Evaluation of the Efficacy of a Cholera Toxin-Based Staphylococcus aureus Vaccine against Bovine Intramammary Challenge


 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, and Growth Conditions
2.2. Protein Expression and Purification
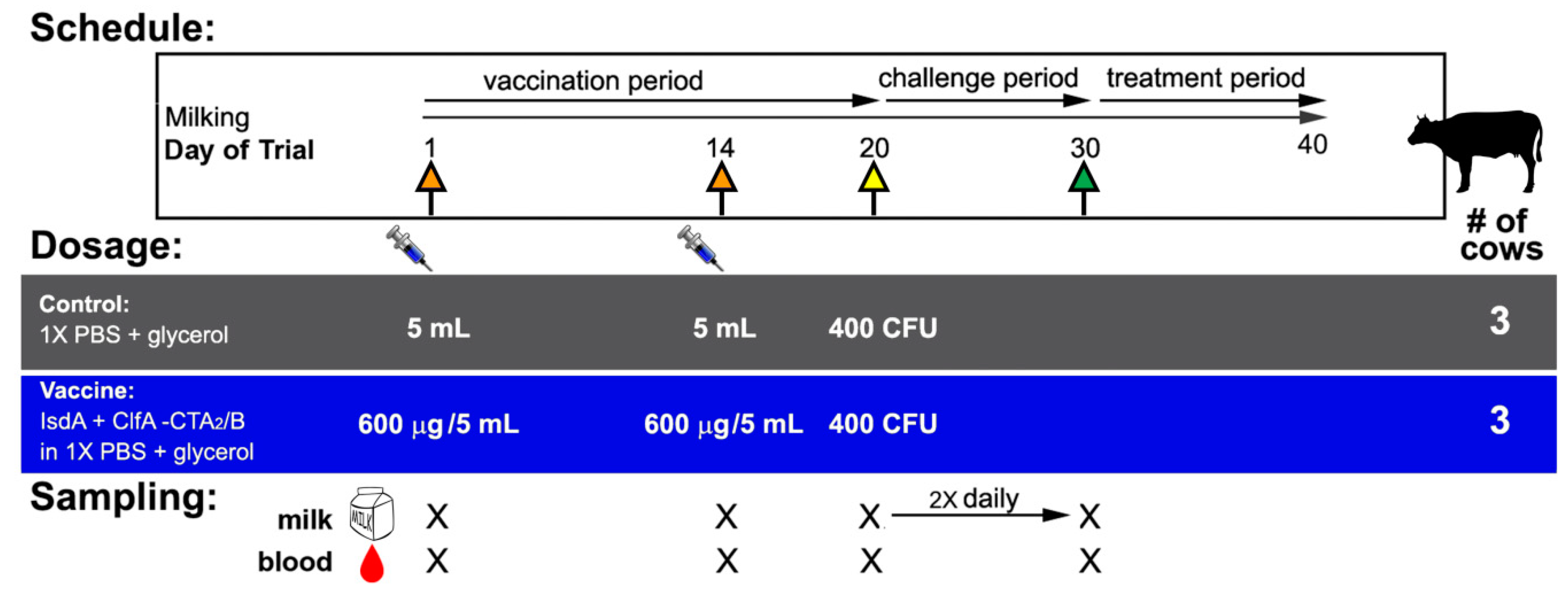
2.3. Animals, Vaccination, Challenge, and Clinical Assessment
2.4. Sample Collection and Milk Culture
2.5. IgG, IgG1, IgG2, and IgA Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Peripheral Blood Mononuclear Cell (PBMC) Isolation and Cytokine qPCR
2.7. Sample Size, Statistical Methods, and Analysis
3. Results
3.1. Bacterial Culture and Clinical Assessment
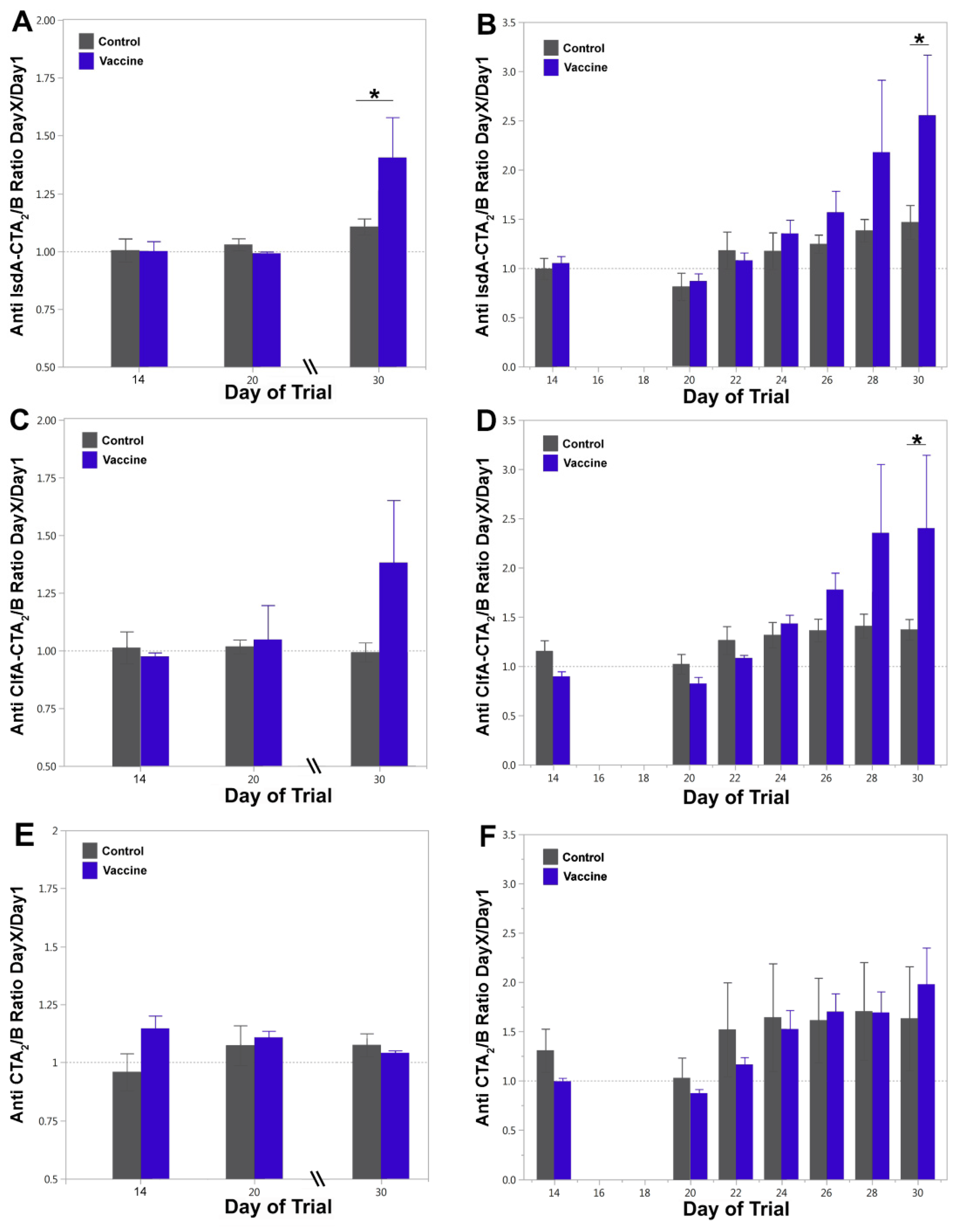
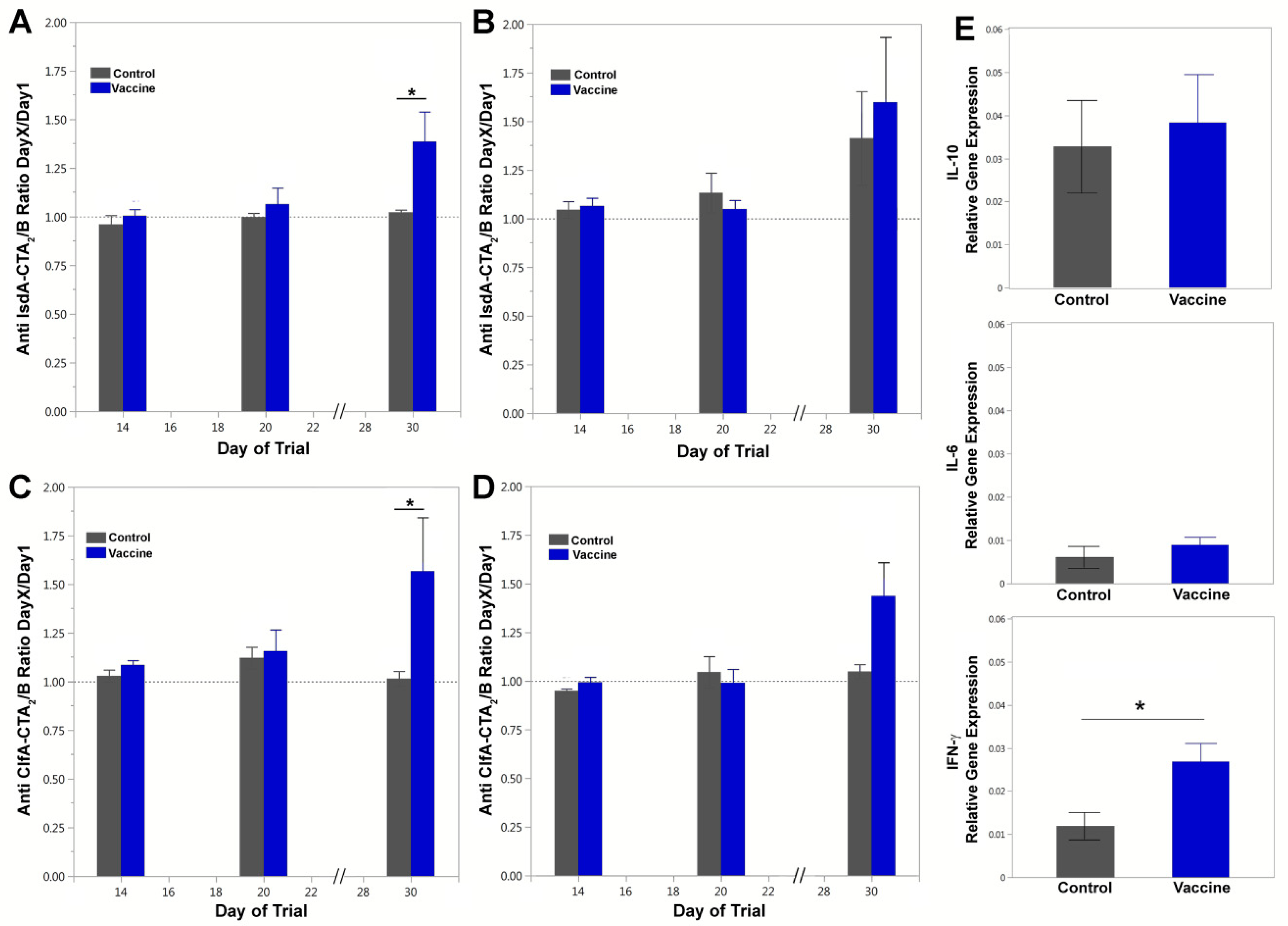
3.2. Vaccine-Specific Antibody Responses in Blood and Milk
3.3. Cytokine Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heikkila, A.M.; Liski, E.; Pyorala, S.; Taponen, S. Pathogen-specific production losses in bovine mastitis. J. Dairy Sci 2018, 101, 9493–9504. [Google Scholar] [CrossRef] [Green Version]
- APHIS, U. Dairy 2014. Milk Quality, Milking Procedures, and Mastitis on U.S. Dairies; United States Department of Agriculture: Washington, DC, USA, 2014; Volume 2016. [Google Scholar]
- Roberson, J.R.; Fox, L.K.; Hancock, D.D.; Gay, J.M.; Besser, T.E. Sources of intramammary infections from Staphylococcus aureus in dairy heifers at first parturition. J. Dairy Sci. 1998, 81, 687–693. [Google Scholar] [CrossRef]
- Hebert, A.; Sayasith, K.; Senechal, S.; Dubreuil, P.; Lagace, J. Demonstration of intracellular Staphylococcus aureus in bovine mastitis alveolar cells and macrophages isolated from naturally infected cow milk. FEMS Microbiol. Lett. 2000, 193, 57–62. [Google Scholar] [CrossRef]
- Sacco, S.C.; Velazquez, N.S.; Renna, M.S.; Beccaria, C.; Baravalle, C.; Pereyra, E.A.L.; Monecke, S.; Calvinho, L.F.; Dallard, B.E. Capacity of two Staphylococcus aureus strains with different adaptation genotypes to persist and induce damage in bovine mammary epithelial cells and to activate macrophages. Microb. Pathog. 2020, 142, 104017. [Google Scholar] [CrossRef] [PubMed]
- Zaatout, N.; Ayachi, A.; Kecha, M. Staphylococcus aureus persistence properties associated with bovine mastitis and alternative therapeutic modalities. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Sol, J.; Sampimon, O.C.; Snoep, J.J.; Schukken, Y.H. Factors associated with bacteriological cure during lactation after therapy for subclinical mastitis caused by Staphylococcus aureus. J. Dairy Sci. 1997, 80, 2803–2808. [Google Scholar] [CrossRef]
