
Intranasal Vaccine Delivery Technology for Respiratory Tract Disease Application with a Special Emphasis on Pneumococcal Disease



 and
and
Abstract
:1. Introduction
2. Main Respiratory Illnesses and Their Treatment Methods
3. Intranasal Vaccine Delivery
4. Pneumococcal Disease and the Elderly
5. Intranasal Vaccine Delivery for Pneumococcal Disease
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, P.V. Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Mizgerd, J.P. Respiratory infection and the impact of pulmonary immunity on lung health and disease. Am. J. Respir. Crit. Care Med. 2012, 186, 824–829. [Google Scholar] [CrossRef]
- Heikkinen, T.; Chonmaitree, T. Importance of respiratory viruses in acute otitis media. Clin. Microbiol. Rev. 2003, 16, 230–241. [Google Scholar] [CrossRef] [Green Version]
- Morris, P.S. Upper respiratory tract infections (including otitis media). Pediatr. Clin. N. Am. 2009, 56, 101–117. [Google Scholar] [CrossRef]
- Carvajal, L.A.; Pérez, C.P. Epidemiology of Respiratory Infections. Pediatr. Respir. Dis. 2020, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Treanor, J.; Falsey, A. Respiratory viral infections in the elderly. Antivir. Res. 1999, 44, 79–102. [Google Scholar] [CrossRef]
- Meyer, K.C. Lung infections and aging. Ageing Res. Rev. 2004, 3, 55–67. [Google Scholar] [CrossRef]
- Abelenda-Alonso, G.; Rombauts, A.; Gudiol, C.; Meije, Y.; Ortega, L.; Clemente, M.; Ardanuy, C.; Niubo, J.; Carratala, J. Influenza and Bacterial Coinfection in Adults with Community-Acquired Pneumonia Admitted to Conventional Wards: Risk Factors, Clinical Features, and Outcomes. Open Forum Infect. Dis. 2020, 7, ofaa066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Haber, M.; Ray, S.; Farley, M.M.; Panozzo, C.A.; Klugman, K.P. Invasive pneumococcal pneumonia and respiratory virus co-infections. Emerg. Infect. Dis. 2012, 18, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Kilgore, P.E.; Lim, K.A.; Wang, S.M.; Lee, J.; Deng, W.; Mo, M.Q.; Nyambat, B.; Ma, J.C.; Favorov, M.O.; et al. Influenza and bacterial pathogen coinfections in the 20th century. Interdiscip. Perspect. Infect. Dis. 2011, 2011, 146376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoorob, R.; Sidani, M.A.; Fremont, R.D.; Kihlberg, C. Antibiotic use in acute upper respiratory tract infections. Am. Fam. Physician 2012, 86, 817–822. [Google Scholar]
- Liberati, A.; D’Amico, R.; Pifferi, S.; Torri, V.; Brazzi, L.; Parmelli, E. Antibiotic prophylaxis to reduce respiratory tract infections and mortality in adults receiving intensive care. Cochrane Database Syst. Rev. 2009, CD000022. [Google Scholar] [CrossRef] [Green Version]
- Abed, Y.; Boivin, G. Treatment of respiratory virus infections. Antivir. Res. 2006, 70, 1–16. [Google Scholar] [CrossRef]
- Whitney, C.G.; Harper, S.A. Lower respiratory tract infections: Prevention using vaccines. Infect. Dis. Clin. N. Am. 2004, 18, 899–917. [Google Scholar] [CrossRef]
- Greenberg, H.B.; Piedra, P.A. Immunization against viral respiratory disease: A review. Pediatr. Infect. Dis. J. 2004, 23, S254–S261. [Google Scholar] [CrossRef] [PubMed]
- Ortman, J.M.; Velkoff, V.A. An Aging Nation: The Older Population in the United States; Current Population Reports. 2014. Available online: https://time.com/wp-content/uploads/2015/01/p25-1140.pdf (accessed on 1 March 2021).
- Chow, E.J.; Doyle, J.D.; Uyeki, T.M. Influenza virus-related critical illness: Prevention, diagnosis, treatment. Crit. Care 2019, 23, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, A.; Cilloniz, C.; Blasi, F.; Chalmers, J.D.; Gaillat, J.; Dartois, N.; Schmitt, H.J.; Welte, T. Burden of pneumococcal community-acquired pneumonia in adults across Europe: A literature review. Respir. Med. 2018, 137, 6–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, M.B.; Passos, S.D. Severe Pertussis in Childhood: Update and Controversy-Systematic Review. Rev. Paul. Pediatr. 2019, 37, 351–362. [Google Scholar] [CrossRef]
- Suarez, I.; Funger, S.M.; Kroger, S.; Rademacher, J.; Fatkenheuer, G.; Rybniker, J. The Diagnosis and Treatment of Tuberculosis. Dtsch. Arztebl. Int. 2019, 116, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Stasi, C.; Fallani, S.; Voller, F.; Silvestri, C. Treatment for COVID-19: An overview. Eur. J. Pharmacol. 2020, 889, 173644. [Google Scholar] [CrossRef]
- Liu, J.; Xie, W.; Wang, Y.; Xiong, Y.; Chen, S.; Han, J.; Wu, Q. A comparative overview of COVID-19, MERS and SARS: Review article. Int. J. Surg. 2020, 81, 1–8. [Google Scholar] [CrossRef]
- Smith, T.; Wolff, K.A.; Nguyen, L. Molecular biology of drug resistance in Mycobacterium tuberculosis. Curr. Top. Microbiol. Immunol. 2013, 374, 53–80. [Google Scholar] [CrossRef] [Green Version]
- Bellerose, M.M.; Proulx, M.K.; Smith, C.M.; Baker, R.E.; Ioerger, T.R.; Sassetti, C.M. Distinct Bacterial Pathways Influence the Efficacy of Antibiotics against Mycobacterium tuberculosis. mSystems 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Gideon, H.P.; Flynn, J.L. Latent tuberculosis: What the host “sees”? Immunol. Res. 2011, 50, 202–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulka, K.; Hatfull, G.; Ojha, A.K. Growth of Mycobacterium tuberculosis biofilms. J. Vis. Exp. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjelievskaia, J.; Erck, D.; Piracha, S.; Schrager, L. Drug-resistant TB: Deadly, costly and in need of a vaccine. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Barkai, G.; Greenberg, D.; Givon-Lavi, N.; Dreifuss, E.; Vardy, D.; Dagan, R. Community prescribing and resistant Streptococcus pneumoniae. Emerg. Infect. Dis. 2005, 11, 829–837. [Google Scholar] [CrossRef]
