
Early and Solid Protection Afforded by the Thiverval Vaccine Provides Novel Vaccination Alternatives Against Classical Swine Fever Virus

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Experimental Design
2.3. Determination of IFN-α Levels in Sera by ELISA Test
2.4. Evaluation of Humoral Response Against CSFV E2 and Erns Glycoproteins and Neutralising Antibodies
2.5. Detection of CSFV RNA
2.6. Statistical Analysis
3. Results
3.1. Thiverval Vaccine Virus Has Weak Replication Capacity and Absence of Transmission among Pigs
3.2. A Single Vaccine Dose Conferred Early and Solid Protection Against CSFV Challenge at 5 and 21 dpv
3.3. The Thiverval-Strain Vaccine Strongly Activates the Innate Immunity in Pigs
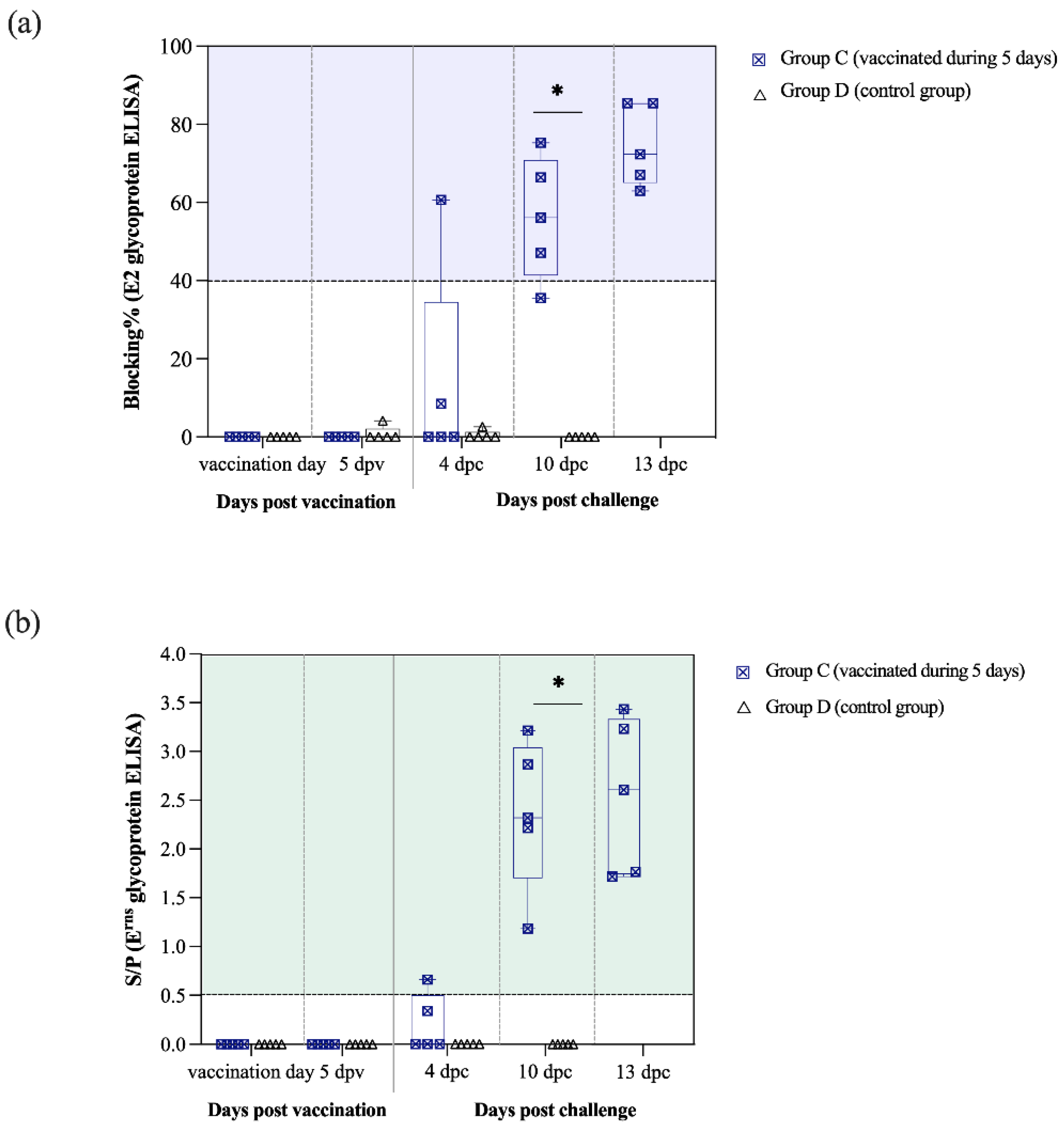
3.4. Antibodies Against CSFV E2 and Erns Glycoproteins with Neutralising Activity Are Detected Two Weeks after Vaccination
3.5. A single Thiverval Vaccine Dose Conferred Protection Against Viral Replication after CSFV Challenge at 21 dpv
3.6. Thiverval Vaccine Generates Rapid CSFV Protection at Five Days after Vaccination in the Absence of Humoral Response
3.7. CSFV-Induced Exacerbated Immune Response Is Prevented by the Thiverval Vaccine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- OIE. Chapter 15.2: Infection with classical swine fever virus. In Terrestrial Animal Health Code; OIE: Paris, France, 2019. [Google Scholar]
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef]
- Blome, S.; Staubach, C.; Henke, J.; Carlson, J.; Beer, M. Classical swine fever—An updated review. Viruses 2017, 9, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rümenapf, T.; Meyers, G.; Stark, R.; Thiel, H.J. Molecular characterization of hog cholera virus. Arch. Virol. Suppl. 1991, 3, 7–18. [Google Scholar]
- Stark, R.; Romenapf, T.; Meyers, G.; Thiel, H.J. Genomic localization of hog cholera virus glycoproteins. Virology 1990, 174, 286–289. [Google Scholar] [CrossRef]
- Rümenapf, T.; Unger, G.; Strauss, J.H.; Thiel, H.J. Processing of the envelope glycoproteins of pestiviruses. J. Virol. 1993, 67, 3288–3294. [Google Scholar] [CrossRef] [Green Version]
- OIE. OIE World Animal Health Information System [WWW Document]. Official Disease Status Interface. Available online: https://www.oie.int/en/animal-health-in-the-world/official-disease-status/classical-swine-fever/list-of-csf-free-member-countries/ (accessed on 23 February 2021).
- Postel, A.; Nishi, T.; Kameyama, K.I.; Meyer, D.; Suckstorff, O.; Fukai, K.; Becher, P. Reemergence of classical swine fever, Japan, 2018. Emerg. Infect. Dis. 2019, 25, 1228–1231. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, Y.; Hayama, Y.; Murato, Y.; Sawai, K.; Yamaguchi, E.; Yamamoto, T. Epidemiology of Classical Swine Fever in Japan—A Descriptive Analysis of the Outbreaks in 2018–2019. Front. Vet. Sci. 2020, 7, 573480. [Google Scholar] [CrossRef] [PubMed]
- Mourits, M.; van Asseldonk, M.; Huirne, R. Multi Criteria Decision Making to evaluate control strategies of contagious animal diseases. Prev. Vet. Med. 2010, 96, 201–210. [Google Scholar] [CrossRef]
