1. Introduction
The HIV-1 Envelope surface glycoprotein (Env) is a trimer of heterodimers composed of non-covalently associated gp120 and gp41 subunits, which mature from a gp160 precursor unpon enzymatic cleavage by cellular proteases. Env mediates binding to the CD4 receptor on immune cells and, upon a series of conformational changes, facilitates infection (as reviewed by [
1]). Antibodies targeting Env in its native conformation can prevent binding to the CD4 receptor or the CCR5 or CXCR4 co-receptor on target cells, or interfere with the fusion process, and thereby prevent infection. However, the induction of neutralizing antibodies (nAbs) is hampered by the dense glycan shield on Env, which covers almost the entire protein surface and thereby restricts accessibility for antibodies [
2]. This, together with the structural flexibility and high degree of sequence variability of Env (up to 35% across all group M clades [
3]) are regarded as the main impediments to the design of a successful vaccine capable of inducing antibodies that can neutralize a broad variety of HIV-1 strains (broadly neutralizing antibodies, bnAbs) (as reviewed by [
4,
5] and others). Therefore, Env immunogen candidates that are stabilized in a native, pre-fusion closed conformation are desirable, and centralized immunogen sequences [
3,
6] that represent circulating virus isolates as closely as possible would be additionally beneficial for the development of breadth.
Over the past two decades, HIV-1 Env trimer-based immunogens have undergone fundamental improvements. The non-covalent association of the gp120 and gp41 subunits, which lead to gp120 dissociation and shedding of gp120, particularly on soluble forms of Env, was overcome by two strategies: (i) The introduction of a disulfide bridge between gp120 and gp41 (A501C/T605C) in combination with the I559P substitution in gp41 prevented shedding of gp120 and stabilized the trimer [
7,
8]. (ii) The replacement of the furin cleavage site by a flexible linker of suitable length allowed cleavage-independent trimers [
9]. These strategies contributed to the generation of closed, native-like trimers with reduced exposure of epitopes that induce non-neutralizing antibodies [
10]. This is of particular importance, as B-cell responses to off-target immunodominant regions such as V3 (as reviewed by [
11]) might compete with other subdominant B-cells, recognizing weakly immunogenic neutralization-relevant regions in the germinal centers [
12]. Today, a variety of native-like trimers from different viral clades are available [
13,
14,
15,
16,
17] and continuous optimization of stabilized Env trimers for favorable immunological traits is ongoing (as reviewed by [
18]). These immunogens, however, were derived from distinct viral isolates and are therefore likely to induce strain-specific responses with low cross-reactivity.
Induction of more broadly reactive antibody responses might be promoted by genetically and antigenically ‘centralized’ immunogens such as consensus, ancestor, or mosaic proteins, which aim at representing the diversity of circulating viruses. Mosaic immunogens, being enriched for T-cell or B-cell epitopes, have demonstrated their ability to induce improved cellular immune responses [
19]. While ancestral sequence reconstruction infers phylogenetic relationships, the generation of a consensus sequence utilizes the most frequently observed amino acids in each position of a multiple sequence alignment and therefore provides a sequence most closely related to all input sequences [
20]. Notably, an ancestral clade C SOSIP gp140 trimer has proven successful to induce broadly neutralizing nanobodies against HIV-1 in camels [
21]. Currently, only a small number of native-like Env trimer constructs derived from consensus sequences are being explored. A soluble stabilized Env trimer was generated with a consensus sequence of clade C [
22] and the underlying repair-and-stabilization concept (RnS) was successfully transferred to representative isolates from other clades [
23]. Neither study, however, assessed the immunogenicity of these trimers. Further, the Env trimer ConM SOSIP, based on a consensus sequence covering the complete group M, induced neutralizing antibody responses in rabbits and macaques [
24], and is currently being tested in a clinical trial in humans (NCT03816137) [
25].
The only phase 3 clinical trial so far to show moderate efficacy of 31% in preventing HIV-1 infection after 3 years was the RV144 trial in Thailand [
26]. Several immunological correlates of protection were identified [
27], which include not only neutralizing antibodies but for example also binding antibodies targeting the V1V2 region in Env. The protective effects of RV144, however, could so far be transferred neither to other populations nor to different immunogens. Therefore, there is still an urgent need to explore new potential immunogens and immunization schedules.