- Butler, J.C.; Cetron, M.S. Pneumococcal drug resistance: The new “special enemy of old age”. Clin. Infect. Dis. 1999, 28, 730–735. [Google Scholar] [CrossRef] [Green Version]
- Dagan, R.; Leibovitz, E.; Greenberg, D.; Yagupsky, P.; Fliss, D.M.; Leiberman, A. Dynamics of pneumococcal nasopharyngeal colonization during the first days of antibiotic treatment in pediatric patients. Pediatr. Infect. Dis. J. 1998, 17, 880–885. [Google Scholar] [CrossRef]
- Pichichero, M.E.; Casey, J.R. Emergence of a multiresistant serotype 19A pneumococcal strain not included in the 7-valent conjugate vaccine as an otopathogen in children. JAMA 2007, 298, 1772–1778. [Google Scholar] [CrossRef] [Green Version]
- Marks, L.R.; Parameswaran, G.I.; Hakansson, A.P. Pneumococcal interactions with epithelial cells are crucial for optimal biofilm formation and colonization in vitro and in vivo. Infect. Immun. 2012, 80, 2744–2760. [Google Scholar] [CrossRef] [Green Version]
- Loughran, A.J.; Orihuela, C.J.; Tuomanen, E.I. Streptococcus pneumoniae: Invasion and Inflammation. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Chen, H.H.; Stringer, A.; Eguale, T.; Rao, G.G.; Ozawa, S. Impact of Antibiotic Resistance on Treatment of Pneumococcal Disease in Ethiopia: An Agent-Based Modeling Simulation. Am. J. Trop. Med. Hyg. 2019, 101, 1042–1053. [Google Scholar] [CrossRef]
- Kim, L.; McGee, L.; Tomczyk, S.; Beall, B. Biological and Epidemiological Features of Antibiotic-Resistant Streptococcus pneumoniae in Pre- and Post-Conjugate Vaccine Eras: A United States Perspective. Clin. Microbiol. Rev. 2016, 29, 525–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mabhula, A.; Singh, V. Drug-resistance in Mycobacterium tuberculosis: Where we stand. Medchemcomm 2019, 10, 1342–1360. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yao, K.; Ma, X.; Shi, W.; Yuan, L.; Yang, Y. Variation in Bordetella pertussis Susceptibility to Erythromycin and Virulence-Related Genotype Changes in China (1970–2014). PLoS ONE 2015, 10, e0138941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Wang, Z.; Luan, Y.; Li, Y.; Liu, X.; Peng, X.; Octavia, S.; Payne, M.; Lan, R. Genomic epidemiology of erythromycin-resistant Bordetella pertussis in China. Emerg. Microbes Infect. 2019, 8, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamachi, K.; Duong, H.T.; Dang, A.D.; Hai, T.; Do, D.; Koide, K.; Otsuka, N.; Shibayama, K.; Hoang, H.T.T. Macrolide-Resistant Bordetella pertussis, Vietnam, 2016–2017. Emerg. Infect. Dis. 2020, 26, 2511–2513. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, H.; Kett, V. Current prospects and future challenges for nasal vaccine delivery. Hum. Vaccines Immunother. 2017, 13, 34–45. [Google Scholar] [CrossRef]
- Ogra, P.L.; Faden, H.; Welliver, R.C. Vaccination strategies for mucosal immune responses. Clin. Microbiol. Rev. 2001, 14, 430–445. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Peng, Y.; Xu, H.; Cui, Z.; Williams, R.O. The COVID-19 Vaccine Race: Challenges and Opportunities in Vaccine Formulation. AAPS PharmSciTech 2020, 21, 225. [Google Scholar] [CrossRef]
- Tomar, J.; Patil, H.P.; Bracho, G.; Tonnis, W.F.; Frijlink, H.W.; Petrovsky, N.; Vanbever, R.; Huckriede, A.; Hinrichs, W.L.J. Advax augments B and T cell responses upon influenza vaccination via the respiratory tract and enables complete protection of mice against lethal influenza virus challenge. J. Control. Release 2018, 288, 199–211. [Google Scholar] [CrossRef]
- Dehghan, S.; Tafaghodi, M.; Bolourieh, T.; Mazaheri, V.; Torabi, A.; Abnous, K.; Tavassoti Kheiri, M. Rabbit nasal immunization against influenza by dry-powder form of chitosan nanospheres encapsulated with influenza whole virus and adjuvants. Int. J. Pharm. 2014, 475, 1–8. [Google Scholar] [CrossRef]
- Li, H.S.; Shin, M.K.; Singh, B.; Maharjan, S.; Park, T.E.; Kang, S.K.; Yoo, H.S.; Hong, Z.S.; Cho, C.S.; Choi, Y.J. Nasal immunization with mannan-decorated mucoadhesive HPMCP microspheres containing ApxIIA toxin induces protective immunity against challenge infection with Actinobacillus pleuropneumoiae in mice. J. Control. Release 2016, 233, 114–125. [Google Scholar] [CrossRef]
- Wu, M.; Zhao, H.; Li, M.; Yue, Y.; Xiong, S.; Xu, W. Intranasal Vaccination with Mannosylated Chitosan Formulated DNA Vaccine Enables Robust IgA and Cellular Response Induction in the Lungs of Mice and Improves Protection against Pulmonary Mycobacterial Challenge. Front. Cell. Infect. Microbiol. 2017, 7, 445. [Google Scholar] [CrossRef]
- Reljic, R.; Sibley, L.; Huang, J.M.; Pepponi, I.; Hoppe, A.; Hong, H.A.; Cutting, S.M. Mucosal vaccination against tuberculosis using inert bioparticles. Infect. Immun. 2013, 81, 4071–4080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khademi, F.; Derakhshan, M.; Yousefi-Avarvand, A.; Najafi, A.; Tafaghodi, M. A novel antigen of Mycobacterium tuberculosis and MPLA adjuvant co-entrapped into PLGA: DDA hybrid nanoparticles stimulates mucosal and systemic immunity. Microb. Pathog. 2018, 125, 507–513. [Google Scholar] [CrossRef]
- Wang, D.; Lu, J.; Yu, J.; Hou, H.; Leenhouts, K.; Van Roosmalen, M.L.; Gu, T.; Jiang, C.; Kong, W.; Wu, Y. A Novel PspA Protein Vaccine Intranasal Delivered by Bacterium-Like Particles Provides Broad Protection against Pneumococcal Pneumonia in Mice. Immunol. Investig. 2018, 47, 403–415. [Google Scholar] [CrossRef]
