
Major Complex Trait for Early De Novo Programming ‘CoV-MAC-TED’ Detected in Human Nasal Epithelial Cells Infected by Two SARS-CoV-2 Variants Is Promising to Help in Designing Therapeutic Strategies




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
1.1. Is There a Paradigm Shift in Understanding Immunology?
1.2. Resilience Can Depend on the Capacity for Efficient Early Reprogramming—Learning from Plants
1.3. Early Reprogramming Can Link to ROS/RNS Equilibration and Sugar-Dependent Fermentation
1.4. Driving a Standardized Collection of Data on Virus-Induced Early Reprogramming
2. Materials and Methods
2.1. Gene Expression Analyses of RNA-Seq Data from SARS-CoV-2-Infected Human Nasal Epithelial Cells
2.2. Statistical Analyses
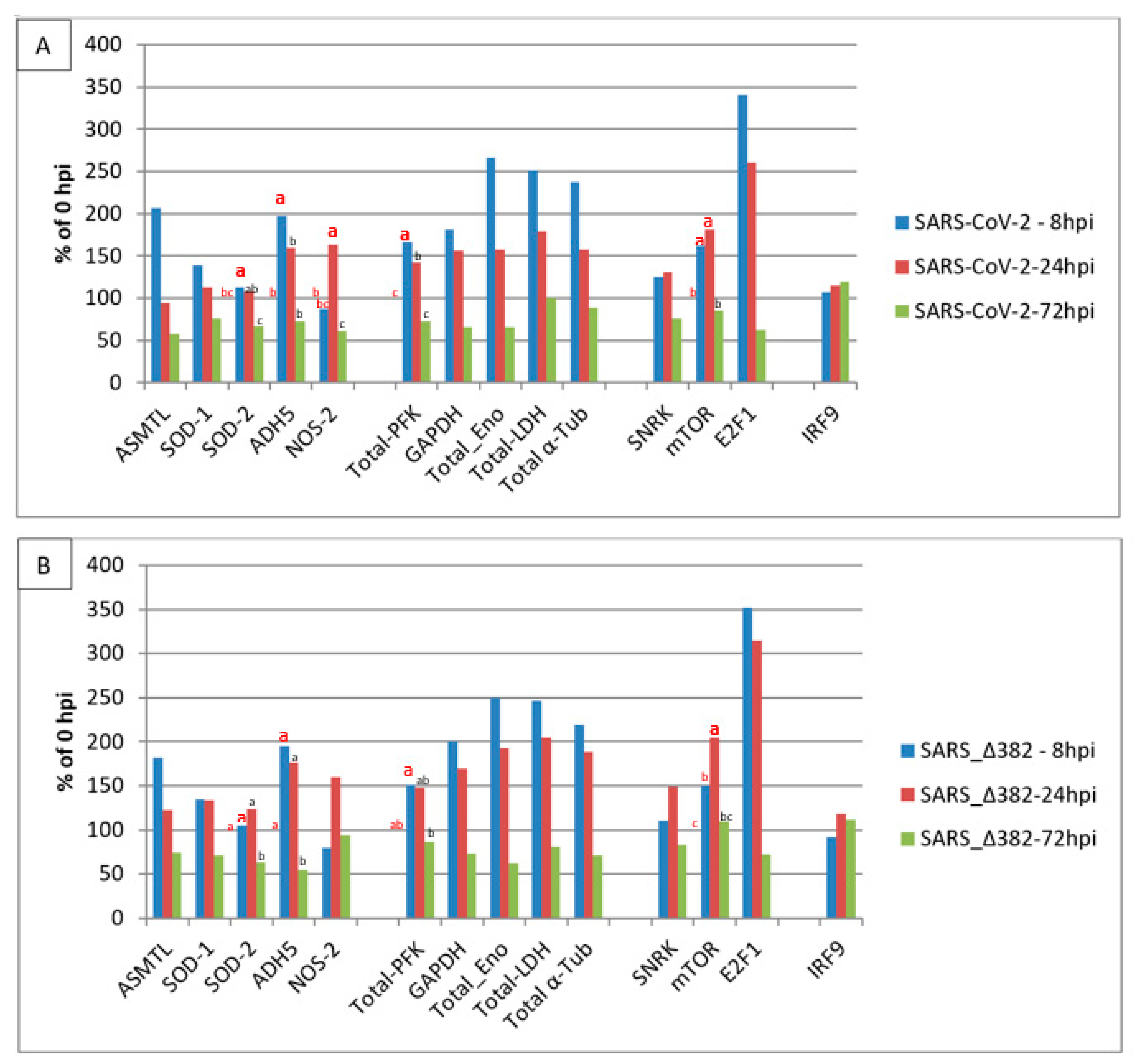
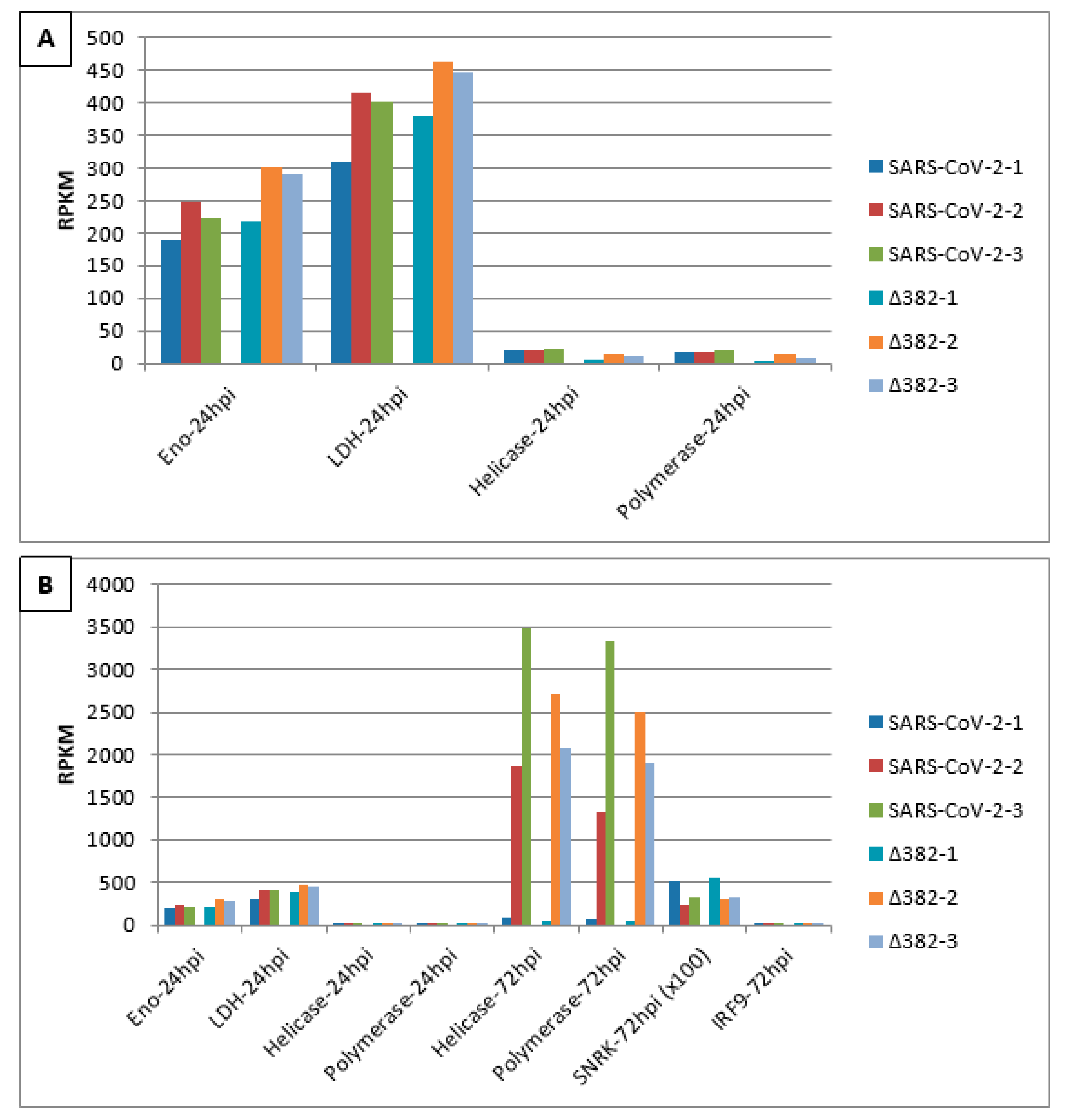
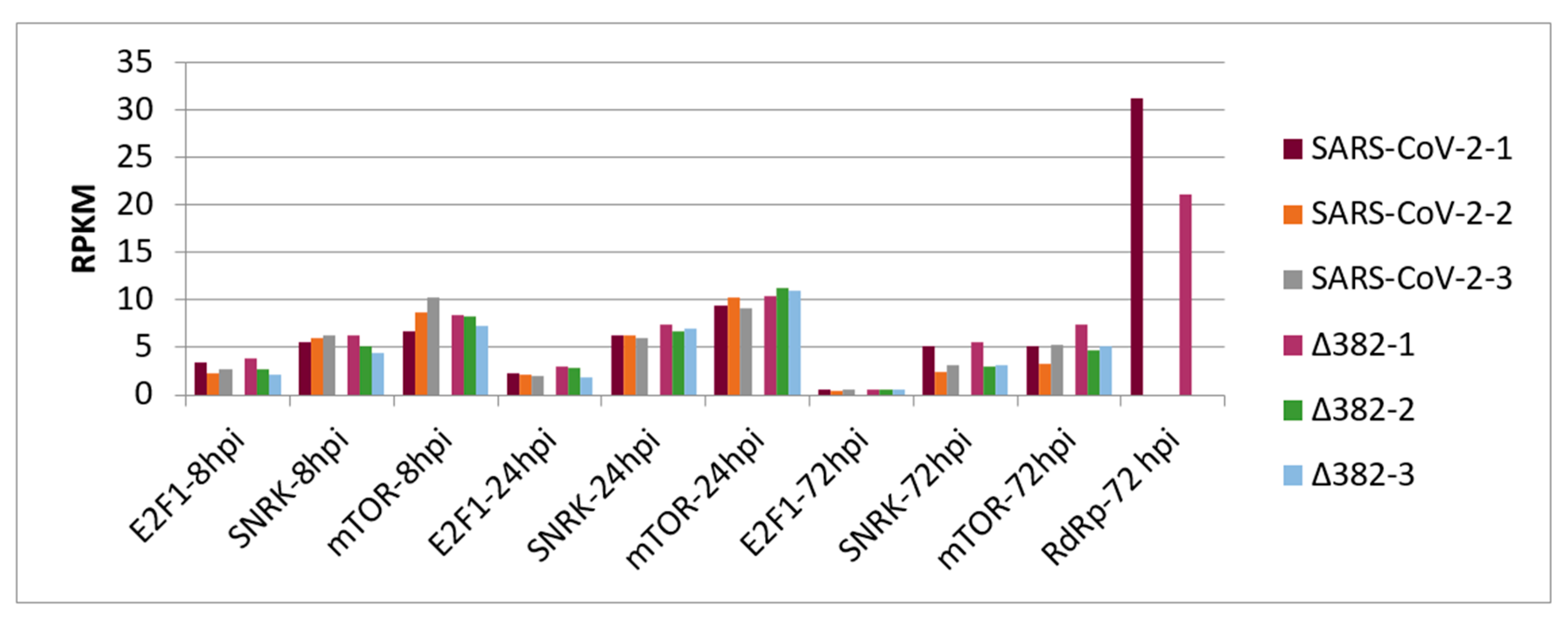
3. Results
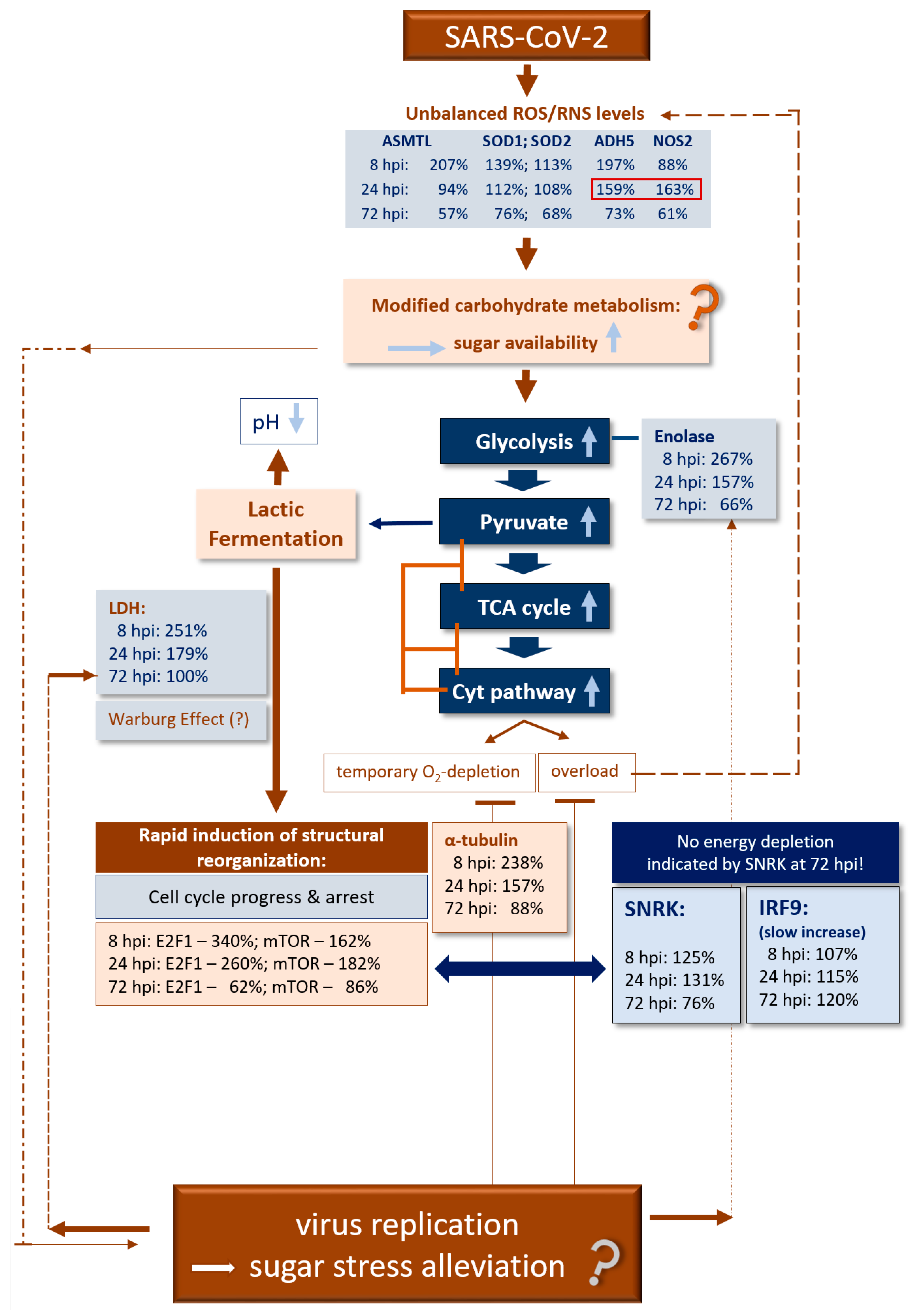
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haney, C.H.; Ausubel, F.M.; Urbach, J.M. Innate Immunity in Plants and Animals: Differences and Similarities. Biochemist 2014, 36, 40–45. [Google Scholar] [CrossRef]
- Gourbal, B.; Pinaud, S.; Beckers, G.J.M.; Van Der Meer, J.W.M.; Conrath, U.; Netea, M.G. Innate Immune Memory: An Evolutionary Perspective. Immunol. Rev. 2018, 283, 21–40. [Google Scholar] [CrossRef]
- Nejat, N.; Mantri, N. Plant Immune System: Crosstalk between Responses to Biotic and Abiotic Stresses the Missing Link in Understanding Plant Defence. Curr. Issues Mol. Biol. 2017, 23, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kirman, J.R.; Quinn, K.M.; Seder, R.A. Immunological Memory. Immunol. Cell Biol. 2019, 97, 615–616. [Google Scholar] [CrossRef]
- Priyadarshini, S.; Aich, P. Effects of Psychological Stress on Innate Immunity and Metabolism in Humans: A Systematic Analysis. PLoS ONE 2012, 7, e43232. [Google Scholar] [CrossRef] [Green Version]
- Delmastro-Greenwood, M.M.; Piganelli, J.D. Changing the Energy of an Immune Response. Am. J. Clin. Exp. Immunol. 2013, 2, 30–54. [Google Scholar] [PubMed]
