
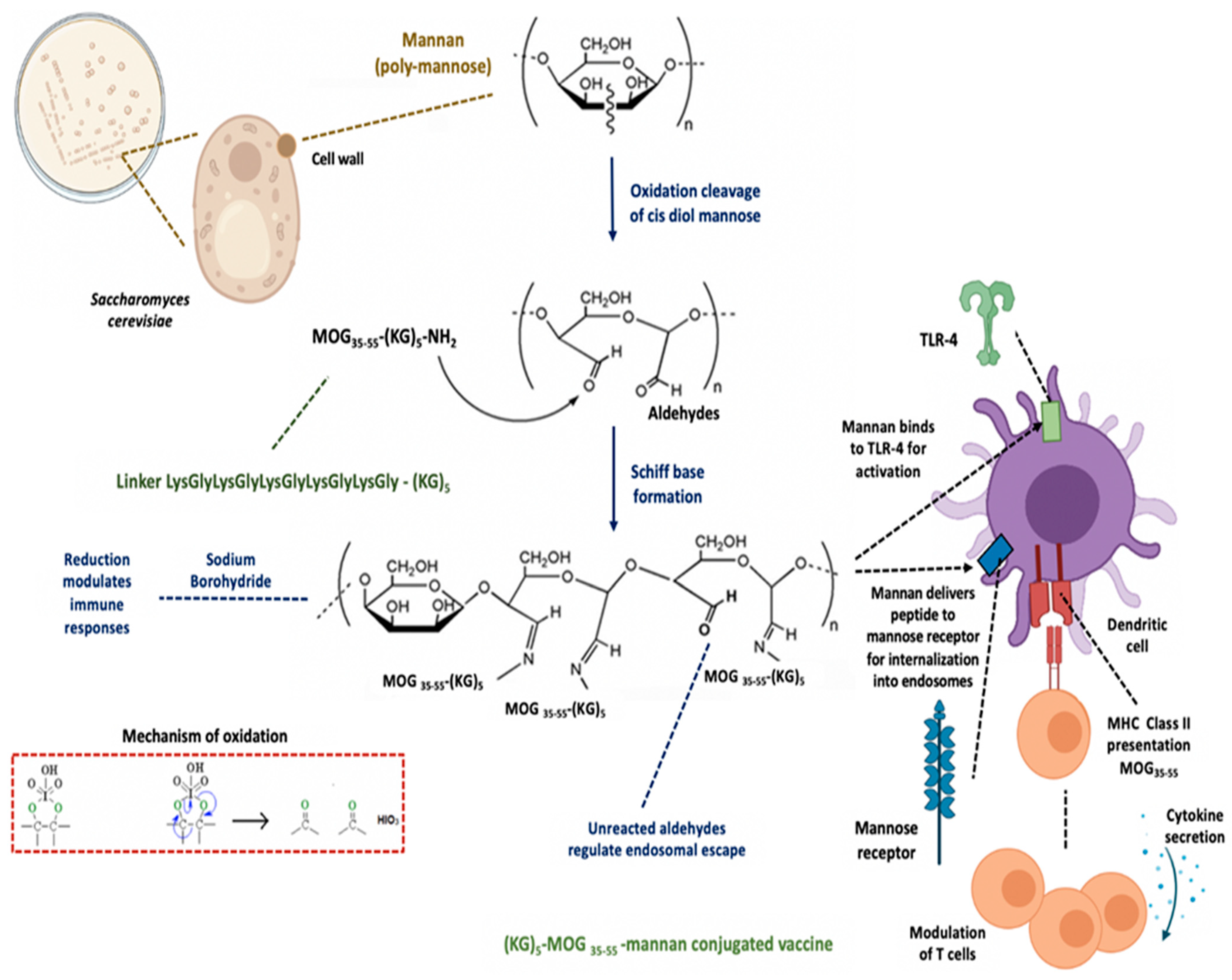
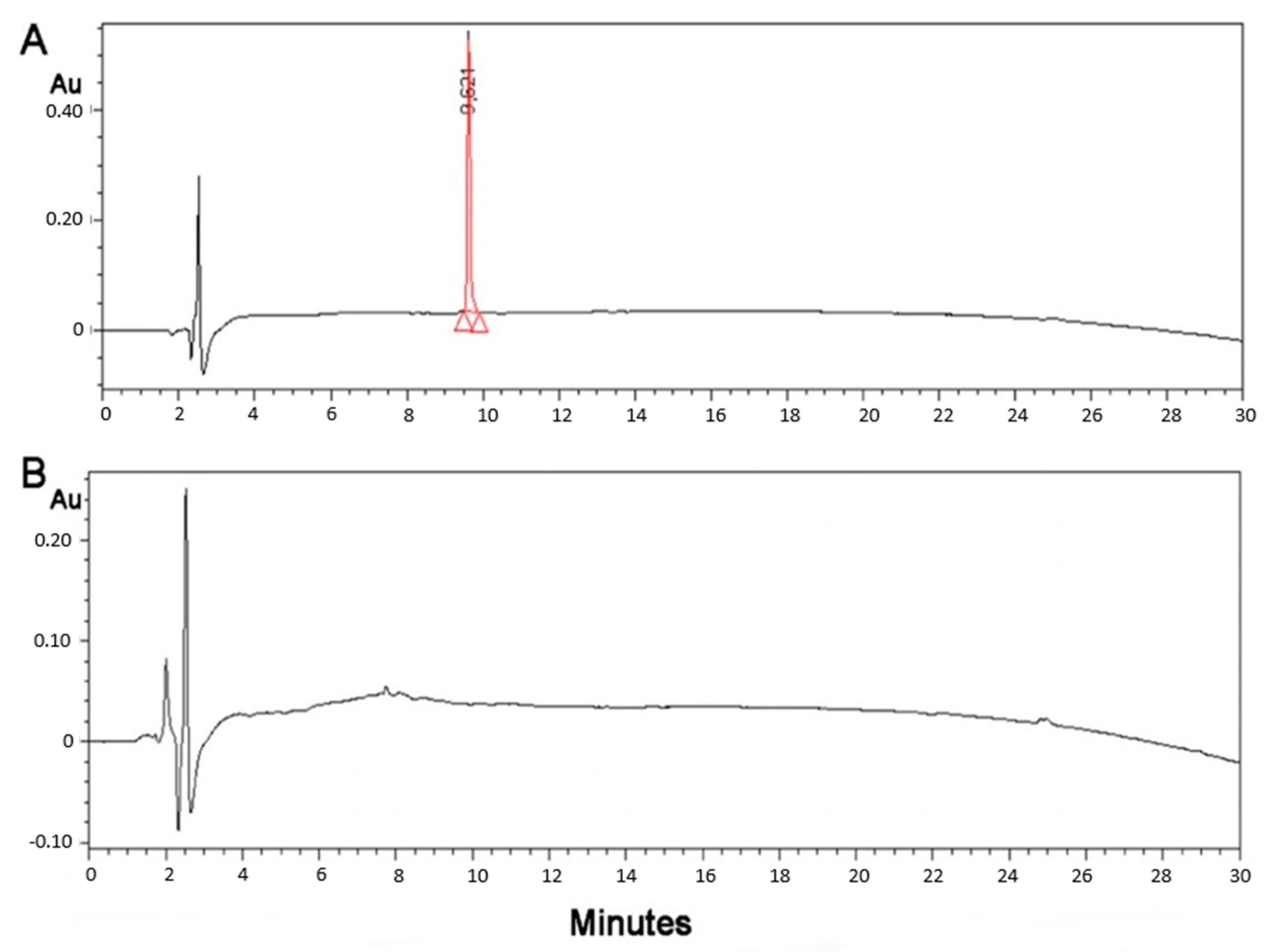
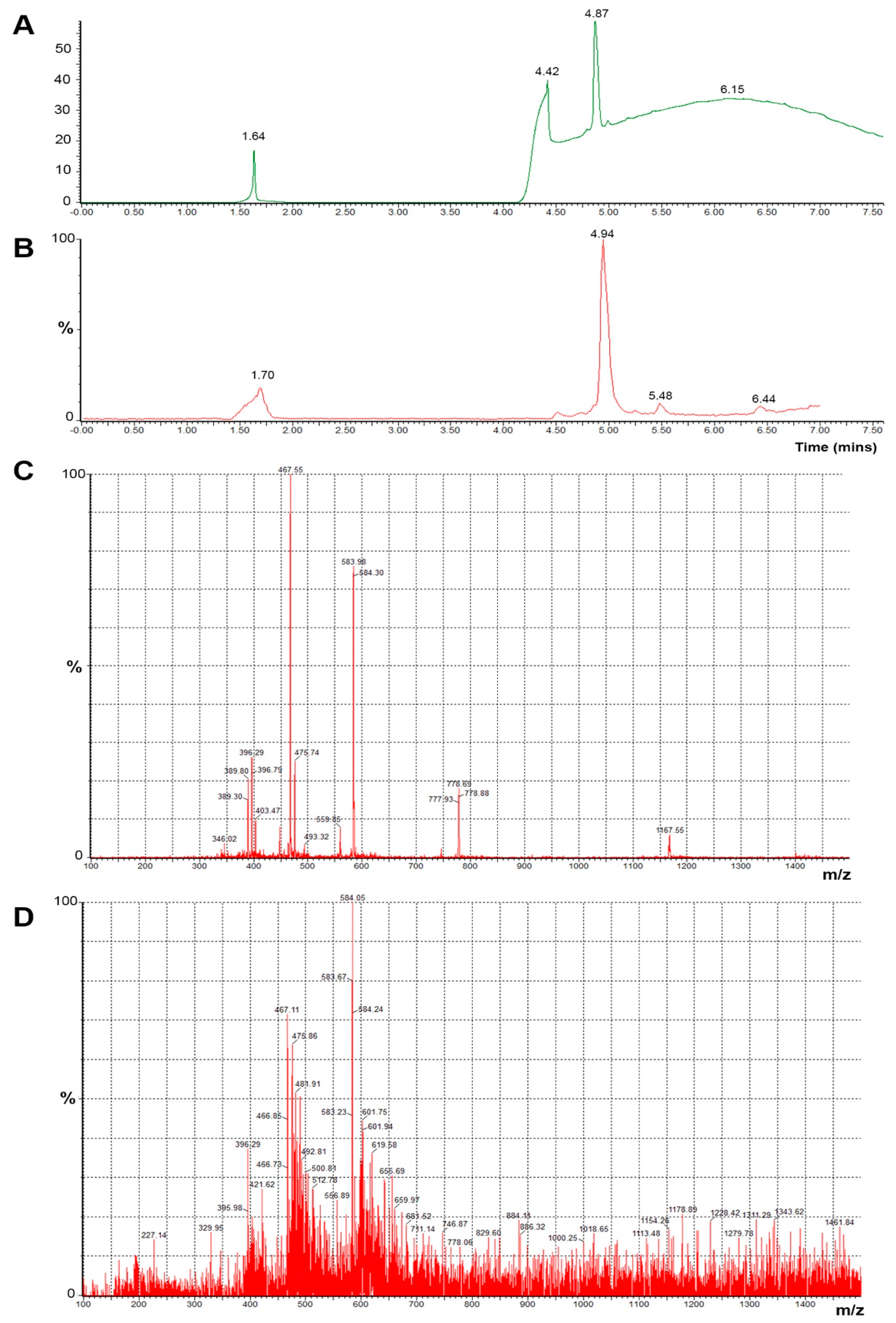
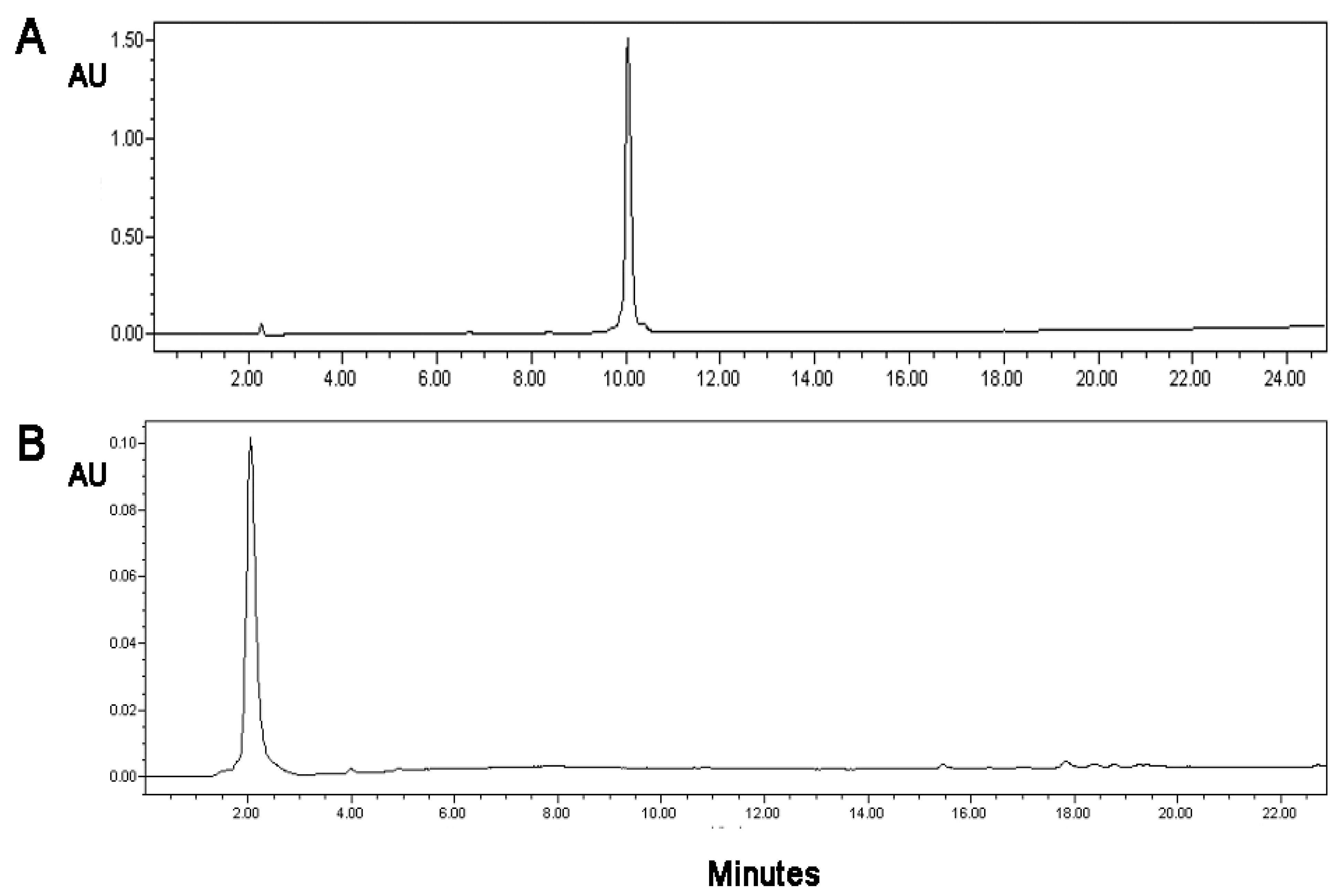
Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient

 ,
, 