- Shim, B.S.; Choi, Y.K.; Yun, C.H.; Lee, E.G.; Jeon, Y.S.; Park, S.M.; Cheon, I.S.; Joo, D.H.; Cho, C.H.; Song, M.S.; et al. Sublingual immunization with M2-based vaccine induces broad protective immunity against influenza. PLoS ONE 2011, 6, e27953. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Patil, H.P.; de Vries-Idema, J.; Wilschut, J.; Huckriede, A. Evaluation of mucosal and systemic immune responses elicited by GPI-0100- adjuvanted influenza vaccine delivered by different immunization strategies. PLoS ONE 2013, 8, e69649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, C.J.; Ault, A.; Sivasubramani, S.K.; Gorres, J.P.; Wei, C.J.; Andersen, H.; Gall, J.; Roederer, M.; Rao, S.S. Aerosolized adenovirus-vectored vaccine as an alternative vaccine delivery method. Respir. Res. 2011, 12, 153. [Google Scholar] [CrossRef] [Green Version]
- Kong, I.G.; Sato, A.; Yuki, Y.; Nochi, T.; Takahashi, H.; Sawada, S.; Mejima, M.; Kurokawa, S.; Okada, K.; Sato, S.; et al. Nanogel-based PspA intranasal vaccine prevents invasive disease and nasal colonization by Streptococcus pneumoniae. Infect. Immun. 2013, 81, 1625–1634. [Google Scholar] [CrossRef] [Green Version]
- Chong, C.P.; Street, P.R. Pneumonia in the elderly: A review of the epidemiology, pathogenesis, microbiology, and clinical features. South. Med. J. 2008, 101, 1141–1145, quiz 1132, 1179. [Google Scholar] [CrossRef]
- Wunderink, R.G.; Waterer, G. Advances in the causes and management of community acquired pneumonia in adults. BMJ 2017, 358, j2471. [Google Scholar] [CrossRef]
- Stupka, J.E.; Mortensen, E.M.; Anzueto, A.; Restrepo, M.I. Community-acquired pneumonia in elderly patients. Aging Health 2009, 5, 763–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiri, T.; Khan, K.; Keaney, K.; Mukherjee, G.; McCarthy, N.D.; Petrou, S. Pneumococcal Disease: A Systematic Review of Health Utilities, Resource Use, Costs, and Economic Evaluations of Interventions. Value Health 2019, 22, 1329–1344. [Google Scholar] [CrossRef]
- de Sevilla, M.F.; Garcia-Garcia, J.J.; Esteva, C.; Moraga, F.; Hernandez, S.; Selva, L.; Coll, F.; Ciruela, P.; Planes, A.M.; Codina, G.; et al. Clinical presentation of invasive pneumococcal disease in Spain in the era of heptavalent conjugate vaccine. Pediatr. Infect. Dis. J. 2012, 31, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Short, K.R.; Habets, M.N.; Hermans, P.W.; Diavatopoulos, D.A. Interactions between Streptococcus pneumoniae and influenza virus: A mutually beneficial relationship? Future Microbiol. 2012, 7, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Foxman, B.; Weinberger, D.M.; Steiner, C.; Viboud, C.; Rohani, P. Identifying the interaction between influenza and pneumococcal pneumonia using incidence data. Sci. Transl. Med. 2013, 5, 191ra84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.T.; Louwen, R.; Elberse, K.; van Amerongen, G.; Yuksel, S.; Luijendijk, A.; Osterhaus, A.D.; Duprex, W.P.; de Swart, R.L. Streptococcus pneumoniae Enhances Human Respiratory Syncytial Virus Infection In Vitro and In Vivo. PLoS ONE 2015, 10, e0127098. [Google Scholar] [CrossRef] [Green Version]
- Kadioglu, A.; Weiser, J.N.; Paton, J.C.; Andrew, P.W. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Microbiol. 2008, 6, 288–301. [Google Scholar] [CrossRef]
- Boe, D.M.; Boule, L.A.; Kovacs, E.J. Innate immune responses in the ageing lung. Clin. Exp. Immunol. 2017, 187, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganaie, F.; Saad, J.S.; McGee, L.; van Tonder, A.J.; Bentley, S.D.; Lo, S.W.; Gladstone, R.A.; Turner, P.; Keenan, J.D.; Breiman, R.F.; et al. A New Pneumococcal Capsule Type, 10D, is the 100th Serotype and Has a Large cps Fragment from an Oral Streptococcus. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Obaro, S.; Adegbola, R. The pneumococcus: Carriage, disease and conjugate vaccines. J. Med. Microbiol. 2002, 51, 98–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Microbiol. 2018, 16, 355–367. [Google Scholar] [CrossRef]
- Henig, O.; Kaye, K.S. Bacterial Pneumonia in Older Adults. Infect. Dis. Clin. N. Am. 2017, 31, 689–713. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/pneumococcal/about/facts.html (accessed on 1 March 2021).
- Centers for Disease Control and Prevention. Active Bacterial Core Surveillance Report, Emerging Infections Program. Network, Streptococcus Pneumoniae. 2017. Available online: https://www.cdc.gov/abcs/reports-findings/survreports/spneu17.html (accessed on 1 March 2021).
- Centers for Disease Control and Prevention. Active Bacterial Core Surveillance Report EIPN, Group A Streptococcus—2015, Findings/Survreports/Gas15.pdf Avtihwcgar-2015. 2015. Available online: https://www.cdc.gov/abcs/reports-findings/survreports/gas15.html (accessed on 1 March 2021).
- Centers for Disease Control and Prevention. Active Bacterial Core Surveillance Report EIPN, Streptococcus Pneumoniae. 2016. Available online: http://www.cdc.gov/abcs/reports-findings/survreports/spneu16.pdf (accessed on 1 March 2021).
- Wroe, P.C.; Finkelstein, J.A.; Ray, G.T.; Linder, J.A.; Johnson, K.M.; Rifas-Shiman, S.; Moore, M.R.; Huang, S.S. Aging population and future burden of pneumococcal pneumonia in the United States. J. Infect. Dis. 2012, 205, 1589–1592. [Google Scholar] [CrossRef]
- Drijkoningen, J.J.; Rohde, G.G. Pneumococcal infection in adults: Burden of disease. Clin. Microbiol. Infect. 2014, 20 (Suppl. 5), 45–51. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Active Bacterial Core Surveillance Report EIPN, S.p. 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 1 March 2021).