- Ganeshan, K.; Nikkanen, J.; Man, K.; Leong, Y.A.; Sogawa, Y.; Maschek, J.A.; Van Ry, T.; Chagwedera, D.N.; Cox, J.E.; Chawla, A. Energetic Trade-Offs and Hypometabolic States Promote Disease Tolerance. Cell 2019, 177, 399–413.e12. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D. The Metabolic Spectrum of Memory T Cells. Immunol. Cell Biol. 2019, 97, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Minceva, M. A Thermodynamic Insight into Viral Infections: Do Viruses in a Lytic Cycle Hijack Cell Metabolism Due to Their Low Gibbs Energy? Heliyon 2020, 6, e03933. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Minceva, M. Thermodynamic Insight Into Viral Infections 2: Empirical Formulas, Molecular Compositions and Thermodynamic Properties of SARS, MERS and SARS-CoV-2 (COVID-19) Viruses. Heliyon 2020, 6, e04943. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.H.; Mohanapriya, G.; Revuru, B.; Noceda, C.; Thiers, K.L.L.; Aziz, S.; Arnholdt-Schmitt, B. ROS/RNS balancing, aerobic fermentation regulation and cell cycle control a complex early trait (‘CoV-MAC-TED’) for combating SARS-CoV-2-induced cell reprogramming. Front. Immunol. 2021, 12, 673692. [Google Scholar] [CrossRef]
- Arnholdt-Schmitt, B.; Mohanapriya, G.; Bharadwaj, R.; Noceda, C.; Santos Macedo, E.; Sathishkumar, R. From plant survival under severe stress to anti-viral human defense—A perspective that calls for common efforts. Front. Immunol. 2021, 12, 673723. [Google Scholar] [CrossRef] [PubMed]
- Arnholdt-Schmitt, B.; Mohanapriya, G.; Sathishkumar, R.; Macedo, E.S.; Costa, J.H. Predicting biomass production from plant robustness and germination efficiency by calorespirometry. In Biofuels: Greenhouse Gas Mitigation and Global Warming. Next Generation Biofuels and Role of Biotechnology; Kumar, A., Ogita, S., Yau, Y.Y., Eds.; Springer Nature: Cham, Switzerland, 2018; Chapter 5; ISBN 978-81-322-3761-7. [Google Scholar] [CrossRef]