- Rollin, E.; Dhuyvetter, K.C.; Overton, M.W. The cost of clinical mastitis in the first 30 days of lactation: An economic modeling tool. Prev. Vet. Med. 2015, 122, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Cha, E.; Bar, D.; Hertl, J.A.; Tauer, L.W.; Bennett, G.; Gonzalez, R.N.; Schukken, Y.H.; Welcome, F.L.; Grohn, Y.T. The cost and management of different types of clinical mastitis in dairy cows estimated by dynamic programming. J. Dairy Sci. 2011, 94, 4476–4487. [Google Scholar] [CrossRef] [Green Version]
- Middleton, J.R.; Ma, J.; Rinehart, C.L.; Taylor, V.N.; Luby, C.D.; Steevens, B.J. Efficacy of different Lysigin formulations in the prevention of Staphylococcus aureus intramammary infection in dairy heifers. J. Dairy Res. 2006, 73, 10–19. [Google Scholar] [CrossRef]
- Schukken, Y.H.; Bronzo, V.; Locatelli, C.; Pollera, C.; Rota, N.; Casula, A.; Testa, F.; Scaccabarozzi, L.; March, R.; Zalduendo, D.; et al. Efficacy of vaccination on Staphylococcus aureus and coagulase-negative staphylococci intramammary infection dynamics in 2 dairy herds. J. Dairy Sci. 2014, 97, 5250–5264. [Google Scholar] [CrossRef] [Green Version]
- Bradley, A.J.; Breen, J.E.; Payne, B.; White, V.; Green, M.J. An investigation of the efficacy of a polyvalent mastitis vaccine using different vaccination regimens under field conditions in the United Kingdom. J. Dairy Sci. 2015, 98, 1706–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piepers, S.; Prenafeta, A.; Verbeke, J.; De Visscher, A.; March, R.; De Vliegher, S. Immune response after an experimental intramammary challenge with killed Staphylococcus aureus in cows and heifers vaccinated and not vaccinated with Startvac, a polyvalent mastitis vaccine. J. Dairy Sci. 2017, 100, 769–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landin, H.; Mork, M.J.; Larsson, M.; Waller, K.P. Vaccination against Staphylococcus aureus mastitis in two Swedish dairy herds. Acta Vet. Scand. 2015, 57, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freick, M.; Frank, Y.; Steinert, K.; Hamedy, A.; Passarge, O.; Sobiraj, A. Mastitis vaccination using a commercial polyvalent vaccine or a herd-specific Staphylococcus aureus vaccine. Results of a controlled field trial on a dairy farm. Tierarztl Prax Ausg G Grosstiere Nutztiere 2016, 44, 219–229. [Google Scholar] [CrossRef]
- Kim, H.K.; DeDent, A.; Cheng, A.G.; McAdow, M.; Bagnoli, F.; Missiakas, D.M.; Schneewind, O. IsdA and IsdB antibodies protect mice against Staphylococcus aureus abscess formation and lethal challenge. Vaccine 2010, 28, 6382–6392. [Google Scholar] [CrossRef] [Green Version]
- Clarke, S.R.; Andre, G.; Walsh, E.J.; Dufrene, Y.F.; Foster, T.J.; Foster, S.J. Iron-regulated surface determinant protein A mediates adhesion of Staphylococcus aureus to human corneocyte envelope proteins. Infect. Immun. 2009, 77, 2408–2416. [Google Scholar] [CrossRef] [Green Version]
