Changes in Serum Biomarkers of Oxidative Stress in Cattle Vaccinated with Tick Recombinant Antigens: A Pilot Study

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Production of Recombinant Subolesin Antigens and Vaccine Formulation
2.3. Determination of Serum Antibody Levels by ELISA
2.4. Antioxidant Biomarkers
2.5. Oxidative Biomarkers
2.6. Statistical Analysis
3. Results
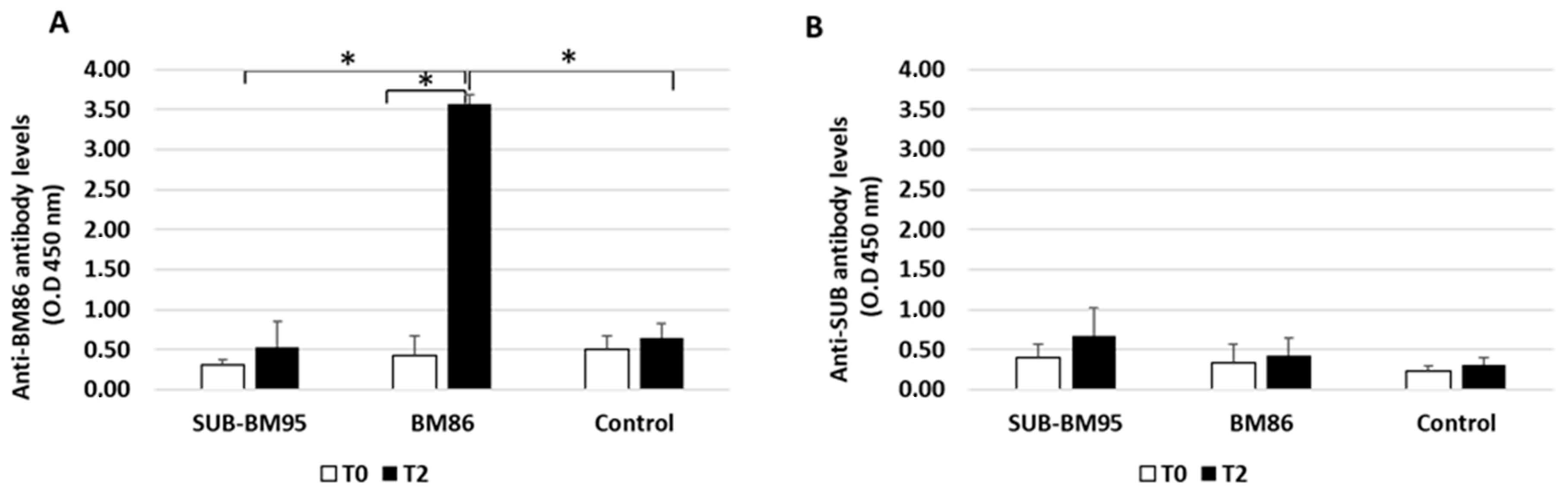
3.1. Characterization of the Antibody Response in Vaccinated Cattle
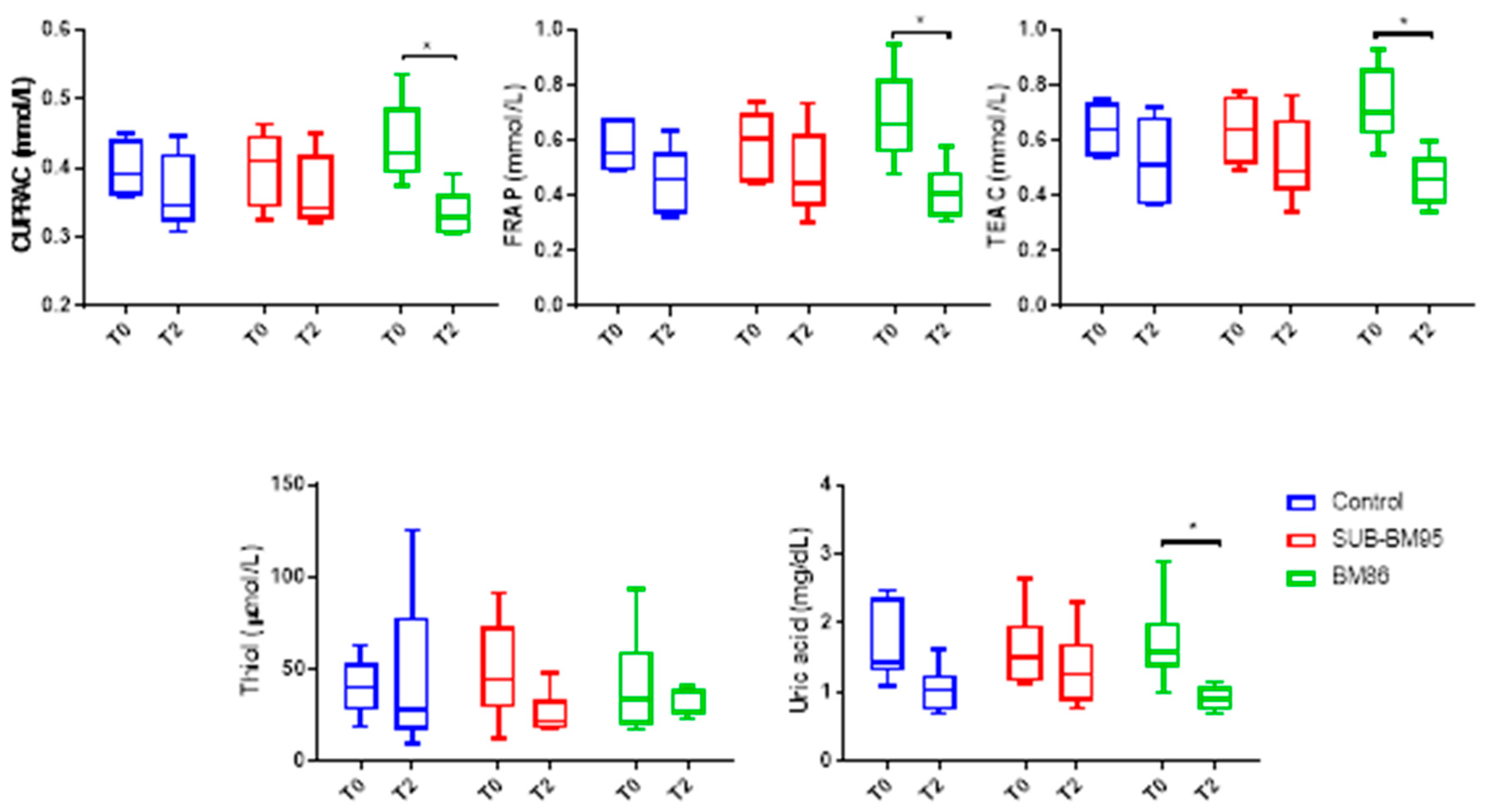
3.2. Changes in Antioxidant Biomarkers
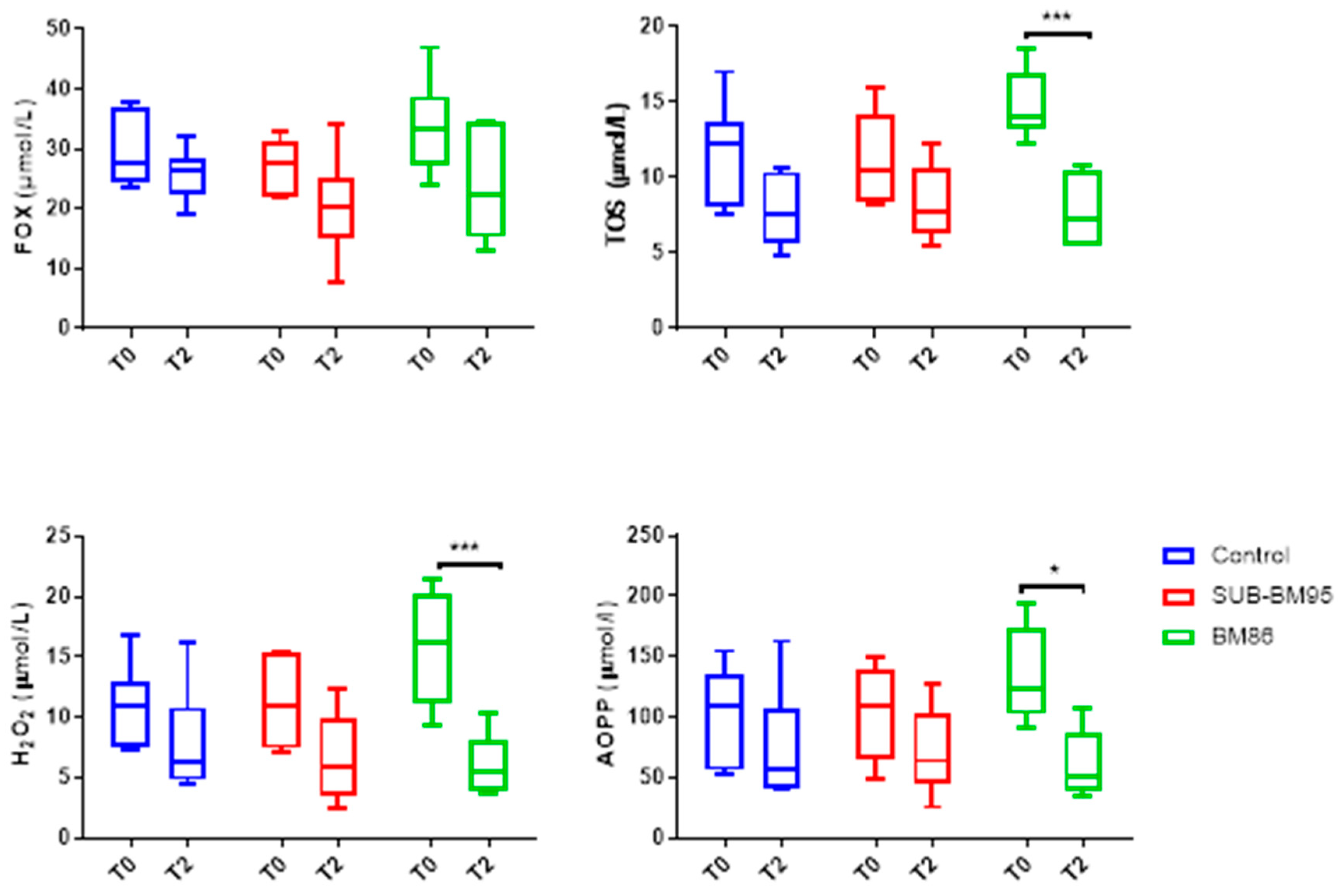
3.3. Changes in Oxidant Biomarkers
3.4. Correlation Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Camicas, J.L.; Hervy, J.P.; Adam, F.; Morel, P.C. The Ticks of the World (Acarida, Ixodida); Orstom Editions: Paris, France, 1998. [Google Scholar]
- Boulanger, N.; Boyer, P.; Talagrand-Reboul, E.; Hansmann, Y. Ticks and tick-borne diseases. Med. Mal. Infect. 2019, 49, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Peter, R.; Van den Bossche, P.; Penzhorn, B.L.; Sharp, B. Tick, fly, and mosquito control—Lessons from the past, solutions for the future. Veter Parasitol. 2005, 132, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T.; Cadar, D.; Spînu, M. Tick prevention at a crossroad: New and renewed solutions. Veter Parasitol. 2012, 187, 357–366. [Google Scholar] [CrossRef]
- Karasuyama, H.; Miyake, K.; Yoshikawa, S. Immunobiology of Acquired Resistance to Ticks. Front. Immunol. 2020, 11, 601504. [Google Scholar] [CrossRef]
- Nuttall, P.A.; Trimnell, A.R.; Kazimirova, M.; Labuda, M. Exposed and concealed antigens as vaccine targets for controlling ticks and tick-borne diseases. Parasite Immunol. 2006, 28, 155–163. [Google Scholar] [CrossRef]
- Tabor, A.E.; Valle, M.R. A review of reverse vaccinology approaches for the development of vaccines against ticks and tick borne diseases. Ticks Tick-Borne Dis. 2016, 7, 573–585. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, J. Controlling ticks and tick-borne diseases looking forward. Ticks Tick-Borne Dis. 2018, 9, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Valle, M.R.; Guerrero, F.D. Anti-tick vaccines in the omics era. Front. Biosci. 