- Arnholdt-Schmitt, B. Stress-Induced Cell Reprogramming. A Role for Global Genome Regulation? Plant Physiol. 2004, 136, 2579–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnholdt-Schmitt, B. Efficient cell reprogramming as a target for functional marker strategies? Towards new perspectives in applied plant nutrition research. J. Plant Nutr. Soil Sci. 2005, 168, 617–624. [Google Scholar] [CrossRef]
- Arnholdt-Schmitt, B. Functional markers and a systemic strategy: Convergency between plant breeding, plant nutrition and molecular biology. Plant Physiol. Biochem. 2005, 43, 817–820. [Google Scholar] [CrossRef]
- Arnholdt-Schmitt, B.; Costa, J.H.; de Melo, D.F. AOX—A Functional Marker for Efficient Cell Reprogramming Under Stress? Trends Plant Sci. 2006, 11, 281–287. [Google Scholar] [CrossRef]
- Sircar, D.; Cardoso, H.G.; Mukherjee, C.; Mitra, A.; Arnholdt-Schmitt, A. Alternative oxidase (AOX) and phenolicmetabolism in methyl-jasmonate-treated hairy root cultures of Daucus carota L. J. Plant Physiol. 2012, 169, 657–663. [Google Scholar] [CrossRef]
- Cardoso, H.G.; Arnholdt-Schmitt, B. Functional Marker Development Across Species in Selected Traits. In Diagnostics in Plant Breeding; Lübberstedt, T., Varshney, R.K., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 467–515. [Google Scholar] [CrossRef]
- Orfanoudakis, M.; Sinapidou, E.; Arnholdt-Schmitt, B. Can AOX gene diversity mark herbal tea quality? A proposal. In Alternative Respiratory Pathways in Higher Plants; Gupta, K.J., MuR, L.A.J., Neelwarne, B., Eds.; John Wiley & Sons, Inc.: Oxford, UK, 2015; pp. 311–314. ISBN 978-1-118-79046-5. [Google Scholar]
- Arnholdt-Schmitt, B.; Hansen, L.D.; Nogales, A. Calorespirometry, oxygen isotope analysis and functional-marker-assisted selection (‘CalOxy-FMAS’) for genotype screening: A novel concept and tool kit for predicting stable plant growth performance and functional marker identification. Brief. Funct. Genom. 2015, 15, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnholdt-Schmitt, B.; Ragonezi, C.; Cardoso, H. A Central Role of Mitochondria for Stress-Induced Somatic Embryogenesis. In In Vitro Embryogenesis in Higher Plants; Germanà, M.A., Lambardi, M., Eds.; Humana Press: New York, NY, USA, 2016; pp. 87–100. [Google Scholar] [CrossRef]