- Thompson, M.G.; Shay, D.K.; Zhou, H.; Bridges, C.B.; Cheng, P.Y.; Burns, E.; Bresee, J.S.; Cox, N.J. Estimates of Deaths Associated with Seasonal Influenza-United States, 1976–2007. JAMA J. Am. Med Assoc. 2010, 304, 1778–1780, reprinted in MMWR 2010, 59, 1057–1062. [Google Scholar]
- Chen, W.H.; Kozlovsky, B.F.; Effros, R.B.; Grubeck-Loebenstein, B.; Edelman, R.; Sztein, M.B. Vaccination in the elderly: An immunological perspective. Trends Immunol. 2009, 30, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Hanage, W.P. Serotype-specific problems associated with pneumococcal conjugate vaccination. Future Microbiol. 2008, 3, 23–30. [Google Scholar] [CrossRef]
- Berical, A.C.; Harris, D.; Dela Cruz, C.S.; Possick, J.D. Pneumococcal Vaccination Strategies. An Update and Perspective. Ann. Am. Thorac. Soc. 2016, 13, 933–944. [Google Scholar] [CrossRef] [Green Version]
- Musher, D. Pneumococcal Vaccination in Adults; Bartlett, J.G., Ed.; Wolters Kluwer: Alphen aan den Rijn, The Netherlands, 2017. [Google Scholar]
- Luck, J.N.; Tettelin, H.; Orihuela, C.J. Sugar-Coated Killer: Serotype 3 Pneumococcal Disease. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.E.; Robinson, D.A.; McDaniel, L.S. Nonencapsulated Streptococcus pneumoniae: Emergence and Pathogenesis. mBio 2016, 7, e01792. [Google Scholar] [CrossRef] [Green Version]
- Chao, Y.; Marks, L.R.; Pettigrew, M.M.; Hakansson, A.P. Streptococcus pneumoniae biofilm formation and dispersion during colonization and disease. Front. Cell. Infect. Microbiol. 2014, 4, 194. [Google Scholar] [CrossRef] [Green Version]
- Marks, L.R.; Davidson, B.A.; Knight, P.R.; Hakansson, A.P. Interkingdom signaling induces Streptococcus pneumoniae biofilm dispersion and transition from asymptomatic colonization to disease. mBio 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Blanchette-Cain, K.; Hinojosa, C.A.; Babu, R.A.S.; Lizcano, A.; Gonzalez-Juarbe, N.; Munoz-Almagro, C.; Sanchez, C.J.; Bergman, M.A.; Orihuela, C.J. Streptococcus pneumoniae Biofilm Formation Is Strain Dependent, Multifactorial, and Associated with Reduced Invasiveness and Immunoreactivity during Colonization. mBio 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogaert, D.; de Groot, R.; Hermans, P.W.M. Streptococcus pneumoniae colonisation: The key to pneumococcal disease. Lancet Infect. Dis. 2004, 4, 144–154. [Google Scholar] [CrossRef]
- Simell, B.; Auranen, K.; Kayhty, H.; Goldblatt, D.; Dagan, R.; O’Brien, K.L.; Grp, P.C. The fundamental link between pneumococcal carriage and disease. Expert Rev. Vaccines 2012, 11, 841–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Mello, A.; Riegler, A.N.; Martinez, E.; Beno, S.M.; Ricketts, T.D.; Foxman, E.F.; Orihuela, C.J.; Tettelin, H. An in vivo atlas of host-pathogen transcriptomes during Streptococcus pneumoniae colonization and disease. Proc. Natl. Acad. Sci. USA 2020, 117, 33507–33518. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.H.; Zhang, G.; Nayerhoda, R.; Beitelshees, M.; Hill, A.; Rostami, P.; Li, Y.; Davidson, B.A.; Knight, P., 3rd; Pfeifer, B.A. Comprehensive vaccine design for commensal disease progression. Sci. Adv. 2017, 3, e1701797. [Google Scholar] [CrossRef] [Green Version]
- Pettigrew, M.M.; Marks, L.R.; Kong, Y.; Gent, J.F.; Roche-Hakansson, H.; Hakansson, A.P. Streptococcus pneumoniae and influenza: Dynamic changes in the pneumococcal transcriptome during transition from biofilm formation to invasive disease. Infect. Immun. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shainheit, M.G.; Muie, M.; Camilli, A. The Core Promoter of the Capsule Operon of Streptococcus pneumoniae Is Necessary for Colonization and Invasive Disease. Infect. Immun. 2014, 82, 694–705. [Google Scholar] [CrossRef] [Green Version]
- Dockrell, D.H.; Whyte, M.K.B.; Mitchell, T.J. Pneumococcal pneumonia: Mechanisms of infection and resolution. Chest 2012, 142, 482–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matanock, A.L.G.; Gierke, R.; Kobayashi, M.; Leidner, A.; Pilishvili, T. Use of 13-Valent Pneumococcal Conjugate Vaccine and 23-Valent Pneumococcal Polysaccharide Vaccine Among Adults Aged ≥65 Years: Updated Recommendations of the Advisory Committee on Immunization Practices. 2019; pp. 1069–1075. Available online: https://pubmed.ncbi.nlm.nih.gov/31751323/ (accessed on 1 March 2021).
- Suzuki, M.; Dhoubhadel, B.G.; Ishifuji, T.; Yasunami, M.; Yaegashi, M.; Asoh, N.; Ishida, M.; Hamaguchi, S.; Aoshima, M.; Ariyoshi, K.; et al. Serotype-specific effectiveness of 23-valent pneumococcal polysaccharide vaccine against pneumococcal pneumonia in adults aged 65 years or older: A multicentre, prospective, test-negative design study. Lancet Infect. Dis. 2017, 17, 313–321. [Google Scholar] [CrossRef]
- Bonten, M.J.; Huijts, S.M.; Bolkenbaas, M.; Webber, C.; Patterson, S.; Gault, S.; van Werkhoven, C.H.; van Deursen, A.M.; Sanders, E.A.; Verheij, T.J.; et al. Polysaccharide conjugate vaccine against pneumococcal pneumonia in adults. N. Engl. J. Med. 2015, 372, 1114–1125. [Google Scholar] [CrossRef] [Green Version]