- Stapleton, M.; Wright, L.; Clarke, S.; Moseby, H.; Tarkowski, A.; Vendrengh, M.; Foster, S. Identification of Conserved Antigens from Staphylococcal and Streptococcal Pathogens. J. Med. Microbiol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Clarke, S.R.; Brummell, K.J.; Horsburgh, M.J.; McDowell, P.W.; Mohamad, S.A.; Stapleton, M.R.; Acevedo, J.; Read, R.C.; Day, N.P.; Peacock, S.J.; et al. Identification of in vivo-expressed antigens of Staphylococcus aureus and their use in vaccinations for protection against nasal carriage. J. Infect. Dis. 2006, 193, 1098–1108. [Google Scholar] [CrossRef] [Green Version]
- Wolf, C.; Kusch, H.; Monecke, S.; Albrecht, D.; Holtfreter, S.; von Eiff, C.; Petzl, W.; Rainard, P.; Broker, B.M.; Engelmann, S. Genomic and proteomic characterization of Staphylococcus aureus mastitis isolates of bovine origin. Proteomics 2011, 11, 2491–2502. [Google Scholar] [CrossRef]
- Herron-Olson, L.; Fitzgerald, J.R.; Musser, J.M.; Kapur, V. Molecular Correlates of Host Specialization in Staphylococcus aureus. PLoS ONE 2007, 2, e1120. [Google Scholar] [CrossRef]
- Bouchard, D.; Peton, V.; Almeida, S.; Le Maréchal, C.; Miyoshi, A.; Azevedo, V.; Berkova, N.; Rault, L.; François, P.; Schrenzel, J.; et al. Genome sequence of Staphylococcus aureus Newbould 305, a strain associated with mild bovine mastitis. J. Bacteriol. 2012, 194, 6292–6293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, N.; Wines, T.F.; Knopp, C.L.; McGuire, M.A.; Tinker, J.K. Expression, immunogenicity and variation of iron-regulated surface protein A from bovine isolates of Staphylococcus aureus. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Hu, C.; Gong, R.; Chen, Y.; Ren, N.; Xiao, G.; Xie, Q.; Zhang, M.; Liu, Q.; Guo, A.; et al. Evaluation of a novel chimeric B cell epitope-based vaccine against mastitis induced by either Streptococcus agalactiae or Staphylococcus aureus in mice. Clin. Vaccine Immunol. 2011, 18, 893–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagliuolo, I.; Piccinini, R.; Beggiao, E.; Palu, G.; Mengoli, C.; Ditadi, F.; Vicenzoni, G.; Zecconi, A. Mucosal genetic immunization against four adhesins protects against Staphylococcus aureus-induced mastitis in mice. Vaccine 2006, 24, 4393–4402. [Google Scholar] [CrossRef]
- Gong, R.; Hu, C.; Xu, H.; Guo, A.; Chen, H.; Zhang, G.; Shi, L. Evaluation of clumping factor A binding region A in a subunit vaccine against Staphylococcus aureus-induced mastitis in mice. Clin. Vaccine Immunol. 2010, 17, 1746–1752. [Google Scholar] [CrossRef] [Green Version]
- Maira-Litran, T.; Bentancor, L.V.; Bozkurt-Guzel, C.; O’Malley, J.M.; Cywes-Bentley, C.; Pier, G.B. Synthesis and Evaluation of a Conjugate Vaccine Composed of Staphylococcus aureus Poly-N-Acetyl-Glucosamine and Clumping Factor A. PLoS ONE 2012, 7, e43813. [Google Scholar] [CrossRef]