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
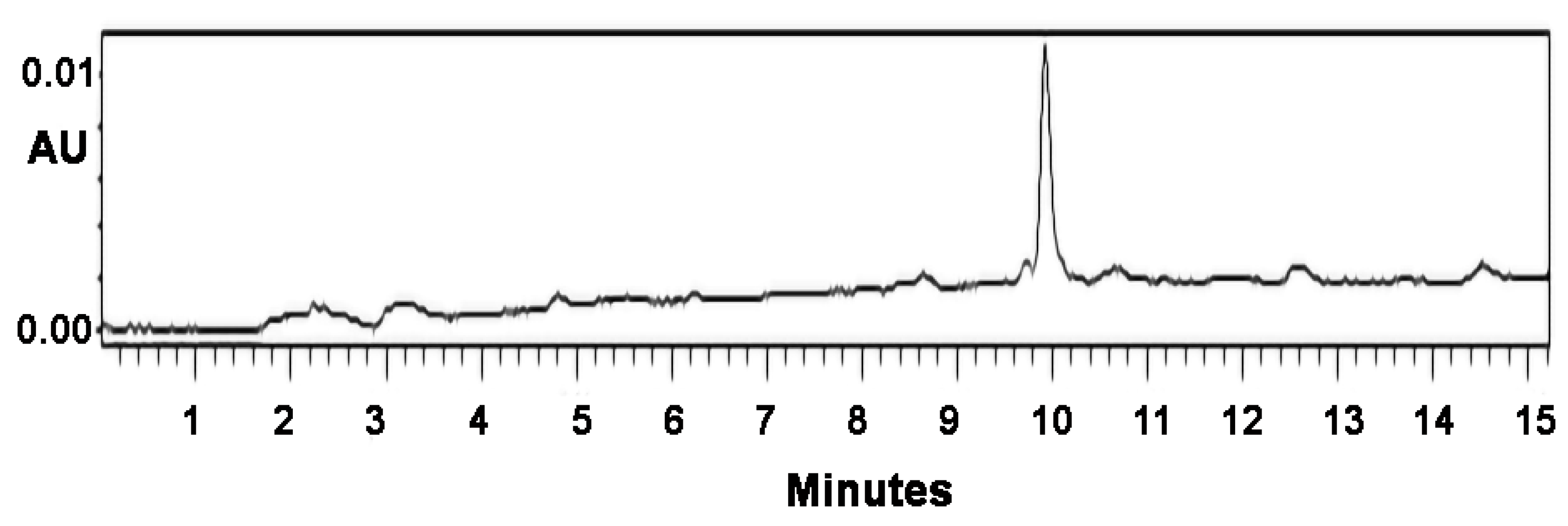
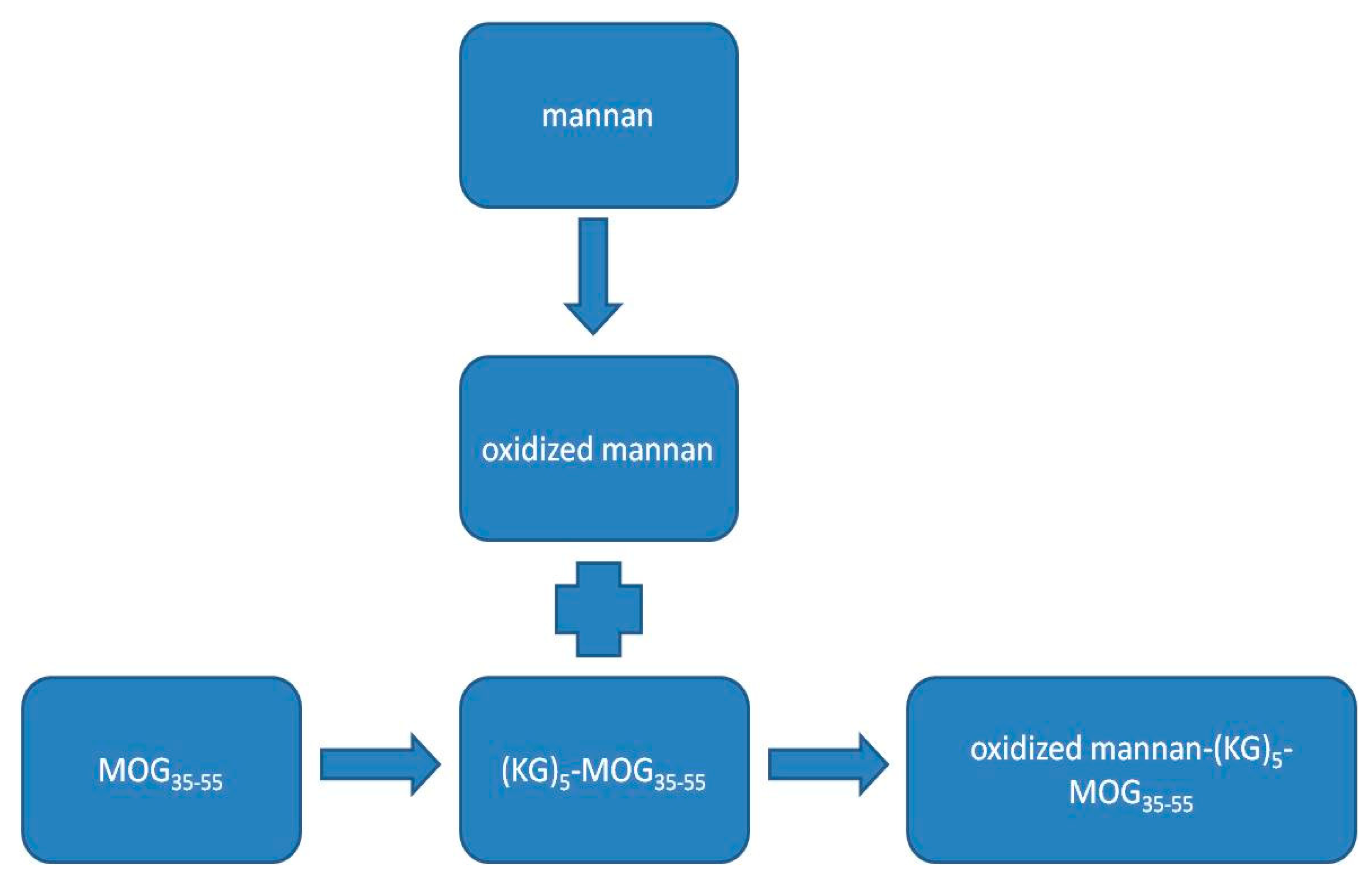
Abstract
Share and Cite
Matsoukas, J.; Deraos, G.; Kelaidonis, K.; Hossain, M.K.; Feehan, J.; Tzakos, A.G.; Matsoukas, E.; Topoglidis, E.; Apostolopoulos, V. Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient. Vaccines 2021, 9, 1456. https://doi.org/10.3390/vaccines9121456
Matsoukas J, Deraos G, Kelaidonis K, Hossain MK, Feehan J, Tzakos AG, Matsoukas E, Topoglidis E, Apostolopoulos V. Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient. Vaccines. 2021; 9(12):1456. https://doi.org/10.3390/vaccines9121456
Chicago/Turabian StyleMatsoukas, John, George Deraos, Kostas Kelaidonis, Md Kamal Hossain, Jack Feehan, Andreas G. Tzakos, Elizabeth Matsoukas, Emmanuel Topoglidis, and Vasso Apostolopoulos. 2021. "Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient" Vaccines 9, no. 12: 1456. https://doi.org/10.3390/vaccines9121456
APA StyleMatsoukas, J., Deraos, G., Kelaidonis, K., Hossain, M. K., Feehan, J., Tzakos, A. G., Matsoukas, E., Topoglidis, E., & Apostolopoulos, V. (2021). Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient. Vaccines, 9(12), 1456. https://doi.org/10.3390/vaccines9121456