- van Werkhoven, C.H.; Huijts, S.M.; Bolkenbaas, M.; Grobbee, D.E.; Bonten, M.J. The Impact of Age on the Efficacy of 13-valent Pneumococcal Conjugate Vaccine in Elderly. Clin. Infect. Dis. 2015, 61, 1835–1838. [Google Scholar] [CrossRef] [Green Version]
- Bou Ghanem, E.N.; Maung, N.H.T.; Siwapornchai, N.; Goodwin, A.E.; Clark, S.; Munoz-Elias, E.J.; Camilli, A.; Gerstein, R.M.; Leong, J.M. Nasopharyngeal Exposure to Streptococcus pneumoniae Induces Extended Age-Dependent Protection against Pulmonary Infection Mediated by Antibodies and CD138(+) Cells. J. Immunol. 2018, 200, 3739–3751. [Google Scholar] [CrossRef]
- Sen, G.; Chen, Q.; Snapper, C.M. Immunization of aged mice with a pneumococcal conjugate vaccine combined with an unmethylated CpG-containing oligodeoxynucleotide restores defective immunoglobulin G antipolysaccharide responses and specific CD4+-T-cell priming to young adult levels. Infect. Immun. 2006, 74, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- Krone, C.L.; van de Groep, K.; Trzcinski, K.; Sanders, E.A.; Bogaert, D. Immunosenescence and pneumococcal disease: An imbalance in host-pathogen interactions. Lancet Respir. Med. 2014, 2, 141–153. [Google Scholar] [CrossRef]
- Bhalla, M.; Simmons, S.R.; Abamonte, A.; Herring, S.E.; Roggensack, S.E.; Bou Ghanem, E.N. Extracellular adenosine signaling reverses the age-driven decline in the ability of neutrophils to kill Streptococcus pneumoniae. Aging Cell 2020, 19, e13218. [Google Scholar] [CrossRef] [PubMed]
- Simmons, S.R.; Bhalla, M.; Herring, S.E.; Tchalla, E.Y.I.; Bou Ghanem, E.N. Older but Not Wiser: The Age-Driven Changes in Neutrophil Responses during Pulmonary Infections. Infect. Immun. 2021, 89. [Google Scholar] [CrossRef]
- Meyer, K.C. The role of immunity in susceptibility to respiratory infection in the aging lung. Respir. Physiol. 2001, 128, 23–31. [Google Scholar] [CrossRef]
- Park, S.; Nahm, M.H. Older adults have a low capacity to opsonize pneumococci due to low IgM antibody response to pneumococcal vaccinations. Infect. Immun. 2011, 79, 314–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerutti, A.; Puga, I.; Magri, G. The B cell helper side of neutrophils. J. Leukoc. Biol. 2013, 94, 677–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adderson, E.E. Antibody repertoires in infants and adults: Effects of T-independent and T-dependent immunizations. Springer Semin. Immunopathol. 2001, 23, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Ridda, I.; Macintyre, C.R.; Lindley, R.; Gao, Z.; Sullivan, J.S.; Yuan, F.F.; McIntyre, P.B. Immunological responses to pneumococcal vaccine in frail older people. Vaccine 2009, 27, 1628–1636. [Google Scholar] [CrossRef]
- Buffa, S.; Bulati, M.; Pellicano, M.; Dunn-Walters, D.K.; Wu, Y.C.; Candore, G.; Vitello, S.; Caruso, C.; Colonna-Romano, G. B cell immunosenescence: Different features of naive and memory B cells in elderly. Biogerontology 2011, 12, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Bou Ghanem, E.N.; Clark, S.; Du, X.; Wu, D.; Camilli, A.; Leong, J.M.; Meydani, S.N. The alpha-tocopherol form of vitamin E reverses age-associated susceptibility to streptococcus pneumoniae lung infection by modulating pulmonary neutrophil recruitment. J. Immunol. 2015, 194, 1090–1099. [Google Scholar] [CrossRef] [Green Version]
- Simell, B.; Vuorela, A.; Ekstrom, N.; Palmu, A.; Reunanen, A.; Meri, S.; Kayhty, H.; Vakevainen, M. Aging reduces the functionality of anti-pneumococcal antibodies and the killing of Streptococcus pneumoniae by neutrophil phagocytosis. Vaccine 2011, 29, 1929–1934. [Google Scholar] [CrossRef]
- Pinti, M.; Appay, V.; Campisi, J.; Frasca, D.; Fulop, T.; Sauce, D.; Larbi, A.; Weinberger, B.; Cossarizza, A. Aging of the immune system: Focus on inflammation and vaccination. Eur. J. Immunol. 2016, 46, 2286–2301. [Google Scholar] [CrossRef]
- Vinuesa, C.G.; Linterman, M.A.; Yu, D.; MacLennan, I.C. Follicular Helper T Cells. Annu. Rev. Immunol. 2016, 34, 335–368. [Google Scholar] [CrossRef]
- Gustafson, C.E.; Weyand, C.M.; Goronzy, J.J. T follicular helper cell development and functionality in immune ageing. Clin. Sci. 2018, 132, 1925–1935. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Estimates of deaths associated with seasonal influenza—United States, 1976–2007. MMWR Morb. Mortal. Wkly. Rep. 2010, 59, 1057–1062. [Google Scholar]
- McCullers, J.A. Insights into the interaction between influenza virus and pneumococcus. Clin. Microbiol. Rev. 2006, 19, 571–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakaletz, L.O. Viral-bacterial co-infections in the respiratory tract. Curr. Opin. Microbiol. 2017, 35, 30–35. [Google Scholar] [CrossRef]
- Davis, B.M.; Aiello, A.E.; Dawid, S.; Rohani, P.; Shrestha, S.; Foxman, B. Influenza and community-acquired pneumonia interactions: The impact of order and time of infection on population patterns. Am. J. Epidemiol. 2012, 175, 363–367. [Google Scholar] [CrossRef]