- Ganges, L.; Núñez, J.I.; Sobrino, F.; Borrego, B.; Fernández-Borges, N.; Frías-Lepoureau, M.T.; Rodríguez, F. Recent advances in the development of recombinant vaccines against classical swine fever virus: Cellular responses also play a role in protection. Vet. J. 2008, 177, 169–177. [Google Scholar] [CrossRef]
- Qiu, H.J.; Shen, R.X.; Tong, G.Z. The Lapinized Chinese Strain Vaccine Against Classical Swine Fever Virus: A Retrospective Review Spanning Half A Century. Agric. Sci. China 2006, 5, 1–14. [Google Scholar] [CrossRef]
- Coronado, L.; Bohórquez, J.A.; Muñoz-González, S.; Perez, L.J.; Rosell, R.; Fonseca, O.; Delgado, L.; Perera, C.L.; Frías, M.T.; Ganges, L. Investigation of chronic and persistent classical swine fever infections under field conditions and their impact on vaccine efficacy. BMC Vet. Res. 2019, 15, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE. Chapter 3.8.3: Classical Swine Fever Virus (infection with Classical Swine Fever Virus). In OIE Terrestrial Manual; OIE: Paris, France, 2019. [Google Scholar]
- Corthier, G.; Aynaud, J.M. Comparison of the immune response in serum and bucco-pharyngeal secretions following immunization by different routes with a live hog cholera virus vaccine (Thiverval strain). Ann. Rech. Vet. 1977, 8, 159–165. [Google Scholar]
- Aynaud, J.M.; Launais, M. Hog cholera: Immunization of young pigs with the Thiverval-strain vaccine in the presence of colostral immunity. Dev. Biol. Stand. 1978, 41, 381–387. [Google Scholar]
- Muñoz-González, S.; Sordo, Y.; Pérez-Simó, M.; Suárez, M.; Canturri, A.; Rodriguez, M.P.; Frías-Lepoureau, M.T.; Domingo, M.; Estrada, M.P.; Ganges, L. Efficacy of E2 glycoprotein fused to porcine CD154 as a novel chimeric subunit vaccine to prevent classical swine fever virus vertical transmission in pregnant sows. Vet. Microbiol. 2017, 205, 110–116. [Google Scholar] [CrossRef]
- Wensvoort, G.; Terpstra, C.; Boonstra, J.; Bloemraad, M.; Van Zaane, D. Production of monoclonal antibodies against swine fever virus and their use in laboratory diagnosis. Vet. Microbiol. 1986, 12, 101–108. [Google Scholar] [CrossRef]
- Reed, L.J.; Müench, H. A simple method of estimating 50 percent end-points. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Muñoz-González, S.; Pérez-Simó, M.; Colom-Cadena, A.; Cabezón, O.; Bohórquez, J.A.; Rosell, R.; Pérez, L.J.; Marco, I.; Lavín, S.; Domingo, M.; et al. Classical swine fever virus vs. Classical swine fever virus: The superinfection exclusion phenomenon in experimentally infected wild boar. PLoS ONE 2016, 11, e0149469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Liniger, M.; Muñoz-González, S.; Bohórquez, J.A.; Hinojosa, Y.; Gerber, M.; López-Soria, S.; Rosell, R.; Ruggli, N.; Ganges, L. A Polyuridine Insertion in the 3′ Untranslated Region of Classical Swine Fever Virus Activates Immunity and Reduces Viral Virulence in Piglets. J. Virol. 2019, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mearns, R. Abortion in sheep. 1. Investigation and principal causes. Pract. 2007, 29, 40–46. [Google Scholar] [CrossRef]