2018, 10, 122–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Cid, J.A.; Jiménez, M.; Cornelie, S.; Molina, R.; Alarcón, P.; Lacroix, M.-N.; Pinal, R.; Delacour, S.; Lucientes, J.; Canales, M.; et al. Characterization of Aedes albopictus akirin for the control of mosquito and sand fly infestations. Vaccine 2011, 29, 77–82. [Google Scholar] [CrossRef]
- Liljeroos, L.; Malito, E.; Ferlenghi, I.; Bottomley, M.J. Structural and Computational Biology in the Design of Immunogenic Vaccine Antigens. J. Immunol. Res. 2015, 2015, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ndawula, C., Jr.; Tabor, A.E. Cocktail Anti-Tick Vaccines: The Unforeseen Constraints and Approaches toward Enhanced Efficacies. Vaccines 2020, 19, 457. [Google Scholar] [CrossRef] [PubMed]
- Achkar, J.M.; Prados-Rosales, R. Updates on antibody functions in Mycobacterium tuberculosis infection and their relevance for developing a vaccine against tuberculosis. Curr. Opin. Immunol. 2018, 53, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Gazzinelli-Guimarães, A.C.; Gazzinelli-Guimarães, P.H.; Nogueira, D.S.; Oliveira, F.M.S.; Barbosa, F.S.; Amorim, C.C.O.; Cardoso, M.S.; Kraemer, L.; Caliari, M.V.; Akamatsu, M.A.; et al. IgG Induced by Vaccination With Ascaris suum Extracts Is Protective Against Infection. Front. Immunol. 2018, 9, 2535. [Google Scholar] [CrossRef]
- Schaefer, J.V.; Sedlák, E.; Kast, F.; Nemergut, M.; Plückthun, A. Modification of the kinetic stability of immunoglobulin G by solvent additives. mAbs 2018, 10, 607–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brader, M.L.; Estey, T.; Bai, S.; Alston, R.W.; Lucas, K.K.; Lantz, S.; Landsman, P.; Maloney, K.M. Examination of Thermal Unfolding and Aggregation Profiles of a Series of Developable Therapeutic Monoclonal Antibodies. Mol. Pharm. 2015, 12, 1005–1017. [Google Scholar] [CrossRef]
- Margiloff, L.; Chaplia, L.; Chow, A.; Singhal, P.C.; Mattana, J. Metal-catalyzed oxidation of immunoglobulin G impairs Fc receptor-mediated binding to macrophages. Free. Radic. Biol. Med. 1998, 25, 780–785. [Google Scholar] [CrossRef]