- Arnholdt-Schmitt, B.; Patil, V.K. Calorespirometry—A novel tool in functional hologenomics to select ‘green’ holobionts for biomass production. Methods Mol. Biol. 2017, 1670, 193–201. [Google Scholar] [CrossRef]
- Arnholdt-Schmitt, B.; Patil, V.K. Studying individual plant AOX gene functionality in early growth regulation—A new approach. Methods Mol. Biol. 2017, 1670, 235–244. [Google Scholar] [CrossRef]
- Mohanapriya, G.; Bharadwaj, R.; Noceda, C.; Costa, J.H.; Kumar, S.R.; Sathishkumar, R.; Thiers, K.L.L.; Santos Macedo, E.; Silva, S.; Annicchiarico, P.; et al. Alternative Oxidase (AOX) Senses Stress Levels to Coordinate Auxin-Induced Reprogramming from Seed Germination to Somatic Embryogenesis—A Role Relevant for Seed Vigor Prediction and Plant Robustness. Front. Plant Sci. 2019, 10, 1134. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, R.B.; Noceda, C.; Gunasekaran, M.; Kumar, S.R.; Thiers, K.L.L.; Costa, J.H.; Arnholdt-Schmitt, B. Adaptive reprogramming during early seed germination requires temporarily enhanced fermentation—A critical role for alternative oxidase (AOX) regulation that concerns also microbiota effectiveness. Front. Plant Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zia, S.F.; Berkowitz, O.; Bedon, F.; Whelan, J.; Franks, A.E.; Plummer, K.M. Direct Comparison of Arabidopsis Gene Expression Reveals Different Responses to Melatonin Versus Auxin. BMC Plant Biol. 2019, 19, 567. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocrinol. 2019, 10, 249. [Google Scholar] [CrossRef]