Here, we present a set of Env variants derived from a novel clade C consensus sequence (ConC). Viruses from HIV-1 clade C account for almost one half of global infections and are furthermore overrepresented in the high prevalence areas of sub-Saharan Africa and south-east Asia [
28]. Regrettably, sub-optimal ART coverage is also reported for these regions [
29]. The panel of Env proteins we have generated consists of one gp120 monomer and four trimers that were generated by stepwise inclusion of amino acid substitutions with known impact on the protein’s propensity to form pre-fusion closed trimers. On top, selected N-linked glycan sites were modified in order to facilitate binding of bnAb 2G12 for affinity purification (KI; glycan knock-in) [
30,
31] and in order to allow for improved accessibility of the CD 4bindig site (KO; glycan knock-out) [
32]. We systematically analyzed the positive impact of the stepwise modification of ConC KIKO Env variants on biochemical and biophysical protein traits and on antigenicity using a panel of well-characterized antibodies targeting the major sites of vulnerability of Env. In addition, we demonstrate that the favorable in vitro characteristics translate into a beneficial immunological outcome in immunized Balb/c mice.
2. Materials and Methods
2.1. Construct Design
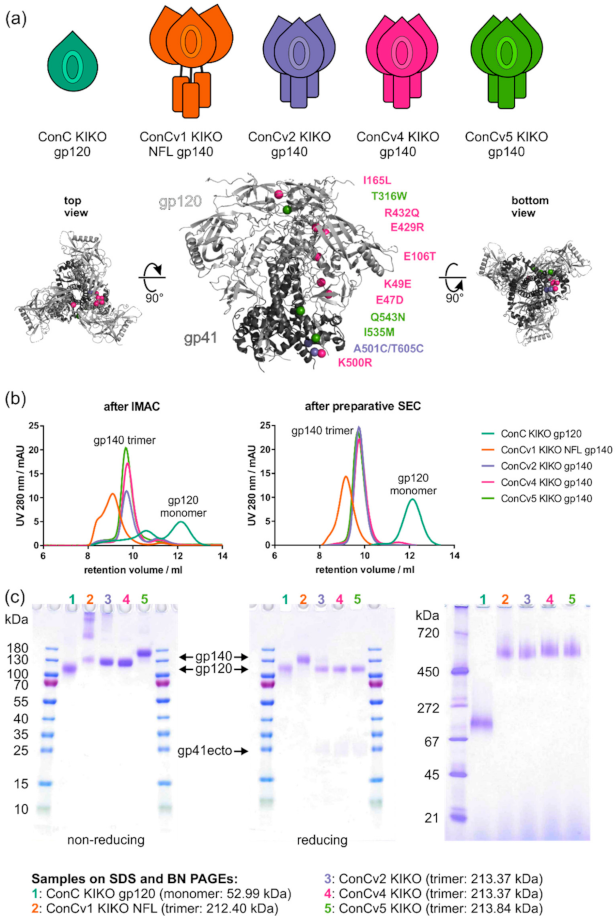
The HIV-1 clade C consensus sequence (ConC) was derived from a multiple-sequence alignment from all available sequences from Los Alamos Sequence Database (cut-off date: 1 April 2016), with restriction to one sequence per patient, using the default method of the platform’s Search Interface. The gp120 monomer and the gp140 trimer constructs were truncated at HXB2 (GeneBank Accession: K03455) positions 507 and 664, respectively, and the autologous signal peptide was replaced by the signal peptide from the tPA (tissue plasminogen activator) serine protease [
33]. The N-linked glycan at position 295 (V295N substation) was introduced to restore binding to antibody 2G12 [
30,
31], and the N-linked glycans in positions 276 (N276Q substitution) and 462/463 (tandem sequon, N463Q/T464A substitutions) were removed by substitution of asparagine to glutamine for better accessibility of the CD4 binding site (CD4bs) in an immune focusing approach [
32]. The resulting constructs were termed KIKO (KI: knock-in of 2G12 relevant N-linked glycans; KO: knock-out of N-linked glycans around the CD4bs). The different ConC trimer versions ConCv1, v2, v4, and v5 were generated essentially as follows: Briefly, for ConCv1 KIKO NFL, only the cleavage site REKR was replaced by (G
4S)
2, a glycine-serine based native flexible linker (NFL), in order to obtain cleavage independent trimers where the gp120 and gp41 subunits were covalently linked [
9]. For ConCv2 KIKO, the SOSIP mutations consisting of a disulfide bridge A501C/T605C between gp120 and gp41 and an I559P substitution in gp41 (for pre-fusion state stabilization) were included [
7,
8]. In addition, for ConCv4 KIKO, a set of eight amino acid substitutions (E47D, K49E, V65K, E106T, I165L, E429R, R432Q, and K500R) termed TD8 [
34] and corresponding amino acid substitutions in spatial proximity (unpublished) were introduced. Furthermore, for ConCv5 KIKO, an additional group of six amino acid substitutions for inhibition of transition to the CD4i state (H66R/T316W) and for enhanced trimer formation (I535M/Q543N) [
35] as well as for additional covalent linkage between gp120 and gp41 (disulfide bond A73C-A561C) [
36] were introduced.
