri, V.C.; Pacheco, M.T.B. Healthy human aging: Intrinsic and environmental factors. Braz. J. Food Technol. 2017, 20. [Google Scholar] [CrossRef] [Green Version]
- Müller, L.; Di Benedetto, S.; Pawelec, G. The immune system and its dysregulation with aging. In Biochemistry and Cell Biology of Ageing: Part II Clinical Science; Springer: Berlin/Heidelberg, Germany, 2019; pp. 21–43. [Google Scholar]
- Pera, A.; Campos, C.; López, N.; Hassouneh, F.; Alonso, C.; Tarazona, R.; Solana, R. Immunosenescence: Implications for response to infection and vaccination in older people. Maturitas 2015, 82, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Dugan, H.L.; Henry, C.; Wilson, P.C. Aging and influenza vaccine-induced immunity. Cell. Immunol. 2020, 348, 103998. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Weinberger, B. Vaccines to Prevent Infectious Diseases in the Older Population: Immunological Challenges and Future Perspectives. Front. Immunol. 2020, 11, 717. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.C.L.; Strickland, M.C.; Larbi, A. Changes in T Cell Homeostasis and Vaccine Responses in Old Age. Interdiscip Top. Gerontol. Geriatr. 2020, 43, 36–55. [Google Scholar] [PubMed]
- Oh, S.J.; Lee, J.K.; Shin, O.S. Aging and the Immune System: The Impact of Immunosenescence on Viral Infection, Immunity and Vaccine Immunogenicity. Immune Netw. 2019, 19, e37. [Google Scholar] [CrossRef] [PubMed]
- Bachi, A.L.; Suguri, V.M.; Ramos, L.R.; Mariano, M.; Vaisberg, M.; Lopes, J.D. Increased production of autoantibodies and specific antibodies in response to influenza virus vaccination in physically active older individuals. Results Immunol. 2013, 3, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, R.R. Protein supplements and exercise. Am. J. Clin. Nutr. 2000, 72 (Suppl. 2), 551S–557S. [Google Scholar] [CrossRef] [Green Version]
- Piattoly, T.; Parish, T.R.; Welsch, M.A. L-Glutamine Supplementation: Effects on endurance, power and recovery. Curr. Top. Nutraceutical Res. 2013, 11, 62–70. [Google Scholar]
- Legault, Z.; Bagnall, N.; Kimmerly, D.S. The Influence of Oral L-Glutamine Supplementation on Muscle Strength Recovery and Soreness Following Unilateral Knee Extension Eccentric Exercise. Int. J. Sport Nutr. Exerc. Metab. 2015, 25, 417–426. [Google Scholar] [CrossRef]
- Almeida, E.B.; Santo, J.D.M.B.D.; Paixão, V.; Amaral, J.B.; Foster, R.; Sperandio, A.; Roseira, T.; Rossi, M.; Cordeiro, T.G.; Monteiro, F.R.; et al. L-Glutamine Supplementation Improves the Benefits of Combined-Exercise Training on Oral Redox Balance and Inflammatory Status in Elderly Individuals. Oxid Med. Cell. Longev. 2020, 2020, 2852181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harriss, D.J.; Atkinson, G. Ethical Standards in Sport and Exercise Science Research: 2016 Update. Int. J. Sports Med. 2015, 36, 1121–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ACSM. ACSM’s Guidelines for Exercise Testing and Prescription; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013. [Google Scholar]
- Craig, C.L.; Marshall, A.L.; Sjöström, M.; E Bauman, A.; Booth, M.L.; E Ainsworth, B.; Pratt, M.; Ekelund, U.; Yngve, A.; Sallis, J.F.; et al. International physical activity questionnaire: 12-country reliability and validity. Med. Sci. Sports Exerc. 2003, 35, 1381–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedetti, T.R.B.; Antunes, P.d.C.; Rodriguez-Añez, C.R.; Mazo, G.Z.; Petroski, É.L. Reprodutibilidade e validade do Questionário Internacional de Atividade Física (IPAQ) em homens idosos. Rev. Bras. Med. Esporte 2007, 13, 11–16. [Google Scholar] [CrossRef]
- World Health Organization. Global Recommendations on Physical Activity for Health; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Fuentes, E.; Fuentes, M.; Alarcon, M.; Palomo, I. Immune system dysfunction in the elderly. An. Acad. Bras. Ciências 2017, 89, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Walsh, N.P.; Gleeson, M.; Shephard, R.J.; Gleeson, M.; Woods, J.A.; Bishop, N.C.; Fleshner, M.; Green, C.J.; Pedersen, B.K.; Hoffman-Goetz, L.; et al. Position statement. Part one: Immune function and exercise. Exerc. Immunol. Rev. 2011, 17, 6–63. [Google Scholar]