- Hakkaart, G.A.; Dassa, E.P.; Jacobs, H.T.; Rustin, P. Allotopic Expression of a Mitochondrial Alternative Oxidase Confers Cyanide Resistance to Human Cell Respiration. EMBO Rep. 2006, 7, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvestri, M.; Rossi, G.A. Melatonin: Its Possible Role in the Management of Viral Infections—A Brief Review. Ital. J. Pediatr. 2013, 39, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oláh, J.; Lehotzky, A.; Szunyogh, S.; Szénási, T.; Orosz, F.; Ovádi, J. Microtubule Associated Proteins with Regulatory Functions by Day and Pathological Potency at Night. Cells 2020, 9, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isola, M.; Lilliu, M.A.; Loy, F.; Isola, R. Diabetic Status Influences the Storage of Melatonin in Human Salivary Glands. Anat. Rec. 2018, 301, 711–716. [Google Scholar] [CrossRef]
- Liu, L.; Labani, N.; Cecon, E.; Jockers, R. Melatonin Target Proteins: Too Many or Not Enough? Front. Endocrinol. 2019, 10, 791. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wang, X.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Liu, C.; Reiter, R.J. COVID-19: Melatonin as a Potential Adjuvant Treatment. Life Sci. 2020, 250, 117583. [Google Scholar] [CrossRef]
- Zhou, Y.; Hou, Y.; Shen, J.; Huang, Y.; Martin, W.; Cheng, F. Network-Based Drug Repurposing for Novel Coronavirus 2019-nCoV/SARS-CoV-2. Cell Discov. 2020, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; Liao, C.L.; Chen, S.J.; Shi, L.G.; Lin, L.; Chen, Y.W.; Lin, G.J. Melatonin possesses an anti-influenza potential through its immune modulatory effect. J. Funct. Foods 2019, 58, 189–198. [Google Scholar] [CrossRef]
- Hou, W.; Singh, R.K.; Martins, V.; Tenllado, F.; Franklin, G.; Dias, A.C.P. Transcriptional responses of Hypericum perforatum cells to Agrobacterium tumefaciens and differential gene expression in dark glands. Funct. Plant Biol. 2021, 48, 936–947. [Google Scholar] [CrossRef]
- Juybari, K.B.; Pourhanifeh, M.H.; Hosseinzadeh, A.; Hemati, K.; Mehrzadi, S. Melatonin potentials against viral infections including COVID-19: Current evidence and new findings. Virus Res. 2021, 48, 936. [Google Scholar]