- Siegel, S.J.; Roche, A.M.; Weiser, J.N. Influenza promotes pneumococcal growth during coinfection by providing host sialylated substrates as a nutrient source. Cell Host Microbe 2014, 16, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Diavatopoulos, D.A.; Short, K.R.; Price, J.T.; Wilksch, J.J.; Brown, L.E.; Briles, D.E.; Strugnell, R.A.; Wijburg, O.L. Influenza A virus facilitates Streptococcus pneumoniae transmission and disease. FASEB J. 2010, 24, 1789–1798. [Google Scholar] [CrossRef]
- Ballinger, M.N.; Standiford, T.J. Postinfluenza bacterial pneumonia: Host defenses gone awry. J. Interferon Cytokine Res. 2010, 30, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Metzger, D.W.; Sun, K. Immune dysfunction and bacterial coinfections following influenza. J. Immunol. 2013, 191, 2047–2052. [Google Scholar] [CrossRef] [Green Version]
- McCullers, J.A. The co-pathogenesis of influenza viruses with bacteria in the lung. Nat. Rev. Microbiol. 2014, 12, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; McCullers, J.A. Secondary bacterial infections in influenza virus infection pathogenesis. Curr. Top. Microbiol. Immunol. 2014, 385, 327–356. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.B.; Beitelshees, M.; Nayerhoda, R.; Pfeifer, B.A.; Jones, C.H. Engineering a Next-Generation Glycoconjugate-Like Streptococcus pneumoniae Vaccine. ACS Infect. Dis. 2018, 4, 1553–1563. [Google Scholar] [CrossRef] [PubMed]
- Malley, R.; Lipsitch, M.; Stack, A.; Saladino, R.; Fleisher, G.; Pelton, S.; Thompson, C.; Briles, D.; Anderson, P. Intranasal immunization with killed unencapsulated whole cells prevents colonization and invasive disease by capsulated pneumococci. Infect. Immun. 2001, 69, 4870–4873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, N.M.; Wasserman, G.A.; Coleman, F.T.; Hilliard, K.L.; Yamamoto, K.; Lipsitz, E.; Malley, R.; Dooms, H.; Jones, M.R.; Quinton, L.J.; et al. Regionally compartmentalized resident memory T cells mediate naturally acquired protection against pneumococcal pneumonia. Mucosal. Immunol. 2018, 11, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, A.T.; Wasserman, G.A.; Arafa, E.I.; Wooten, A.K.; Smith, N.M.S.; Martin, I.M.C.; Jones, M.R.; Quinton, L.J.; Mizgerd, J.P. Lung CD4(+) resident memory T cells remodel epithelial responses to accelerate neutrophil recruitment during pneumonia. Mucosal Immunol. 2020, 13, 334–343. [Google Scholar] [CrossRef]
- Malley, R.; Trzcinski, K.; Srivastava, A.; Thompson, C.M.; Anderson, P.W.; Lipsitch, M. CD4+ T cells mediate antibody-independent acquired immunity to pneumococcal colonization. Proc. Natl. Acad. Sci. USA 2005, 102, 4848–4853. [Google Scholar] [CrossRef] [Green Version]
- Jochems, S.P.; de Ruiter, K.; Solorzano, C.; Voskamp, A.; Mitsi, E.; Nikolaou, E.; Carniel, B.F.; Pojar, S.; German, E.L.; Reine, J.; et al. Innate and adaptive nasal mucosal immune responses following experimental human pneumococcal colonization. J. Clin. Investig. 2019, 129, 4523–4538. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.K.; Ferreira, D.M.; Gritzfeld, J.F.; Wright, A.D.; Armitage, K.; Jambo, K.C.; Bate, E.; El Batrawy, S.; Collins, A.; Gordon, S.B. Human nasal challenge with Streptococcus pneumoniae is immunising in the absence of carriage. PLoS Pathog. 2012, 8, e1002622. [Google Scholar] [CrossRef]
- Wright, A.K.; Bangert, M.; Gritzfeld, J.F.; Ferreira, D.M.; Jambo, K.C.; Wright, A.D.; Collins, A.M.; Gordon, S.B. Experimental human pneumococcal carriage augments IL-17A-dependent T-cell defence of the lung. PLoS Pathog. 2013, 9, e1003274. [Google Scholar] [CrossRef]
- Mitsi, E.; Carniel, B.; Reine, J.; Rylance, J.; Zaidi, S.; Soares-Schanoski, A.; Connor, V.; Collins, A.M.; Schlitzer, A.; Nikolaou, E.; et al. Nasal Pneumococcal Density Is Associated with Microaspiration and Heightened Human Alveolar Macrophage Responsiveness to Bacterial Pathogens. Am. J. Respir. Crit. Care Med. 2020, 201, 335–347. [Google Scholar] [CrossRef]
- Ferreira, D.M.; Neill, D.R.; Bangert, M.; Gritzfeld, J.F.; Green, N.; Wright, A.K.; Pennington, S.H.; Bricio-Moreno, L.; Moreno, A.T.; Miyaji, E.N.; et al. Controlled human infection and rechallenge with Streptococcus pneumoniae reveals the protective efficacy of carriage in healthy adults. Am. J. Respir. Crit. Care Med. 2013, 187, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, V.M.; Kaneko, K.; Solorzano, C.; MacLoughlin, R.; Saleem, I.; Miyaji, E.N. Progress in mucosal immunization for protection against pneumococcal pneumonia. Expert Rev. Vaccines 2019, 18, 781–792. [Google Scholar] [CrossRef]
- Mutsch, M.; Zhou, W.; Rhodes, P.; Bopp, M.; Chen, R.T.; Linder, T.; Spyr, C.; Steffen, R. Use of the inactivated intranasal influenza vaccine and the risk of Bell’s palsy in Switzerland. N. Engl. J. Med. 2004, 350, 896–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, D.J.; Huo, Z.; Barnett, S.; Kromann, I.; Giemza, R.; Galiza, E.; Woodrow, M.; Thierry-Carstensen, B.; Andersen, P.; Novicki, D.; et al. Transient facial nerve paralysis (Bell’s palsy) following intranasal delivery of a genetically detoxified mutant of Escherichia coli heat labile toxin. PLoS ONE 2009, 4, e6999. [Google Scholar] [CrossRef]