2.2. Expression and Purification of Recombinant Env Proteins
Plasmids encoding for the env genes of Env variants AMC008 SOSIPv4.2 and AMC011 SOSIPv4.2 were kindly provided by Rogier W. Sanders (AMC, Amsterdam, The Netherlands). All used Envelope variants had a C-terminal hexa-histidine tag (His6) for purification that was directly fused to the Env proteins via a glycine-serine-linker (GS). Briefly, FreeStyle™ 293-F cells (Thermo Fisher Scientific, Waltham, MA, USA) cells were cultivated in FreeStyle™ 293 Expression Medium (Thermo Fisher Scientific, Waltham, MA, USA) and transfected with 1 µg DNA/mL at a cell density of 1 × 106 cells/mL with plasmids encoding for the env and furin protease genes at a 1:3 ratio using polyethylenimine (PEI; Polysciences Inc., Warrington, PA, USA). In the case of variants that did not require cleavage by the furin protease (ConC KIKO gp120, ConCv1 KIKO NFL and 96ZM651 REKS.664), only plasmid DNA encoding for the respective env genes was used. Supernatants were harvested 5 days after transfection. Env proteins were purified by immobilized metal ion affinity chromatography (IMAC) using HisTrap excel columns (Cytiva, Marlborough, MA, USA) according to the manufacturer’s recommendations followed by buffer exchange to PBS. Proteins were subsequently further purified to homogeneity by size exclusion chromatography (SEC) using a Superdex® 200 Increase 10/300 GL Column (Cytiva, Marlborough, MA, USA). The fractions representing the main peak were analyzed by BN PAGE (Blue Native Polyacrylamide Gel Electrophoresis). Suitable fractions were pooled for further use. Analytical SEC runs were performed with 10 µg of protein being loaded on a Superdex® 200 Increase 10/300 GL Column (Cytiva, Marlborough, MA, USA) at a concentration of 0.1 µg/mL via a 100 µL sample loop overloaded with a suitable volume.
2.3. Monoclonal Antibodies
Antibodies 5F3 and 2G12 were purchased from Polymun Scientific, Austria. Antibodies and plasmids for antibodies PGT145 and PGT151 were kindly provided by Dennis Burton (The Scripps Research Institute, Torrey Pines Rd, La Jolla, CA, USA). Antibody and plasmids for antibody 10-1074 were kindly provided by Michel Nussenzweig (The Rockefeller University, New York, NY, USA). Plasmids for antibodies 14E and 19b were kindly provided by James Robinson (CHAVI-ID, USA). The following reagents were obtained through the NIH HIV Reagent Program, Division of AIDS, NIAID, NIH: Anti-Human Immunodeficiency Virus 1 gp120 Monoclonal Antibody (PG9), ARP-11557; Anti-Human Immunodeficiency Virus (HIV)-1 gp120 Monoclonal Antibody (PG16), ARP-12150; and Anti-Human Immunodeficiency Virus (HIV)-1 gp120 Monoclonal Antibody (PGT121), ARP-12343, were contributed by International AIDS Vaccine Initiative. Human Immunodeficiency Virus 1 (HIV-1) VRC01 Monoclonal Antibody Heavy Chain Expression Vector, ARP-12035 and Human Immunodeficiency Virus 1 (HIV-1) VRC01 Monoclonal Antibody Light Chain Expression Vector, ARP-12036, were contributed by John Mascola. Anti-Human Immunodeficiency Virus (HIV)-1 gp120 Monoclonal Antibody (F105), ARP-857, was contributed by Dr. Marshall Posner and Dr. Lisa Cavacini. Human Immunodeficiency Virus 1 (HIV-1) 447-52D mAb Heavy Chain Expression Vector, ARP-13617 and Human Immunodeficiency Virus 1 (HIV-1) 447-52D mAb Light Chain Expression Vector, ARP-13618 were contributed by Dr. Susan Zolla-Pazner. Monoclonal Anti-Human Immunodeficiency Virus Type 1 (HIV-1) gp120 Protein, Clone 17b (produced in vitro), ARP-4091, was contributed by Dr. James E. Robinson; Plasmid pcDNA3.1(-) Expressing Human CD4 Fused with Human IgG1 Hinge and Fc Regions, ARP-12960, was contributed by Dr. Xueling Wu. For in-house production of monoclonal human anti-Env antibodies, plasmids encoding for the respective heavy and light chain genes were transfected to either FreeStyle™ 293-F cells or Expi293F™ cells (Thermo Fisher Scientific, Waltham, MA, USA) at a 1:1 ratio. Transfection of FreeStyle™ 293-F cells was performed essentially as described above. Expi293F™ cells were cultivated in Expi293™ Expression Medium (Thermo Fisher Scientific, Waltham, MA, USA) and transfected at a density of 2.9 × 106 cells/mL using the ExpiFectamine™ 293 Transfection Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s recommendations. In both cases, supernatants were harvested 5 days post transfection and purified by Protein A affinity chromatography using HiTrap rProtein A FF columns (Cytiva, Marlborough, MA, USA).