- Rezaie, A.; Parker, R.D.; Abdollahi, M. Oxidative stress and pathogenesis of inflammatory bowel disease: An epiphenomenon or the cause? Dig. Dis. Sci. 2007, 52, 2015–2021. [Google Scholar] [CrossRef]
- Han, S.; Wang, G.; Xu, N.; Liu, H. Quantitative Assessment of the Effects of Oxidants on Antigen-Antibody Binding In Vitro. Oxidative Med. Cell. Longev. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Campos, C.; Guzmán, R.; López-Fernández, E.; Casado, Á. Evaluation of the copper (II) reduction assay using bathocuproinedisulfonic acid disodium salt for the total antioxidant capacity assessment: The CUPRAC–BCS assay. Anal. Biochem. 2009, 392, 37–44. [Google Scholar] [CrossRef]
- Bigagli, E.; Lodovici, M. Circulating Oxidative Stress Biomarkers in Clinical Studies on Type 2 Diabetes and Its Complications. Oxidative Med. Cell. Longev. 2019, 2019, 1–17. [Google Scholar] [CrossRef]
- Bottari, N.B.; Crivellenti, L.Z.; Borin-Crivellenti, S.; Oliveira, J.R.; Coelho, S.B.; Contin, C.M.; Tatsch, E.; Moresco, R.N.; Santana, A.E.; Tonin, A.A.; et al. Iron metabolism and oxidative profile of dogs naturally infected by Ehrlichia canis: Acute and subclinical disease. Microb. Pathog. 2016, 92, 26–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.G.; Chang, T.-S.; Jeong, W.; Kang, D. Methods for detection and measurement of hydrogen peroxide inside and outside of cells. Mol. Cells 2010, 29, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Almazán, C.; Moreno-Cantú, O.; Moreno-Cid, J.A.; Galindo, R.C.; Canales, M.; Villar, M.; De La Fuente, J. Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens. Vaccine 2012, 30, 265–272. [Google Scholar] [CrossRef]
- Canales, M.; Almazán, C.; De La Lastra, J.M.P.; De La Fuente, J.; De La Lastra, J.M.P. Anaplasma marginale major surface protein 1a directs cell surface display of tick BM95 immunogenic peptides on Escherichia coli. J. Biotechnol. 2008, 135, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Canales, M.; Almazán, C.; Naranjo, M.V.; Jongejan, F.; De La Fuente, J. Vaccination with recombinant Boophilus annulatus Bm86 ortholog protein, Ba86, protects cattle against B. annulatus and B. microplus infestations. BMC Biotechnol. 2009, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Willadsen, P.; Smith, D.; Cobon, G.; McKenna, R.V. Comparative vaccination of cattle against Boophilus microplus with recombinant antigen Bm86 alone or in combination with recombinant Bm. Parasite Immunol. 1996, 18, 241–246. [Google Scholar] [CrossRef]