- Hawkins, J.; Kodali, S.; Matsuka, Y.V.; McNeil, L.K.; Mininni, T.; Scully, I.L.; Vernachio, J.H.; Severina, E.; Girgenti, D.; Jansen, K.U.; et al. A recombinant Clumping factor A containing vaccine induces functional antibodies to Staphylococcus aureus that are not observed after natural exposure. Clin. Vaccine Immunol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Creech, C.B.; Frenck, R.W.; Fiquet, A.; Feldman, R.; Kankam, M.K.; Pathirana, S.; Baber, J.; Radley, D.; Cooper, D.; Eiden, J.; et al. Persistence of Immune Responses Through 36 Months in Healthy Adults After Vaccination with a Novel. Open Forum Infect. Dis. 2020, 7, ofz532. [Google Scholar] [CrossRef] [Green Version]
- Fluit, A.C.; Terlingen, A.M.; Andriessen, L.; Ikawaty, R.; van Mansfeld, R.; Top, J.; Cohen Stuart, J.W.; Leverstein-van Hall, M.A.; Boel, C.H. Evaluation of the DiversiLab system for detection of hospital outbreaks of infections by different bacterial species. J. Clin. Microbiol. 2010, 48, 3979–3989. [Google Scholar] [CrossRef] [Green Version]
- Gomez, M.I.; Sordelli, D.O.; Buzzola, F.R.; Garcia, V.E. Induction of cell-mediated immunity to Staphylococcus aureus in the mouse mammary gland by local immunization with a live attenuated mutant. Infect. Immun. 2002, 70, 4254–4260. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; O’Brien, C.N.; Guidry, A.J.; Paape, M.J.; Shafer-Weaver, K.A.; Zhao, X. Effect of a trivalent vaccine against Staphylococcus aureus mastitis lymphocyte subpopulations, antibody production, and neutrophil phagocytosis. Can. J. Vet. Res. 2005, 69, 11–18. [Google Scholar] [PubMed]
- Camussone, C.M.; Veaute, C.M.; Porporatto, C.; Morein, B.; Marcipar, I.S.; Calvinho, L.F. Immune response of heifers against a Staphylococcus aureus CP5 whole cell vaccine formulated with ISCOMATRIX adjuvant. J. Dairy Res. 2012, 1–9. [Google Scholar] [CrossRef]
- Riollet, C.; Rainard, P.; Poutrel, B. Cell subpopulations and cytokine expression in cow milk in response to chronic Staphylococcus aureus infection. J. Dairy Sci 2001, 84, 1077–1084. [Google Scholar] [CrossRef]
- Lin, L.; Ibrahim, A.S.; Xu, X.; Farber, J.M.; Avanesian, V.; Baquir, B.; Fu, Y.; French, S.W.; Edwards, J.E., Jr.; Spellberg, B. Th1-Th17 cells mediate protective adaptive immunity against Staphylococcus aureus and Candida albicans infection in mice. PLoS Pathog. 2009, 5, e1000703. [Google Scholar] [CrossRef]
- Snider, D.P. The Mucosal Adjuvant Activities of ADP-Ribosylating Bacterial Enterotoxins. Crit Rev. Immunol. 2017, 37, 499–530. [Google Scholar] [CrossRef] [PubMed]
- George-Chandy, A.; Eriksson, K.; Lebens, M.; Nordstrom, I.; Schon, E.; Holmgren, J. Cholera toxin B subunit as a carrier molecule promotes antigen presentation and increases CD40 and CD86 expression on antigen-presenting cells. Infect. Immun. 2001, 69, 5716–5725. [Google Scholar] [CrossRef] [Green Version]