2.4. Blue Native PAGE
Blue Native PAGE (BN PAGE) was performed for verification of the purity of the trimer pools. Therefore, 2 µg protein per lane was loaded on a SERVAGel™ N 4–16 (SERVA Electrophoresis GmbH, Heidelberg, Germany) and run for 20 min at 50 V followed by 4 h at 100 V. Gels were stained for 10–20 min with Coomassie staining solution (1.25% (w/v) Coomassie Brilliant Blue R-250, 50% (v/v) ethanol, 7% (v/v) glacial acetic acid) and destained with 7% (v/v) acetic acid overnight.
2.5. SDS PAGE
Reducing and non-reducing SDS-PAGEs were performed for verification of complete cleavage of the gp120 and gp41 subunits (where applicable). Briefly, 2 µg of protein were denatured for 5 min at 95 °C, samples were loaded on pre-cast SERVAGel™ TG PRiME™ 8–16% gels (SERVA Electrophoresis GmbH, Heidelberg, Germany) and run at constant voltage for 10 min at 100 V followed by 45 min at 250 V. Staining and destaining of the gels were performed as described for BN PAGEs.
2.6. Animal Study
Eight-week old Balb/c females were ordered from Charles River Laboratories, UK and housed at University Biomedical Services, University of Cambridge. Animal work was carried out under project license P8143424B with approval from the Animal Welfare Ethical Review Body, University of Cambridge.
Mice were immunized with 20 µg of Env trimers adjuvanted with monophosphoryl lipid A liposome solution (Polymun Scientific, Klosterneuburg, Austria), a Toll-like receptor 4 agonist, at a 1:2.5 ratio in eight groups. Immunizations were administered in 100 µL subcutaneously in the rear flank. Animals received a total of three immunizations at weeks 0, 4, and 8, and sera were collected at the pre-immune state and 2 weeks after each immunization (weeks 0, 2, 6, and 10). Animals were terminally bled at week 10 by cardiac puncture under non-recovery anesthesia.
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
Binding affinities of monoclonal antibodies to various Env trimer variants were determined by Ni-NTA capture ELISA. Briefly, 350 ng Env gp140 trimer or the triple molar amount of gp120 monomer (261 ng) was coated onto Ni-NTA HisSorb plates (Qiagen, Hilden, Germany) over night at 4 °C. Plates were washed with TBS and serial dilutions of the monoclonal antibodies were added in PBS containing 2% skimmed milk and incubated for 2 h at room temperature. After washing with TBS, the detection antibody (rabbit anti-human IgG/HRP, P0214, Agilent, CA, USA) was added at a 1:5000 dilution in PBS containing 1% BSA and incubated for 1 h. After washing with TBS, plates were developed using in-house TMB substrate solution (1:20 dilution of substrate (1 mM TMB, 80 mM H2O2, 10% (v/v) acetone, 90% (v/v) ethanol) in dilution buffer (30 mM tripotassium citrate)) and the reaction was stopped with 1 M sulfuric acid. The absorbance was measured at 450 nm (specific signal) and 655 nm (background) on an iMark™ Microplate Reader (Biorad, Hercules, CA, USA). Data were analyzed using GraphPad Prism Software (GraphPad Software Inc., San Diego, CA, USA).
2.8. Env Binding IgG antibody Responses in Mouse Sera
Binding antibody titers in sera of mice immunized with different Env variants were determined by capture ELISA. Briefly, 350 ng of Env gp140 trimers or 261 ng of Env gp120 monomer were coated onto Ni-NTA HisSorb plates (Qiagen, Hilden, Germany) for 2 h at room temperature or overnight at 4 °C. Plates were washed with TBS and blocked for 30 min with PBS containing 0.1% (w/v) I-BlockTM (Invitrogen, Waltham, MA, USA) and 1x Roti®Block (Carl Roth, Karlsruhe, Germany) (blocking buffer). Plates were washed with TBS and serial dilutions of mouse sera starting at a 1:50 dilution were added in blocking buffer and incubated for 2 h at room temperature. After washing with TBS, the detection antibody (goat anti-mouse IgG/HRP; Jackson ImmunoResearch, West Grove, PA, USA) was added at a 1:10,000 dilution in PBS containing 1% (w/v) BSA and incubated for 1 h at room temperature. Plates were washed with TBS supplemented with 0.05% (v/v) Tween and again with TBS in order to remove residual Tween. Plates were developed as described for monoclonal antibodies. Midpoint titers were calculated using GraphPad Prism Software (GraphPad Software Inc, San Diego, CA, USA). Absence of Env reactivity in the pre-immune sera was confirmed for each animal towards the respective immunogen.