- Kasaija, P.D.; Contreras, M.; Kabi, F.; Mugerwa, S.; De La Fuente, J. Vaccination with Recombinant Subolesin Antigens Provides Cross-Tick Species Protection in Bos indicus and Crossbred Cattle in Uganda. Vaccines 2020, 8, 319. [Google Scholar] [CrossRef]
- Rubio, C.P.; Tvarijonaviciute, A.; Caldin, M.; Hernández-Ruiz, J.; Cerón, J.J.; Martínez-Subiela, S.; Tecles, F. Stability of biomarkers of oxidative stress in canine serum. Res. Veter. Sci. 2018, 121, 85–93. [Google Scholar] [CrossRef]
- Rubio, C.P.; Martínez-Subiela, S.; Tvarijonaviciute, A.; Hernández-Ruiz, J.; Pardo-Marin, L.; Segarra, S.; Ceron, J.J. Changes in serum biomarkers of oxidative stress after treatment for canine leishmaniosis in sick dogs. Comp. Immunol. Microbiol. Infect. Dis. 2016, 49, 51–57. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E.; Altun, M. Total antioxidant capacity assay of human serum using copper (II)-neocuproine as chromogenic oxidant: The CUPRAC method. Free. Radic. Res. 2005, 39, 949–961. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jocelyn, P.C. Spectrophotometric assay of thiols. Methods Enzymol. 1987, 143, 44–67. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, C.M.; Dos Santos, R.C.C.; Lima, E.S. A simple automated procedure for thiol measurement in human serum samples. J. Bras. Patol. Med. Lab. 2006, 42, 345–350. [Google Scholar] [CrossRef]
- Fossati, P.; Prencipe, L.; Berti, G. Use of 3,5-dichloro-2-hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzymic assay of uric acid in serum and urine. Clin. Chem. 1980, 26, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Arab, K.; Steghens, J.-P. Plasma lipid hydroperoxides measurement by an automated xylenol orange method. Anal. Biochem. 2004, 325, 158–163. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [Green Version]