- Terpstra, C.; Bloemraad, M.; Gielkens, A. The neutralising peroxidase-linked assay for detection of antibody against swine fever virus. Vet. Microbiol. 1984, 9, 113–120. [Google Scholar] [CrossRef]
- Hoffmann, B.; Beer, M.; Schelp, C.; Schirrmeier, H.; Depner, K. Validation of a real-time RT-PCR assay for sensitive and specific detection of classical swine fever. J. Virol. Methods 2005, 130, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.J.; Kwon, T.; Kang, K.; Kim, H.; Kang, S.C.; Richt, J.A.; Lyoo, Y.S. Genetic evolution of classical swine fever virus under immune environments conditioned by genotype 1-based modified live virus vaccine. Transbound. Emerg. Dis. 2018, 65, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Leifer, I.; Lange, E.; Reimann, I.; Blome, S.; Juanola, S.; Duran, J.P.; Beer, M. Modified live marker vaccine candidate CP7_E2alf provides early onset of protection against lethal challenge infection with classical swine fever virus after both intramuscular and oral immunization. Vaccine 2009, 27, 6522–6529. [Google Scholar] [CrossRef]
- Summerfield, A.; Ruggli, N. Immune responses against classical swine fever virus: Between ignorance and lunacy. Front. Vet. Sci. 2015, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-González, S.; Ruggli, N.; Rosell, R.; Pérez, L.J.; Frías-Leuporeau, M.T.; Fraile, L.; Montoya, M.; Cordoba, L.; Domingo, M.; Ehrensperger, F.; et al. Postnatal persistent infection with classical swine fever virus and its immunological implications. PLoS ONE 2015, 10, e0125692. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamothe-Reyes, Y.; Bohórquez, J.A.; Wang, M.; Alberch, M.; Pérez-Simó, M.; Rosell, R.; Ganges, L. Early and Solid Protection Afforded by the Thiverval Vaccine Provides Novel Vaccination Alternatives Against Classical Swine Fever Virus. Vaccines 2021, 9, 464. https://doi.org/10.3390/vaccines9050464
Lamothe-Reyes Y, Bohórquez JA, Wang M, Alberch M, Pérez-Simó M, Rosell R, Ganges L. Early and Solid Protection Afforded by the Thiverval Vaccine Provides Novel Vaccination Alternatives Against Classical Swine Fever Virus. Vaccines. 2021; 9(5):464. https://doi.org/10.3390/vaccines9050464
Chicago/Turabian StyleLamothe-Reyes, Yaneysis, José Alejandro Bohórquez, Miaomiao Wang, Mònica Alberch, Marta Pérez-Simó, Rosa Rosell, and Llilianne Ganges. 2021. "Early and Solid Protection Afforded by the Thiverval Vaccine Provides Novel Vaccination Alternatives Against Classical Swine Fever Virus" Vaccines 9, no. 5: 464. https://doi.org/10.3390/vaccines9050464
APA StyleLamothe-Reyes, Y., Bohórquez, J. A., Wang, M., Alberch, M., Pérez-Simó, M., Rosell, R., & Ganges, L. (2021). Early and Solid Protection Afforded by the Thiverval Vaccine Provides Novel Vaccination Alternatives Against Classical Swine Fever Virus. Vaccines, 9(5), 464. https://doi.org/10.3390/vaccines9050464