2.9. V1V2 Responses
The following reagents were obtained through the NIH HIV Reagent Program, Division of AIDS, NIAID, NIH: Human Immunodeficiency Virus 1 (HIV-1) Env V1-V2 Recombinant Protein (AE.A244 V1-V2.tags), ARP-12567 and (C.1086 V1-V2.tags), ARP-12568, contributed by Drs. Barton F. Haynes and Hua-Xin Liao. For analysis of V1V2 reactive antibodies, 70 ng of Env V1V2 recombinant protein was coated onto Ni-NTA HisSorb plates (Qiagen, Hilden, Germany) and ELISA analysis was performed as described for Env reactive antibody responses. AUC (area under curve) analysis was carried out using GraphPad Prism Software (GraphPad Software Inc., San Diego, CA, USA).
2.10. Luminex Binding Antibody Multiplex Assay
Breadth of serum reactivity was assessed towards a panel of Env variants including trimers from several viral clades (clade A: BG505 SOSIP.664, clade B: AMC008 SOSIPv4.2, AMC011 SOSIPv4.2, clade C: ConCv1 KIKO NFL, ConCv2 KIKO, ConCv4 KIKO, ConCv5 KIKO, 16055 SOSIP, 96ZM651 REKS.664) and one clade C gp120 monomer (ConC KIKO gp120). MagPlexAvidin Microspheres (2000 beads/test; Luminex, Austin, TX, USA) were washed with PBS, 2 mM EDTA, 1% BSA, 0.05% Tween-20, 0.1% ProClinTM 300 (Sigma Aldrich, St. Louis, MO, USA) (dilution buffer) and then coated at 250 beads/µL with in-house biotinylated anti-His6 IgY (Thermo Fisher Scientific, Waltham, MA, USA) at 5 µg/mL in dilution buffer for 1 h at RT. After washing with dilution buffer, the trimeric antigens were captured at 15 µg/mL and the monomeric gp120 was captured at an equimolar amount over night at 4 °C. After washing with dilution buffer, beads were further washed with PBS containing 0.1% (w/v) I-BlockTM (Invitrogen, Waltham, MA, USA) and 1x Roti®Block (Carl Roth, Karlsruhe, Germany) (blocking buffer) and then blocked for 1 h at RT. After blocking, the different bead regions were pooled and distributed in a 96-well plate (Greiner, Kremsmünster, Austria) and sera were applied in blocking buffer at a 1:80 dilution in a volume of 50 µL and incubated for 2 h at RT. After incubation, beads were washed with blocking buffer and again with dilution buffer. Then a R-Phycoerythrin-conjugated donkey anti-mouse IgG (H + L) AffiniPure F(ab’)2 fragment (Jackson ImmunoResearch, West Grove, PA, USA) was added at a 1:200 dilution for detection and incubated for 1 h at RT. After washing, beads were resuspended in dilution buffer and measurement was performed with a MAGPIX® device (Luminex, Austin, TX, USA). A response was defined as being positive if the specific, background corrected, signal for an individual animal was higher than (1) two times the mean signal of non-immunized animals and (2) the mean plus three times the standard deviation of sera from non-immunized animals.
2.11. Nano Differential Scanning Fluorimetry (nanoDSF)
Thermostability of the trimers was characterized by thermal unfolding with a Prometheus NT.48 nanoDSF (NanoTemper Technologies GmbH, Munich, Germany). Proteins were diluted to a final concentration of 0.05 mg/mL. After loading the samples to High Sensitivity capillaries (NanoTemper Technologies GmbH, Munich Germany), intrinsic fluorescence was measured at a ramp rate of 1 °C/min with an excitation power of 30%. Protein unfolding was monitored by the changes in fluorescence emission at 350 and 330 nm. The thermal unfolding midpoint (Tm) of the proteins was determined with Prometheus NT software.
2.12. Dynamic Light Scattering (DLS)
Dynamic Light Scattering was used for isothermal particle sizing. Briefly, proteins were diluted to a final concentration of 2.5 µM and loaded to High Sensitivity capillaries (NanoTemper Technologies GmbH, Munich, Germany). Measurements were carried out at 20 °C in triplicates of 10 acquisitions each with a Prometheus Panta device (NanoTemper Technologies GmbH, Munich, Germany). The hydrodynamic radius of the particles in solvated state and the polydispersity index (PDI) of the samples were determined with the associated software.