- Schnitzler, A.C.; Burke, J.M.; Wetzler, L.M. Induction of cell signaling events by the cholera toxin B subunit in antigen-presenting cells. Infect. Immun. 2007, 75, 3150–3159. [Google Scholar] [CrossRef] [Green Version]
- Nashar, T.O.; Hirst, T.R.; Williams, N.A. Modulation of B-cell activation by the B subunit of Escherichia coli enterotoxin: Receptor interaction up-regulates MHC class II, B7, CD40, CD25 and ICAM-1. Immunology 1997, 91, 572–578. [Google Scholar] [CrossRef]
- Bromander, A.K.; Kjerrulf, M.; Holmgren, J.; Lycke, N. Cholera toxin enhances antigen presentation. Adv. Exp. Med. Biol. 1995, 371B, 1501–1506. [Google Scholar]
- Cong, Y.; Weaver, C.T.; Elson, C.O. The mucosal adjuvanticity of cholera toxin involves enhancement of costimulatory activity by selective up-regulation of B7.2 expression. J. Immunol. 1997, 159, 5301–5308. [Google Scholar]
- Eriksson, K.; Fredriksson, M.; Nordstrom, I.; Holmgren, J. Cholera toxin and its B subunit promote dendritic cell vaccination with different influences on Th1 and Th2 development. Infect. Immun. 2003, 71, 1740–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu-Amano, J.; Jackson, R.J.; Fujihashi, K.; Kiyono, H.; Staats, H.F.; McGhee, J.R. Helper Th1 and Th2 cell responses following mucosal or systemic immunization with cholera toxin. Vaccine 1994, 12, 903–911. [Google Scholar] [CrossRef]
- Mattsson, J.; Schon, K.; Ekman, L.; Fahlen-Yrlid, L.; Yrlid, U.; Lycke, N.Y. Cholera toxin adjuvant promotes a balanced Th1/Th2/Th17 response independently of IL-12 and IL-17 by acting on Gsalpha in CD11b(+) DCs. Mucosal Immunol. 2015, 8, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Jobling, M.G.; Holmes, R.K. Fusion proteins containing the A2 domain of cholera toxin assemble with B polypeptides of cholera toxin to form immunoreactive and functional holotoxin-like chimeras. Infect. Immun. 1992, 60, 4915–4924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajishengallis, G.; Hollingshead, S.K.; Koga, T.; Russell, M.W. Mucosal immunization with a bacterial protein antigen genetically coupled to cholera toxin A2/B subunits. J. Immunol. 1995, 154, 4322–4332. [Google Scholar]
- Martin, M.; Hajishengallis, G.; Metzger, D.J.; Michalek, S.M.; Connell, T.D.; Russell, M.W. Recombinant antigen-enterotoxin A2/B chimeric mucosal immunogens differentially enhance antibody responses and B7-dependent costimulation of CD4(+) T cells. Infect. Immun. 2001, 69, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Sultan, F.; Jin, L.L.; Jobling, M.G.; Holmes, R.K.; Stanley, S.L., Jr. Mucosal immunogenicity of a holotoxin-like molecule containing the serine-rich Entamoeba histolytica protein (SREHP) fused to the A2 domain of cholera toxin. Infect. Immun. 1998, 66, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Misra, N.; Wines, T.F.; Knopp, C.L.; Hermann, R.; Bond, L.; Mitchell, B.; McGuire, M.A.; Tinker, J.K. Immunogenicity of a Staphylococcus aureus-cholera toxin A. Vaccine 2018, 36, 3513–3521. [Google Scholar] [CrossRef]