- Fernandes, P.A.; Kinker, G.S.; Navarro, B.V.; Jardim, V.C.; Ribeiro-Paz, E.D.; Córdoba-Moreno, M.O.; Markus, R.P. Melatonin-Index as a biomarker for predicting the distribution of presymptomatic and asymptomatic SARS-CoV-2 carriers. Melatonin Res. 2021, 4, 189–205. [Google Scholar] [CrossRef]
- Shi, Y.-F.; Wang, D.-L.; Wang, C.; Culler, A.H.; Kreiser, M.A.; Suresh, J.; Cohen, J.; Pan, J.; Baker, B.; Liu, J.-Z. Loss of GSNOR1 Function Leads to Compromised Auxin Signaling and Polar Auxin Transport. Mol. Plant 2015, 8, 1350–1365. [Google Scholar] [CrossRef] [Green Version]
- Rizza, S.; Filomeni, G. Role, Targets and Regulation of (De)Nitrosylation in Malignancy. Front. Oncol. 2018, 8, 334. [Google Scholar] [CrossRef] [PubMed]
- Barnett, S.D.; Buxton, I.L.O. The Role of S-Nitrosoglutathione Reductase (GSNOR) in Human Disease and Therapy. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 340–354. [Google Scholar] [CrossRef]
- Akaike, T.; Maeda, H. Nitric Oxide and Virus Infection. Immunology 2000, 101, 300–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey-Serres, J.; Pierik, R.; Ruban, A.; Wingler, A. The Dynamic Plant: Capture, Transformation, and Management of Energy. Plant Physiol. 2018, 176, 961–966. [Google Scholar] [CrossRef] [Green Version]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.-D.; Ogé, L.; Hamama, L.; Atanassova, R. The Sugar-Signaling Hub: Overview of Regulators and Interaction with the Hormonal and Metabolic Network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurzinger, B.; Nukarinen, E.; Nägele, T.; Weckwerth, W.; Teige, M. The SnRK1 Kinase as Central Mediator of Energy Signaling Between Different Organelles. Plant Physiol. 2018, 176, 1085–1094. [Google Scholar] [CrossRef] [Green Version]
- McNulty, S.; Flint, M.; Nichol, S.T.; Spiropoulou, C.F. Host mTORc1 Signaling Regulates Andes Virus Replication. J. Virol. 2013, 87, 912–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamage, A.M.; Tan, K.S.; Chan, W.O.Y.; Liu, J.; Tan, C.W.; Ong, Y.K.; Thong, M.; Andiappan, A.K.; Anderson, D.E.; Wang, Y.; et al. Infection of human Nasal Epithelial Cells with SARS-CoV-2 and a 382-nt deletion isolate lacking ORF8 reveals similar viral kinetics and host transcriptional profiles. PLoS Pathog. 2020, 16, e1009130. [Google Scholar] [CrossRef]