- Kohut, M.L.; Cooper, M.M.; Nickolaus, M.S.; Russell, D.R.; Cunnick, J.E. Exercise and psychosocial factors modulate immunity to influenza vaccine in elderly individuals. J. Gerontol. A Biol. Sci. Med. Sci. 2002, 57, M557–M562. [Google Scholar] [CrossRef] [Green Version]
- Long, J.; Ring, C.; Drayson, M.; Bosch, J.; Campbell, J.P.; Bhabra, J.; Browne, D.; Dawson, J.; Harding, S.; Lau, J.; et al. Vaccination response following aerobic exercise: Can a brisk walk enhance antibody response to pneumococcal and influenza vaccinations? Brain Behav. Immun. 2012, 26, 680–687. [Google Scholar] [CrossRef]
- Minuzzi, L.G.; Rama, L.; Chupel, M.U.; Rosado, F.; Dos Santos, J.V.; Simpson, R.; Martinho, A.; Paiva, A.; Teixeira, A.M. Effects of lifelong training on senescence and mobilization of T lymphocytes in response to acute exercise. Exerc. Immunol. Rev. 2018, 24, 72–84. [Google Scholar]
- Simpson, R.J.; Bosch, J.A. Special issue on exercise immunology: Current perspectives on aging, health and extreme performance. Brain Behav. Immun. 2014, 39, 1–7. [Google Scholar] [CrossRef]
- Silva, L.C.R.; De Araújo, A.L.; Fernandes, J.R.; Matias, M.D.S.T.; Silva, P.R.; Duarte, A.J.S.; Leme, L.E.G.; Benard, G. Moderate and intense exercise lifestyles attenuate the effects of aging on telomere length and the survival and composition of T cell subpopulations. Age 2016, 38, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.E. Is immunosenescence influenced by our lifetime “dose” of exercise? Biogerontology 2016, 17, 581–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, L.; Pawelec, G. Aging and immunity—Impact of behavioral intervention. Brain Behav. Immun. 2014, 39, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Stervbo, U.; Pohlmann, D.; Baron, U.; Bozzetti, C.; Jürchott, K.; Mälzer, J.N.; Nienen, M.; Olek, S.; Roch, T.; Schulz, A.R.; et al. Age dependent differences in the kinetics of γδ T cells after influenza vaccination. PLoS ONE 2017, 12, e0181161. [Google Scholar] [CrossRef]
- Carlock, M.A.; Ingram, J.G.; Clutter, E.F.; Cecil, N.C.; Ramgopal, M.; Zimmerman, R.K.; Warren, W.; Kleanthous, H.; Ross, T.M. Impact of age and pre-existing immunity on the induction of human antibody responses against influenza B viruses. Hum. Vaccin. Immunother. 2019, 15, 2030–2043. [Google Scholar] [CrossRef] [Green Version]
- Cetinbas, F.; Yelken, B.; Gulbas, Z. Role of glutamine administration on cellular immunity after total parenteral nutrition enriched with glutamine in patients with systemic inflammatory response syndrome. J. Crit. Care 2010, 25, 661.e1–661.e6. [Google Scholar] [CrossRef]
- Singh, N.; Mishra, S.K.; Sachdev, V.; Sharma, H.; Upadhyay, A.D.; Arora, I.; Saraya, A. Effect of oral glutamine supplementation on gut permeability and endotoxemia in patients with severe acute pancreatitis: A randomized controlled trial. Pancreas 2014, 43, 867–873. [Google Scholar] [CrossRef]
- Shariatpanahi, Z.V.; Eslamian, G.; Ardehali, S.H.; Baghestani, A.R. Effects of Early Enteral Glutamine Supplementation on Intestinal Permeability in Critically Ill Patients. Indian J. Crit. Care Med. 2019, 23, 356–362. [Google Scholar] [CrossRef]
- Klentrou, P.; Cieslak, T.; MacNeil, M.; Vintinner, A.; Plyley, M. Effect of moderate exercise on salivary immunoglobulin A and infection risk in humans. Eur. J. Appl. Physiol. 2002, 87, 153–158. [Google Scholar] [CrossRef]
- Shimizu, K.; Kimura, F.; Akimoto, T.; Akama, T.; Otsuki, T.; Nishijima, T.; Kuno, S.; Kono, I. Effects of exercise, age and gender on salivary secretory immunoglobulin A in elderly individuals. Exerc. Immunol. Rev. 2007, 13, 55–66. [Google Scholar]