- Tatzber, F.; Griebenow, S.; Wonisch, W.; Winkler, R. Dual method for the determination of peroxidase activity and total peroxides-iodide leads to a significant increase of peroxidase activity in human sera. Anal. Biochem. 2003, 316, 147–153. [Google Scholar] [CrossRef]
- Kumar, R. Molecular markers and their application in the monitoring of acaricide resistance in Rhipicephalus microplus. Exp. Appl. Acarol. 2019, 78, 149–172. [Google Scholar] [CrossRef]
- Artigas-Jerónimo, S.; Villar, M.; Cabezas-Cruz, A.; Valdés, J.J.; Estrada-Peña, A.; Alberdi, P.; De La Fuente, J. Functional Evolution of Subolesin/Akirin. Front. Physiol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Elias, S.C.; Collins, K.A.; Halstead, F.D.; Choudhary, P.; Bliss, C.M.; Ewer, K.J.; Sheehy, S.H.; Duncan, C.J.A.; Biswas, S.; Hill, A.V.S.; et al. Assessment of Immune Interference, Antagonism, and Diversion following Human Immunization with Biallelic Blood-Stage Malaria Viral-Vectored Vaccines and Controlled Malaria Infection. J. Immunol. 2013, 190, 1135–1147. [Google Scholar] [CrossRef]
- De La Fuente, J.; Rodríguez, M.; Montero, C.V.; Redondo, M.L.; García-García, J.C.; Méndez, L.; Serrano, E.; Valdes, M.; Enríquez, A.; Canales, M.; et al. Vaccination against ticks (Boophilus spp.): The experience with the Bm86-based vaccine Gavac™. Genet. Anal. Biomol. Eng. 1999, 15, 143–148. [Google Scholar] [CrossRef]
- Rodríguez-Valle, M.; Montero, C.; Machado, H.; Joglar, M.; de la Fuente, J.; Garcia-Garcia, J.C. The evaluation of yeast derivatives as adjuvants for the immune response to the Bm86 antigen in cattle. BMC Biotechnol. 2001, 1, 2. [Google Scholar] [CrossRef]
- Yardim-Akaydin, S.; Ozkan, Y.; Ozkan, E.; Torun, M.; Simşek, B. The role of plasma thiol compounds and antioxidant vitamins in patients with cardiovascular diseases. Clin. Chim. Acta 2003, 338, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, D.A.; Forman, H.J. Cellular glutathione and thiols metabolism. Biochem. Pharmacol. 2002, 64, 1019–1026. [Google Scholar] [CrossRef]