- Prasad, L.B.; Newbould, F.H. Inoculation of the bovine teat duct with Staph. aureus: The relationship of teat duct lenght, milk yield and milking rate to development of intramammary infection. Can. Vet. J. 1968, 9, 107–115. [Google Scholar]
- Arlian, B.M.; Tinker, J.K. Mucosal immunization with a Staphylococcus aureus IsdA-cholera toxin A2/B chimera induces antigen-specific Th2-type responses in mice. Clin. Vaccine Immunol. 2011, 18, 1543–1551. [Google Scholar] [CrossRef] [Green Version]
- Gonzales, V.K.; de Mulder, E.L.; de Boer, T.; Hannink, G.; van Tienen, T.G.; van Heerde, W.L.; Buma, P. Platelet-rich plasma can replace fetal bovine serum in human meniscus cell cultures. Tissue Eng. Part. C Methods 2013, 19, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Coussens, P.M.; Verman, N.; Coussens, M.A.; Elftman, M.D.; McNulty, A.M. Cytokine gene expression in peripheral blood mononuclear cells and tissues of cattle infected with Mycobacterium avium subsp. paratuberculosis: Evidence for an inherent proinflammatory gene expression pattern. Infect. Immun. 2004, 72, 1409–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenz, J.R.; Garry, F.B.; Barrington, G.M. Comparison of disease severity scoring systems for dairy cattle with acute coliform mastitis. J. Am. Vet. Med. Assoc. 2006, 229, 259–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atalla, H.; Gyles, C.; Wilkie, B.; Leslie, K.; Mallard, B. Somatic cell scores and clinical signs following experimental intramammary infection of dairy cows with a Staphylococcus aureus small colony variant (S. aureus SCV) in comparison to other bovine strains. Vet. Microbiol. 2009, 137, 326–334. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shkreta, L.; Talbot, B.G.; Diarra, M.S.; Lacasse, P. Immune responses to a DNA/protein vaccination strategy against Staphylococcus aureus induced mastitis in dairy cows. Vaccine 2004, 23, 114–126. [Google Scholar] [CrossRef]
- Leitner, G.; Lubashevsky, E.; Glickman, A.; Winkler, M.; Saran, A.; Trainin, Z. Development of a Staphylococcus aureus vaccine against mastitis in dairy cows. I. Challenge trials. Vet. Immunol. Immunopathol. 2003, 93, 31–38. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Burvenich, C.; Van Merris, V.; Mehrzad, J.; Diez-Fraile, A.; Duchateau, L. Severity of E. coli mastitis is mainly determined by cow factors. Vet. Res. 2003, 34, 521–564. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.A. Immunity to Staphylococcus aureus: Implications for Vaccine Development. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Wilson, G.J.; Tuffs, S.W.; Wee, B.A.; Seo, K.S.; Park, N.; Connelley, T.; Guinane, C.M.; Morrison, W.I.; Fitzgerald, J.R. Bovine Staphylococcus aureus Superantigens Stimulate the Entire T Cell Repertoire of Cattle. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunther, J.; Petzl, W.; Bauer, I.; Ponsuksili, S.; Zerbe, H.; Schuberth, H.J.; Brunner, R.M.; Seyfert, H.M. Differentiating Staphylococcus aureus from Escherichia coli mastitis: S. aureus triggers unbalanced immune-dampening and host cell invasion immediately after udder infection. Sci Rep. 2017, 7, 4811. [Google Scholar] [CrossRef] [PubMed]
- Schukken, Y.H.; Mallard, B.A.; Dekkers, J.C.; Leslie, K.E.; Stear, M.J. Genetic impact on the risk of intramammary infection following Staphylococcus aureus challenge. J. Dairy Sci. 1994, 77, 639–647. [Google Scholar] [CrossRef]
- Kim, Y.; Atalla, H.; Mallard, B.; Robert, C.; Karrow, N. Changes in Holstein cow milk and serum proteins during intramammary infection with three different strains of Staphylococcus aureus. BMC Vet. Res. 2011, 7, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Wei, Y.; Liu, G.; Shi, B.; Giovanni, S.; Peterson, J.W.; Chopra, A.K. A mutated cholera toxin without the ADP-ribosyltransferase activity induces cytokine production and inhibits apoptosis of splenocytes in mice possibly via toll-like receptor-4 signaling. Mol. Immunol. 2016, 75, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Wiedinger, K.; Pinho, D.; Bitsaktsis, C. Utilization of cholera toxin B as a mucosal adjuvant elicits antibody-mediated protection against S. pneumoniae infection in mice. Ther. Adv. Vaccines 2017, 5, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjuere, F.; George-Chandy, A.; Audant, F.; Rousseau, D.; Holmgren, J.; Czerkinsky, C. Transcutaneous immunization with cholera toxin B subunit adjuvant suppresses IgE antibody responses via selective induction of Th1 immune responses. J. Immunol. 2003, 170, 1586–1592. [Google Scholar] [CrossRef]
- Albu, D.I.; Jones-Trower, A.; Woron, A.M.; Stellrecht, K.; Broder, C.C.; Metzger, D.W. Intranasal vaccination using interleukin-12 and cholera toxin subunit B as adjuvants to enhance mucosal and systemic immunity to human immunodeficiency virus type 1 glycoproteins. J. Virol. 2003, 77, 5589–5597. [Google Scholar] [CrossRef] [Green Version]
- Baldauf, K.J.; Royal, J.M.; Hamorsky, K.T.; Matoba, N. Cholera toxin B: One subunit with many pharmaceutical applications. Toxins 2015, 7, 974–996. [Google Scholar] [CrossRef] [Green Version]
- Holmgren, J.; Harandi, A.M.; Czerkinsky, C. Mucosal adjuvants and anti-infection and anti-immunopathology vaccines based on cholera toxin, cholera toxin B subunit and CpG DNA. Expert Rev. Vaccines 2003, 2, 205–217. [Google Scholar] [CrossRef]