- Zaman, M.; Ozberk, V.; Langshaw, E.L.; McPhun, V.; Powell, J.L.; Phillips, Z.N.; Ho, M.F.; Calcutt, A.; Batzloff, M.R.; Toth, I.; et al. Novel platform technology for modular mucosal vaccine that protects against streptococcus. Sci. Rep. 2016, 6, 39274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ickovic, M.R.; Relyveld, E.H.; Henocq, E.; David, B.; Marie, F.N. Calcium-phosphate-adjuvanted allergens: Total and specific IgE levels before and after immunotherapy with house dust and Dermatophagoides pteronyssinus extracts. Ann. Immunol. 1983, 134D, 385–398. [Google Scholar] [CrossRef]
- He, Q.; Mitchell, A.R.; Johnson, S.L.; Wagner-Bartak, C.; Morcol, T.; Bell, S.J. Calcium phosphate nanoparticle adjuvant. Clin. Diagn. Lab. Immunol. 2000, 7, 899–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudgal, R.; Nehul, S.; Tomar, S. Prospects for mucosal vaccine: Shutting the door on SARS-CoV-2. Hum. Vaccines Immunother. 2020, 16, 2921–2931. [Google Scholar] [CrossRef]
- Sokolova, V.; Knuschke, T.; Kovtun, A.; Buer, J.; Epple, M.; Westendorf, A.M. The use of calcium phosphate nanoparticles encapsulating Toll-like receptor ligands and the antigen hemagglutinin to induce dendritic cell maturation and T cell activation. Biomaterials 2010, 31, 5627–5633. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, X.; Huang, X.; Zhang, J.; Xia, N.; Zhao, Q. Calcium phosphate nanoparticles as a new generation vaccine adjuvant. Expert Rev. Vaccines 2017, 16, 895–906. [Google Scholar] [CrossRef]
- Relyveld, E.H.; Ickovic, M.R.; Henocq, E.; Garcelon, M. Calcium phosphate adjuvanted allergens. Ann. Allergy 1985, 54, 521–529. [Google Scholar] [PubMed]
- Masson, J.D.; Thibaudon, M.; Belec, L.; Crepeaux, G. Calcium phosphate: A substitute for aluminum adjuvants? Expert Rev. Vaccines 2017, 16, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Kodama, S.; Abe, N.; Hirano, T.; Suzuki, M. Safety and efficacy of nasal application of CpG oligodeoxynucleotide as a mucosal adjuvant. Laryngoscope 2006, 116, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkus, F.W.; Verhoef, J.C.; Schipper, N.G.; Marttin, E. Nasal mucociliary clearance as a factor in nasal drug delivery. Adv. Drug Deliv. Rev. 1998, 29, 13–38. [Google Scholar] [CrossRef]
- Beule, A.G. Physiology and pathophysiology of respiratory mucosa of the nose and the paranasal sinuses. GMS Curr. Top. Otorhinolaryngol. Head Neck Surg. 2010, 9, Doc07. [Google Scholar] [CrossRef]
- Shekhar, S.; Khan, R.; Schenck, K.; Petersen, F.C. Intranasal Immunization with the Commensal Streptococcus mitis Confers Protective Immunity against Pneumococcal Lung Infection. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Tada, R.; Suzuki, H.; Takahashi, S.; Negishi, Y.; Kiyono, H.; Kunisawa, J.; Aramaki, Y. Nasal vaccination with pneumococcal surface protein A in combination with cationic liposomes consisting of DOTAP and DC-chol confers antigen-mediated protective immunity against Streptococcus pneumoniae infections in mice. Int. Immunopharmacol. 2018, 61, 385–393. [Google Scholar] [CrossRef]
- Xu, J.H.; Dai, W.J.; Chen, B.; Fan, X.Y. Mucosal immunization with PsaA protein, using chitosan as a delivery system, increases protection against acute otitis media and invasive infection by Streptococcus pneumoniae. Scand. J. Immunol. 2015, 81, 177–185. [Google Scholar] [CrossRef]
- Suzuki, H.; Watari, A.; Hashimoto, E.; Yonemitsu, M.; Kiyono, H.; Yagi, K.; Kondoh, M.; Kunisawa, J. C-Terminal Clostridium perfringens Enterotoxin-Mediated Antigen Delivery for Nasal Pneumococcal Vaccine. PLoS ONE 2015, 10, e0126352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, T.C.; Oliveira, M.L.S.; Soares-Schanoski, A.; Chavez-Rico, S.L.; Figueiredo, D.B.; Goncalves, V.M.; Ferreira, D.M.; Kunda, N.K.; Saleem, I.Y.; Miyaji, E.N. Mucosal immunization with PspA (Pneumococcal surface protein A)-adsorbed nanoparticles targeting the lungs for protection against pneumococcal infection. PLoS ONE 2018, 13, e0191692. [Google Scholar] [CrossRef] [Green Version]
- Salha, D.; Szeto, J.; Myers, L.; Claus, C.; Sheung, A.; Tang, M.; Ljutic, B.; Hanwell, D.; Ogilvie, K.; Ming, M.; et al. Neutralizing antibodies elicited by a novel detoxified pneumolysin derivative, PlyD1, provide protection against both pneumococcal infection and lung injury. Infect. Immun. 2012, 80, 2212–2220. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Q.; Lv, Z.Y.; Gan, H.Q.; Xian, M.; Zhang, K.X.; Mai, J.Y.; Yu, X.B.; Wu, Z.D. Intranasal immunization with autolysin (LytA) in mice model induced protection against five prevalent Streptococcus pneumoniae serotypes in China. Immunol. Res. 2011, 51, 108–115. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Kim, S.Y.; Kim, M.S.; Lee, S.E.; Rhee, J.H. Intranasal immunization with recombinant PspA fused with a flagellin enhances cross-protective immunity against Streptococcus pneumoniae infection in mice. Vaccine 2011, 29, 5731–5739. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Yao, R.; Wang, H.; Pang, D.; Liu, Y.; Xu, H.; Zhang, S.; Zhang, X.; Yin, Y. Mucosal and systemic immunization with a novel attenuated pneumococcal vaccine candidate confer serotype independent protection against Streptococcus pneumoniae in mice. Vaccine 2014, 32, 4179–4188. [Google Scholar] [CrossRef] [PubMed]