- Gao, M.; Li, Y.; Zheng, A.; Xue, X.; Chen, L.; Kong, Y. Lymphocyte Oxidative Stress/Genotoxic Effects Are Related to Serum IgG and IgA Levels in Coke Oven Workers. Sci. World J. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, S.A.; Bustnes, J.O.; Schnug, L.; Bourgeon, S.; Johnsen, T.V.; Ballesteros, M.; Sonne, C.; Herzke, D.; Eulaers, I.; Jaspers, V.L.B.; et al. Antiparasite treatments reduce humoral immunity and impact oxidative status in raptor nestlings. Ecol. Evol. 2013, 3, 5157–5166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Fuente, J.; Kocan, K.M. Development of Vaccines for Control of Tick Infestations and Interruption of Pathogen Transmission. In Biology of Ticks, 2nd ed.; Sonenshine, D., Roe, M., Eds.; Chapter Oxford University Press: New York, NY, USA, 2013; pp. 333–352. [Google Scholar]
- Shokri, M.; Roohvand, F.; Alimohammadian, M.H.; Ebrahimi-Rad, M.; Ajdary, S. Comparing Montanide ISA 720 and 50-V2 adjuvants formulated with LmSTI1 protein of Leishmania major indicated the potential cytokine patterns for induction of protective immune responses in BALB/c mice. Mol. Immunol. 2016, 76, 108–115. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Heading | CUPRAC | FRAP | TEAC | Thiol | Uric Acid | FOX | TOS | H2O2 | AOPP |
|---|---|---|---|---|---|---|---|---|---|
| Spearman r | −0.257 | −0.222 | −0.215 | 0.087 | −0.198 | 0.202 | −0.418 | −0.214 | −0.209 |
| p value | 0.303 | 0.376 | 0.390 | 0.729 | 0.430 | 0.421 | 0.084 | 0.393 | 0.404 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras, M.; Peres Rubio, C.; de la Fuente, J.; Villar, M.; Merino, O.; Mosqueda, J.; Cerón, J.J. Changes in Serum Biomarkers of Oxidative Stress in Cattle Vaccinated with Tick Recombinant Antigens: A Pilot Study. Vaccines 2021, 9, 5. https://doi.org/10.3390/vaccines9010005
Contreras M, Peres Rubio C, de la Fuente J, Villar M, Merino O, Mosqueda J, Cerón JJ. Changes in Serum Biomarkers of Oxidative Stress in Cattle Vaccinated with Tick Recombinant Antigens: A Pilot Study. Vaccines. 2021; 9(1):5. https://doi.org/10.3390/vaccines9010005
Chicago/Turabian StyleContreras, Marinela, Camila Peres Rubio, José de la Fuente, Margarita Villar, Octavio Merino, Juan Mosqueda, and José Joaquín Cerón. 2021. "Changes in Serum Biomarkers of Oxidative Stress in Cattle Vaccinated with Tick Recombinant Antigens: A Pilot Study" Vaccines 9, no. 1: 5. https://doi.org/10.3390/vaccines9010005