2.13. Negative Staining Electron Microscopy and Image Processing
HIV Env proteins were visualized by negative-stain electron microscopy (EM) using 3–4 µL aliquots at concentrations of 0.1–0.2 mg/mL. Samples were applied for 10 s onto a mica carbon film and transferred to 400-mesh Cu grid that had been glow discharged at 20 mA for 30 s and then negatively stained with 2% (wt/vol) Sodium silicotungstate (SST) for 30 s. Previous to data collection, the grids were screened to assess stain quality and particle distribution. Data were collected on an FEI Tecnai T12 LaB6-EM operating at 120 kV accelerating voltage with a pixel size of 2.8 Å on the specimen plane using a Gatan Orius 1000 CCD Camera. On average, 30–40 micrographs were collected per sample. Classification of closed and open trimers was performed as described [
16]. Briefly, two-dimensional (2D) class averaging was performed with the software Relion [
37], using 35,829 particles for the analysis of ConCv2 KIKO; 56,407 particles for ConCv4 KIKO; and 55,248 particles for ConCv5 KIKO. The 2-D classes were then segregated into three structural groups, closed or open-native particles and non-native particles.
2.14. Peptide Microarray Design
Custom peptide microarrays were designed to systematically map and compare IgG recognition of linear HIV-1 Env regions in preclinical vaccine studies of the EHVA consortium and consisted of 15-mer peptides overlapping by 11 amino acids. Envelope sequences matching EHVA vaccine candidates were therefore included in the array design. These include: 96ZM651_AF286224, BG505_DQ208458, ConC, HKM3, ngp41CM, CN54gp140_AF286226, MVA-CMDR_AFJ93253, and unpublished. In addition to these, multiple other peptide variants were included for 15 previously identified immunodominant regions. These were identified in studies on different vaccine trials and natural HIV infection conducted by us and others [
38,
39,
40,
41,
42,
43]. Peptide variants covering these immunodominant regions in more depth with respect to the antigenic variation were selected as follows: All existing pre-seroconversion (n = 913) and recent (n = 723) HIV infection sequences from 192 subjects collected within 2010–2018 were obtained from the HIV database (
www.hiv.lanl.gov (accessed on 29 May 2018)). These sequences were interrogated to identify the most frequently occurring molecular forms for the 15 selected Env regions. Moreover, peptide variants frequently identified in previous studies ([
38,
39,
40,
41,
42,
43] and unpublished data) are included to bridge newly generated data to results from these earlier studies. Peptides included in the microarray are representative of all HIV-1 clades, however, due to the focus on EHVA immunogens, Clade C is overrepresented. In total, 1034 overlapping 15-mer peptides cover the gp160 extracellular domains of the selected back-bone immunogen sequences and, in addition, frequently occurring antigenic variants within 15 selected immunodominant regions of specific interest. Each peptide is printed on the array in triplicates.
2.15. Peptide Microarray Mapping of HIV Env-Specific IgG Responses in Vaccinated Mice
Mouse sera were analyzed for the presence of HIV-1 Env-specific IgG antibodies as described before [
40] with minor modifications. After blocking of the peptide microarray slides with blocking buffer (0.1% I-Block Protein-Based Blocking Reagent, Thermo Fisher Scientific, Waltham, MA, USA and 1 × ROTI Block, Carl Roth, Karlsruhe, Germany, in PBS), mouse sera were diluted 1:250 in blocking buffer and incubated for 2 h at RT. Bound mouse IgG antibodies were then detected using a cross-adsorbed goat anti-mouse IgG DyLight 650 antibody (Thermo Fisher Scientific, Waltham, MA, USA). Baseline (w0) and post-vaccination (w10) sera of each mouse were processed in the same run, with the baseline samples used for background determination. Microarrays were scanned using a GenePix 4000A scanner at 650 (signal) and 532 nm (background). Resulting images were analyzed using the GenePix Pro 6.0 software (Molecular Devices, San José, CA, USA) by overlaying the array layout encoded in an array-specific grid (gal file). Layout positioning was then controlled manually for accuracy. Results were exported from GenePix Pro 6.0 as gpr files, which link each position on the array with the measured fluorescence intensity (FI) of bound secondary antibody. These were processed using R to calculate the mean FI of the triplicate peptides, which excludes outliers. These resulting mean FI values, linked with each 15-mer peptide sequence, are then assigned to the alignment of all sequences included in the microarray contained in a single fasta file and base-line (w0) values are subtracted from post-vaccination (w10) values. FI values for each amino acid position in the Env alignment are then calculated from FI values of overlapping peptides. To calculate the frequency of responders (FOR) or mean FI per vaccination group, the variant with the strongest response for each position within the backbone alignment was used. IgG responses against individual amino acid positions were considered positive, if the corresponding mean FI value was above 200 after subtraction of the pre-vaccination value. Mean FI values of all mice in one group were calculated, if at least 2 out of the 6 mice showed a positive response against the corresponding array position. Immunodominant antigenic regions (IDRs) were defined as being recognized by at least 60% of mice per immunogen group at the post-vaccination time point and with a mean FI for individual amino acids above 10,512.17 (mean of all responses in all groups + 2 × SD). For variant comparisons, the mean response over all peptide variants starting at HXB2 Env position aa304 was calculated per vaccination group. Calculations were carried out using R version 3.5.1 and Microsoft Excel.