- Jwa, M.Y.; Jeong, S.; Ko, E.B.; Kim, A.R.; Kim, H.Y.; Kim, S.K.; Seo, H.S.; Yun, C.H.; Han, S.H. Gamma-irradiation of Streptococcus pneumoniae for the use as an immunogenic whole cell vaccine. J. Microbiol. 2018, 56, 579–585. [Google Scholar] [CrossRef]
- Babb, R.; Chen, A.; Hirst, T.R.; Kara, E.E.; McColl, S.R.; Ogunniyi, A.D.; Paton, J.C.; Alsharifi, M. Intranasal vaccination with gamma-irradiated Streptococcus pneumoniae whole-cell vaccine provides serotype-independent protection mediated by B-cells and innate IL-17 responses. Clin. Sci. 2016, 130, 697–710. [Google Scholar] [CrossRef] [Green Version]


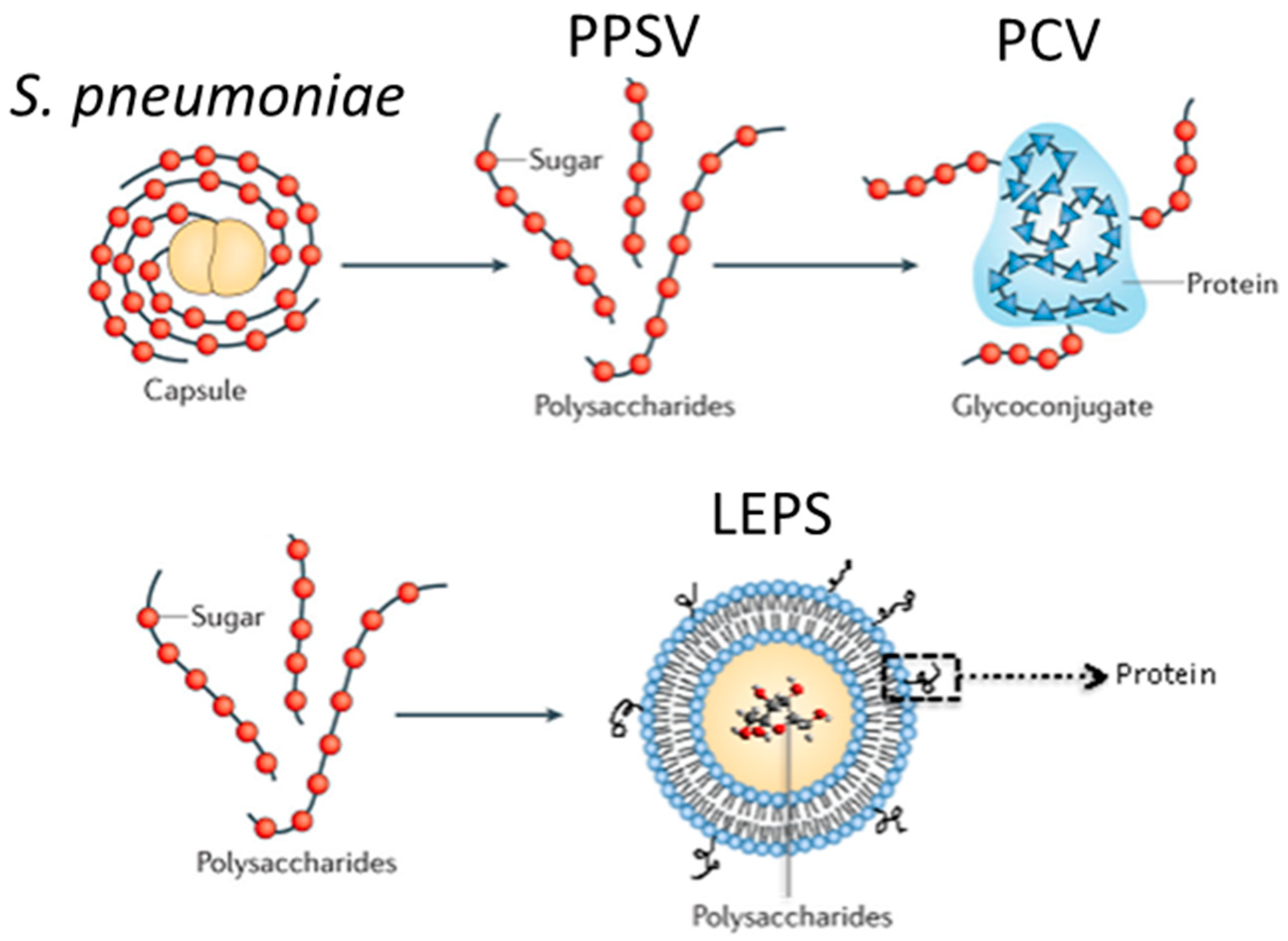{kind=link}
| Disease | Infectious Agent | Notes | Therapeutic/Preventative Options | References |
|---|---|---|---|---|
| Influenza | Primarily Influenza A virus |
|
| [17] |
| Pneumococcal Disease | Streptococcus pneumoniae (bacteria) |
|
| [18] |
| Whooping Cough (Pertussis) | Bordetella pertussis (bacteria) |
|
| [19] |
| Tuberculosis | Mycobacterium tuberculosis (bacteria) |
|
| [20] |
| Coronavirus-based diseases | Various viruses (including SARS-CoV-2) |
|
| [21,22] |
| Disease | Number of IN Applications (Identified Using the PubMed Search Engine for Indicated Year) | ||||
|---|---|---|---|---|---|
| 2009 | 2014 | 2019 | 2020 | 2021 | |
| Influenza | 14 | 23 | 17 | 6 | 2 |
| Pneumococcal Disease | 4 | 2 | 9 | 3 | 2 |
| Whooping Cough | 1 | 3 | 0 | 3 | 1 |
| Tuberculosis | 4 | 6 | 3 | 3 | 3 |
| COVID-19 | N/A | N/A | N/A | 6 | 6 |
| IN Method Used | Rational | Times Tested | Disease Application | Reference |
|---|---|---|---|---|
| Dry Powder -Inulin nanoparticles -Chitosan nanospheres |
| 2 | Influenza (Influenza A virus) | [43,44] |
| Intranasal Instillation -Stabilized protein subunit -Nanoparticles -Inert bacterial spores -Bacterium-like particles |
| 7 | Pneumonia (Actinobacillus pleuropneumoniae) | [45] |
| Tuberculosis (M. tuberculosis) | [46,47,48] | |||
| Pneumococcal Disease (S. pneumoniae) | [49] | |||
| Influenza (Influenza A virus) | [50,51] | |||
| Aerosolization -Adenovirus vector-based |
| 1 | Influenza (Influenza A virus) | [52] |
| Nasal Gel -Cationic cholesteryl pullulan |
| 1 | Pneumococcal Disease (S. pneumoniae) | [53] |
| Vaccine Category | Antigen Content (IN Method) | Adjuvant Included | Reference |
|---|---|---|---|
| Subunit Vaccine | S. pneumoniae serotype 4 capsule (Intranasal Instillation) | No adjuvant | [147] |
| Recombinant PspA (Nasal Gel) | No adjuvant | [53] | |
| Recombinant PspA (Intranasal Instillation) | DOTAP/DC-chol liposome | [148] | |
| Recombinant PspA (Intranasal Instillation) | Chitosan | [149] | |
| Recombinant PspA (Intranasal Instillation) | Clostridium perfringens enterotoxin (C-CPE) | [150] | |
| Recombinant PspA/BLP (Intranasal Instillation) | No adjuvant | [49] | |
| Recombinant PspA (nanoparticle absorbed) (Intranasal Instillation) | (PGA-co-PDL) nanoparticles | [151] | |
| Recombinant PLY (Intranasal Instillation) | Aluminum hydroxide | [152] | |
| Recombinant LytA (Intranasal Instillation) | CpG oligodeoxynucleotides | [153] | |
| Recombinant PspA/FlaB fusion (Intranasal Instillation) | Recombinant FlaB | [154] | |
| Live and/or Attenuated | S. pneumoniae Y1 (Intranasal Instillation) | Cholera Toxin | [155] |
| S. pneumoniae serotype 19F/23A/35B (Intranasal Instillation) | No adjuvant | [124] | |
| S. pneumoniae serotype 19F (Intranasal Instillation) | No adjuvant | [125] | |
| S. pneumoniae serotype 23F (Intranasal Instillation) | No adjuvant | [129] | |
| S. pneumoniae serotype 6B (Intranasal instillation) | No adjuvant | [130] | |
| S. pneumoniae serotype 6B (Intranasal instillation) | No adjuvant | [131] | |
| Inactivated/Killed | Gamma irradiated nonencapsulated S. pneumoniae TIGR4 (Intranasal Instillation) | Cholera toxin | [156] |
| Gamma irradiated whole cell S. pneumoniae (Intranasal Instillation) | No adjuvant | [157] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walkowski, W.; Bassett, J.; Bhalla, M.; Pfeifer, B.A.; Ghanem, E.N.B. Intranasal Vaccine Delivery Technology for Respiratory Tract Disease Application with a Special Emphasis on Pneumococcal Disease. Vaccines 2021, 9, 589. https://doi.org/10.3390/vaccines9060589
Walkowski W, Bassett J, Bhalla M, Pfeifer BA, Ghanem ENB. Intranasal Vaccine Delivery Technology for Respiratory Tract Disease Application with a Special Emphasis on Pneumococcal Disease. Vaccines. 2021; 9(6):589. https://doi.org/10.3390/vaccines9060589
Chicago/Turabian StyleWalkowski, William, Justin Bassett, Manmeet Bhalla, Blaine A. Pfeifer, and Elsa N. Bou Ghanem. 2021. "Intranasal Vaccine Delivery Technology for Respiratory Tract Disease Application with a Special Emphasis on Pneumococcal Disease" Vaccines 9, no. 6: 589. https://doi.org/10.3390/vaccines9060589
APA StyleWalkowski, W., Bassett, J., Bhalla, M., Pfeifer, B. A., & Ghanem, E. N. B. (2021). Intranasal Vaccine Delivery Technology for Respiratory Tract Disease Application with a Special Emphasis on Pneumococcal Disease. Vaccines, 9(6), 589. https://doi.org/10.3390/vaccines9060589