- Young, B.E.; Fong, S.W.; Chan, Y.H.; Mak, T.M.; Ang, L.W.; Anderson, D.E.; Lee, C.Y.; Amrun, S.N.; Lee, B.; Goh, Y.S.; et al. Effects of a major deletion in the SARS-CoV-2 genome on the severity of infection and the inflammatory response: An observational cohort study. Lancet 2020, 396, 603–611. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Thierry-Mieg, J.; Thierry-Mieg, D.; Busby, B.; Madden, T.L. Magic-BLAST, an accurate RNA-seq aligner for long and short reads. BMC Bioinform. 2019, 20, 405. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; Mccue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Hirschauer, N.; Becker, C. Paradigmenwechsel Warum statistische Signifikanztests abgeschafft werden sollten. Signifikanztests Forsch. Lehre 2020, 6, 20. [Google Scholar]
- Wasserstein, R.L.; Lazar, N.A. The ASA Statement on p-Values: Context, Process, and Purpose. Am. Stat. 2016, 70, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Wasserstein, R.L.; Schirm, A.L.; Lazar, N.A. Moving to a World Beyond “p < 0.05”. Am. Stat. 2019, 73, 1–19. [Google Scholar]
- Haaf, J.M.; Ly, A.; Wagenmakers, E.J. Retire significance, but still test hypotheses. Nature 2019, 567, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Academies of Sciences, Engineering, and Medicine. Reproducibility and Replicability in Science; The National Academies Press: Washington, DC, USA, 2019. [Google Scholar]
- Tan, K.S.; Andiappan, A.K.; Lee, B.; Yan, Y.; Liu, J.; Tang, S.A.; Lum, J.; He, T.T.; Ong, Y.K.; Thong, M.; et al. RNA Sequencing of H3N2 Influenza Virus-Infected Human Nasal Epithelial Cells from Multiple Subjects Reveals Molecular Pathways Associated with Tissue Injury and Complications. Cells 2019, 8, 986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, J.H. Transcriptome data from human nasal epithelial cells infected by H3N2 influenza virus indicate early unbalanced ROS/RNA levels, temporarily increased aerobic fermentation linked to enhanced α-tubulin and rapid energy-dependent IRF9-marked immunization. bioRxiv 2021. [Google Scholar] [CrossRef]