4. Discussion
Induction of broadly neutralizing antibodies (bnAbs) that confer sterile immunity will probably be a key feature of an effective HIV-1 vaccination strategy. The structure and function of HIV-1 Env has been the center of intense research efforts, being the sole viral protein on the surface of virions. Over the years, increasing the stability of the HIV-1 Env trimer and thereby generating native-like pre-fusion closed trimer mimetics has been deemed extremely important (as reviewed by [
18]).
In the current study, we presented a set of Env variants (one gp120 monomer and four gp140 trimers) derived from a novel clade C consensus sequence. These four trimers were stabilized in a stepwise manner by selective incorporation of previously identified amino acid substitutions that help stabilize native-like trimer mimetics. With an increasing degree of stabilization, we observed improved biochemical and biophysical protein properties as well as favorable antigenicity profiles towards bnAbs and non-nAbs. When tested as immunogens in mice, these in vitro findings translated into a favorable immunological outcome.
Biochemical analyses revealed that the predicted stepwise increasing degree of stability introduced by our protein design turned out as intended. The mostly unmodified trimer ConCv1 KIKO NFL, where the gp120 subunit was only loosely attached to gp41 by the native flexible linker [
9], tended to aggregate. However, it transformed into the well-behaved trimer ConCv5 KIKO, which is secreted from cells as homogeneous protein species consisting primarily of trimers. Of note, neither of our presented trimers were purified by negative or positive selection. In parallel, manufacturability of the protein variants could be improved, as derived from continuously increasing trimer yields. For ConCv1 KIKO NFL, we repeatedly observed different multimerization states of the protomers under non-reducing denaturing conditions. Interestingly, this finding was not confirmed under native conditions where only trimers could be found. Analytical SEC of final protein pools (after preparative SEC) also did not suggest the presence of multimers of trimers.
Further biophysical analyses with regard to thermal unfolding, structural integrity, and particle size confirmed our initial assessment. The ConCv1 KIKO NFL trimer appeared more open with a cumulant radius of 10 nm. The particle sizes of the three SOSIP-based trimers ConCv2, v4, and v5 were comparable. Melting temperatures, however, continuously increased, albeit not to the same extent as has been reported previously for the clade C 16055 trimer that was stabilized in a comparable manner [
34]. Interestingly, the modifications added to v5 (amongst others a second disulfide bond) increased the melting temperature but at the same time led to a loss in cooperativity of the unfolding compared to v2 and v4. Nevertheless, structural integrity gauged by the proportion of closed trimers in negative stain electron micrographs was highest for ConCv5 KIKO and ConCv4 KIKO without enriching closed trimers by either negative or positive selection-based purification strategies.
Continuously decreasing binding affinities and overall signal intensities of several non-nAbs (F105, 447-52D, 17b, 5F3) supported our intended stabilization approach. Along the same lines, quaternary structure-dependent bnAbs (PGT145, PGT151, PG16) primarily bound to the most stabilized trimers ConCv4 KIKO and ConCv5 KIKO. For all other non-structure-dependent bnAbs, we found comparable binding patterns for all trimers, which was not surprising based on their high similarity. Interestingly, binding affinities of some non-structure dependent bnAbs (10-1074, PGT121, 2G12) towards the gp120 monomer decreased by a factor of three to six compared to the trimers. However, overall, binding of antibodies to the gp120 monomer was in most cases intermediate.
Immunization of Balb/c mice confirmed observations from the in vitro assays. The more open trimers induced autologous Env binding titers more rapidly and with higher plateaus. Based on responses towards selected Env regions (V1V2 and V3), however, quantity did not go hand in hand with quality. We therefore hypothesized that the high titers in mice immunized with more open trimers were primarily driven by antibodies targeting more open structure, including V3-directed antibodies (
Figure S2b). Almost all mice produced antibodies that bound the most stabilized trimer ConCv5 KIKO. Interestingly, the median midpoint titers towards this protein were nearly identical for all groups except group 1 (immunized with the gp120 monomer). Only the scatter of the midpoint titers induced in the single animals within the individual groups increased with decreasing degree of stabilization.