- Beyer, W.E.; Van der Logt, J.T.; van Beek, R.; Masurel, N. Immunoglobulin G, A and M response to influenza vaccination in different age groups: Effects of priming and boosting. J. Hyg. 1986, 96, 513–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, J.W.; Crowe, M.; Blank, S.E. Chronic glutamine supplementation increases nasal but not salivary IgA during 9 days of interval training. J. Appl. Physiol. 2004, 97, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.-H.; Xu, R.-M.; Zhang, Q.-H.; Shen, G.-Q.; Ma, M.; Zhao, X.-P.; Guo-Qing, S.; Wang, Y. Glutamine supplementation and immune function during heavy load training. Int. J. Clin. Pharmacol. Ther. 2015, 53, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.; MacDonald, T.; Blumberg, R. Principles of Mucosal Immunology; Garland Science: New York, NY, USA, 2012. [Google Scholar]
- Loeb, N.; Andrew, M.K.; Loeb, M.; A Kuchel, G.; Haynes, L.; McElhaney, J.E.; Verschoor, C.P.; A George, K. Frailty Is Associated with Increased Hemagglutination-Inhibition Titers in a 4-Year Randomized Trial Comparing Standard- and High-Dose Influenza Vaccination. Open Forum Infect. Dis. 2020, 7, ofaa148. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Cell | Profile | CD Markers | |
|---|---|---|---|
| CD4+ T cells | Activated | CD3+ | CD4+CD38+HLA-DR+ |
| Naïve | CD3+ | CD4+CD27+CD45RA+CD197+ | |
| Effector | CD3+ | CD4+CD27-CD45RA+CD197- | |
| Groups Characteristics | Volunteers (n = 84) | ||||
|---|---|---|---|---|---|
| Non-Practitioners (NP, n = 31) | Combined-Exercise Training (CET, n = 53) | p Value | |||
| Placebo (n = 17) | l-Glutamine (n = 14) | Placebo (n = 27) | l-Glutamine (n = 26) | ||
| Age (year) | 75.1 ± 7.1 | 72.9 ± 5.4 | 72.2 ± 5.9 | 71.2 ± 5.9 | >0.05 |
| Height (m) | 154.2 ± 9.5 | 162.3 ± 8.3 * | 156 ± 9.7 | 156.8 ± 8.8 | <0.05 |
| Weight (kg) | 66.0 ± 10.7 | 75.7 ± 14.5 * | 61.9 ± 10.3 | 62.9 ± 13.0 | <0.05 |
| Body mass index (kg/m²) | 27.7 ± 4.0 # | 28.5 ± 3.7 $ | 25.4 ± 3.6 | 25.4 ± 3.9 | <0.05 |
| Total body fat (%) | 39.6 ± 10.1 | 38.1 ± 9.1 | 35.4 ± 7.7 | 35.3 ± 7.7 | >0.05 |
| Fat-free mass (%) | 60.4 ± 9.3 | 61.9 ± 9.2 | 64.6 ± 7.8 | 64.7 ± 7.5 | >0.05 |
| Skeletal muscle mass (kg) | 18.3 ± 4.1 | 22.3 ± 4.0 | 19.6 ± 3.6 | 19.8 ± 3.9 | >0.05 |
| IPAQ | |||||
| Physical activity (min/week) | 433.7 ± 76.1 # | 371.4 ± 69.2 $ | 677.3 ± 60.4 | 754.1 ± 85.6 | <0.05 |
| Sitting (min/week) | 1685 ± 176.5 # | 1874 ± 178.6 $ | 1224 ± 112.21 | 1326 ± 107.74 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, F.R.; Roseira, T.; Amaral, J.B.; Paixão, V.; Almeida, E.B.; Foster, R.; Sperandio, A.; Rossi, M.; Amirato, G.R.; Apostólico, J.S.; et al. Combined Exercise Training and l-Glutamine Supplementation Enhances Both Humoral and Cellular Immune Responses after Influenza Virus Vaccination in Elderly Subjects. Vaccines 2020, 8, 685. https://doi.org/10.3390/vaccines8040685
Monteiro FR, Roseira T, Amaral JB, Paixão V, Almeida EB, Foster R, Sperandio A, Rossi M, Amirato GR, Apostólico JS, et al. Combined Exercise Training and l-Glutamine Supplementation Enhances Both Humoral and Cellular Immune Responses after Influenza Virus Vaccination in Elderly Subjects. Vaccines. 2020; 8(4):685. https://doi.org/10.3390/vaccines8040685
Chicago/Turabian StyleMonteiro, Fernanda R., Tamaris Roseira, Jonatas B. Amaral, Vitória Paixão, Ewin B. Almeida, Roberta Foster, Adriane Sperandio, Marcelo Rossi, Gislene R. Amirato, Juliana S. Apostólico, and et al. 2020. "Combined Exercise Training and l-Glutamine Supplementation Enhances Both Humoral and Cellular Immune Responses after Influenza Virus Vaccination in Elderly Subjects" Vaccines 8, no. 4: 685. https://doi.org/10.3390/vaccines8040685
APA StyleMonteiro, F. R., Roseira, T., Amaral, J. B., Paixão, V., Almeida, E. B., Foster, R., Sperandio, A., Rossi, M., Amirato, G. R., Apostólico, J. S., Santos, C. A. F., Felismino, E. S., Leal, F. B., Thomazelli, L. M., Durigon, E. L., Oliveira, D. B. L., Vieira, R. P., Santos, J. M. B., & Bachi, A. L. L. (2020). Combined Exercise Training and l-Glutamine Supplementation Enhances Both Humoral and Cellular Immune Responses after Influenza Virus Vaccination in Elderly Subjects. Vaccines, 8(4), 685. https://doi.org/10.3390/vaccines8040685