- Royal, J.M.; Matoba, N. Therapeutic Potential of Cholera Toxin B Subunit for the Treatment of Inflammatory Diseases of the Mucosa. Toxins 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Huang, Y.; Lin, Y.; Shan, Y.; Tan, S.; Cai, W.; Li, H.; Zhang, B.; Men, X.; Lu, Z. Anti-inflammatory effect of cholera toxin B subunit in experimental stroke. J. Neuroinflammation 2016, 13, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, M.W.; Moldoveanu, Z.; White, P.L.; Sibert, G.J.; Mestecky, J.; Michalek, S.M. Salivary, nasal, genital, and systemic antibody responses in monkeys immunized intranasally with a bacterial protein antigen and the Cholera toxin B subunit. Infect. Immun. 1996, 64, 1272–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Genotype or Characteristics | Source | |
| E. coli ClearColi® | BL21(DE3) | Lucigen, Madison, WI | |
| S. aureus Newbould 305 | Bovine clinical isolate | [50] | |
| Plasmids | Gene | Vector | Source |
| pLR001 | isdA (Newbould) | pARLDR19 | [49] |
| pLR003 | clfA (Newbould) | pARLDR19 | [49] |
| Bovine Cytokine qPCR Primers | Gene | Amplicon | Source |
| FW 5′-GCATCGTGGAGGGACTTATGA-3′ | GAPDH | 67 | [52] |
| RV 5′-GGGCCATCCACAGTCTTCTG-3′ | |||
| FW 5′-CTTGTCGGAAATGATCCAGTTTT-3′ | IL-10 | 66 | [53] |
| RV 5′-TCAGGCCCGTGGTTCTCA-3′ | |||
| FW 5′-CAGAAAGCGGAAGAGAAGTCAGA-3′ | IFN-γ | 72 | [52] |
| RV 5′-TGCAGGCAGGAGGACCAT-3′ | |||
| FW 5′-GGCTCCCATGATTGTGGTAGTT-3′ | IL-6 | 64 | [53] |
| RV 5′-GCCCAGTGGACAGGTTTCTG-3′ | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alabdullah, H.A.; Overgaard, E.; Scarbrough, D.; Williams, J.E.; Mohammad Mousa, O.; Dunn, G.; Bond, L.; McGuire, M.A.; Tinker, J.K. Evaluation of the Efficacy of a Cholera Toxin-Based Staphylococcus aureus Vaccine against Bovine Intramammary Challenge. Vaccines 2021, 9, 6. https://doi.org/10.3390/vaccines9010006
Alabdullah HA, Overgaard E, Scarbrough D, Williams JE, Mohammad Mousa O, Dunn G, Bond L, McGuire MA, Tinker JK. Evaluation of the Efficacy of a Cholera Toxin-Based Staphylococcus aureus Vaccine against Bovine Intramammary Challenge. Vaccines. 2021; 9(1):6. https://doi.org/10.3390/vaccines9010006
Chicago/Turabian StyleAlabdullah, Hussain A., Elise Overgaard, Danielle Scarbrough, Janet E. Williams, Omid Mohammad Mousa, Gary Dunn, Laura Bond, Mark A. McGuire, and Juliette K. Tinker. 2021. "Evaluation of the Efficacy of a Cholera Toxin-Based Staphylococcus aureus Vaccine against Bovine Intramammary Challenge" Vaccines 9, no. 1: 6. https://doi.org/10.3390/vaccines9010006
APA StyleAlabdullah, H. A., Overgaard, E., Scarbrough, D., Williams, J. E., Mohammad Mousa, O., Dunn, G., Bond, L., McGuire, M. A., & Tinker, J. K. (2021). Evaluation of the Efficacy of a Cholera Toxin-Based Staphylococcus aureus Vaccine against Bovine Intramammary Challenge. Vaccines, 9(1), 6. https://doi.org/10.3390/vaccines9010006