The reduction in overall immunogenicity with increasing degree of stabilization was confirmed by peptide microarray analysis. While the overall pattern of IgG Env recognition was comparable between the gp120 monomer, the ConC trimers, and also the clade A BG505 SOSIP.664, we observed a decrease in breadth and depth of IgG recognition of linear gp120 epitopes with increasing stabilization of the ConC immunogens. This was most apparent in the more N-terminal region of gp120 (IDR5). Recognition of the linear V3 region (IDR4) continuously decreased with increasing degree of conformational stability and was almost completely abrogated in animals immunized with ConCv5 KIKO. This variant contained the T316W substitution designed to retain the V3 loop in its ground state location, and its capability to abrogate induction of V3 directed antibodies has been demonstrated before in mice and in rabbits [
35]. Binding of the V1V2 region, which has been associated with protection in RV144 [
27,
42], was low towards the more open trimers v1, v2, and v4, but rather strong towards the most stable trimer ConCv5 KIKO and, interestingly, also towards the ConC KIKO gp120 monomer. Responses towards BG505 SOSIP.664 were comparable to ConCv5 KIKO. In contrast to the differential V1V2 recognition found with the 15-mer peptides by peptide microarray analysis, binding assessed by ELISA towards the V1V2 recombinant proteins AE.A244 and C.1086 covering the complete V1V2 region was more homogeneous. More biologically relevant data might be obtained using V1V2 gp70 scaffolds that mimic a more native conformation of this region. While offering the opportunity to scan Env-specific antibody responses against a large variety of peptides, the peptide microarray analysis is limited in that it reports on binding of linear 15-mer epitopes. Taking this into consideration, however, we nevertheless confirmed our previous finding of an inverse correlation between the degree of conformational stability and overall immunogenicity of our ConC-derived immunogens in mice.
Despite the limited antibody repertoire of mice, this observation is also in line with earlier peptide microarray analyses of clinical trials with Env-based immunogens conducted by us [
38,
39,
40,
43] and others [
42,
48]. Immunogens based on gp120 monomers such as those used in RV144 [
42] and UKHVC003 [
38,
43] in general induced stronger and broader Env-specific IgG responses, targeting various antigenic regions on Env, while the membrane-tethered, more native-like trimeric proteins encoded by MVA-CMDR used in the HIVIS and TaMoVac trials [
38,
39,
40] induced more focused and narrow antibody responses.
Despite favorable biochemical, biophysical, and—to the extent analyzed—immunological properties of ConCv4 KIKO or ConCv5 KIKO, we refrained from probing neutralizing activity of the obtained mouse sera for several reasons: (i) Volumes of sera were small and did not allow a comprehensive analysis of neutralization potency and breadth. (ii) Pseudotype assays performed with mouse sera following Env immunization are known to suffer from a high background that cannot easily be removed by e.g., negative selection or affinity purification. (iii) Mice have a limited antibody repertoire that does not match the diversity of the human response. It is therefore well accepted that following selection of promising candidates in mouse models, breadth and potency of neutralizing antibodies is generally determined in guinea pigs or rabbits. Immunizations with a single stabilized HIV Env, including some clade C stabilized trimers, have so far proven the potential to induce neutralizing antibodies, which are in most cases limited to the autologous pseudovirus and tier 1 pseudoviruses. Broadly neutralizing nanobodies have successfully been induced in camels. However, some limited breadth of neutralizing antibodies towards tier 2 isolates has been observed only in the context of prime-boost regimens. Exemplarily, using the fusion peptide coupled to a carrier providing T cell help followed by booster immunizations using a carefully selected panel of stabilized Env trimers [
49] or using a panel of glycan-modified HIV NFL envelope trimers representing different HIV clades displayed on liposomes [
50], proved successful in the induction of tier 2 neutralizing antibodies. Great hopes are also associated with a strategy where a carefully orchestrated series of consecutive guiding immunogens are administered to drive maturation of initially targeted naïve B cell lineages towards bnAb responses [
51].
The experiments and results presented in this study were generated within in the European HIV Vaccine Alliance (EHVA), one of the two major HIV vaccine networks funded under the EU H2020 program. The ConCv5 KIKO Env variant studied here is under further investigation in rabbit and NHP trials. These and other animal trials explore the potential of this stabilized trimer and rationally designed derivatives thereof to contribute to the induction of neutralization breadth, pursuing some of the strategies described above [
52]. The results of the here presented study ultimately contributed—together with additional data—to the selection and promotion of the described ConCv5 KIKO pre-fusion stabilized clade C trimer for GMP manufacturing and testing in a phase 1 clinical trial.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}