- Melkonian, E.A.; Schury, M.P. Biochemistry, Anaerobic Glycolysis. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 9 August 2021. [Google Scholar] [PubMed]
- Kutschera, U.; Pieruschka, R.; Farmer, S.; Berry, J.A. The Warburg-effects: Basic metabolic processes with reference to cancer development and global photosynthesis. Plant Signal. Behav. 2020, 15, 1776477. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, J.H.; Aziz, S.; Noceda, C.; Arnholdt-Schmitt, B. Major Complex Trait for Early De Novo Programming ‘CoV-MAC-TED’ Detected in Human Nasal Epithelial Cells Infected by Two SARS-CoV-2 Variants Is Promising to Help in Designing Therapeutic Strategies. Vaccines 2021, 9, 1399. https://doi.org/10.3390/vaccines9121399
Costa JH, Aziz S, Noceda C, Arnholdt-Schmitt B. Major Complex Trait for Early De Novo Programming ‘CoV-MAC-TED’ Detected in Human Nasal Epithelial Cells Infected by Two SARS-CoV-2 Variants Is Promising to Help in Designing Therapeutic Strategies. Vaccines. 2021; 9(12):1399. https://doi.org/10.3390/vaccines9121399
Chicago/Turabian StyleCosta, José Hélio, Shahid Aziz, Carlos Noceda, and Birgit Arnholdt-Schmitt. 2021. "Major Complex Trait for Early De Novo Programming ‘CoV-MAC-TED’ Detected in Human Nasal Epithelial Cells Infected by Two SARS-CoV-2 Variants Is Promising to Help in Designing Therapeutic Strategies" Vaccines 9, no. 12: 1399. https://doi.org/10.3390/vaccines9121399
APA StyleCosta, J. H., Aziz, S., Noceda, C., & Arnholdt-Schmitt, B. (2021). Major Complex Trait for Early De Novo Programming ‘CoV-MAC-TED’ Detected in Human Nasal Epithelial Cells Infected by Two SARS-CoV-2 Variants Is Promising to Help in Designing Therapeutic Strategies. Vaccines, 9(12), 1399. https://doi.org/10.3390/vaccines9121399