
Effect of Bivalent Vaccines against Vibrio anguillarum and Aeromonas salmonicida Subspecie achromogenes on Health and Survival of Turbot

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Vaccines
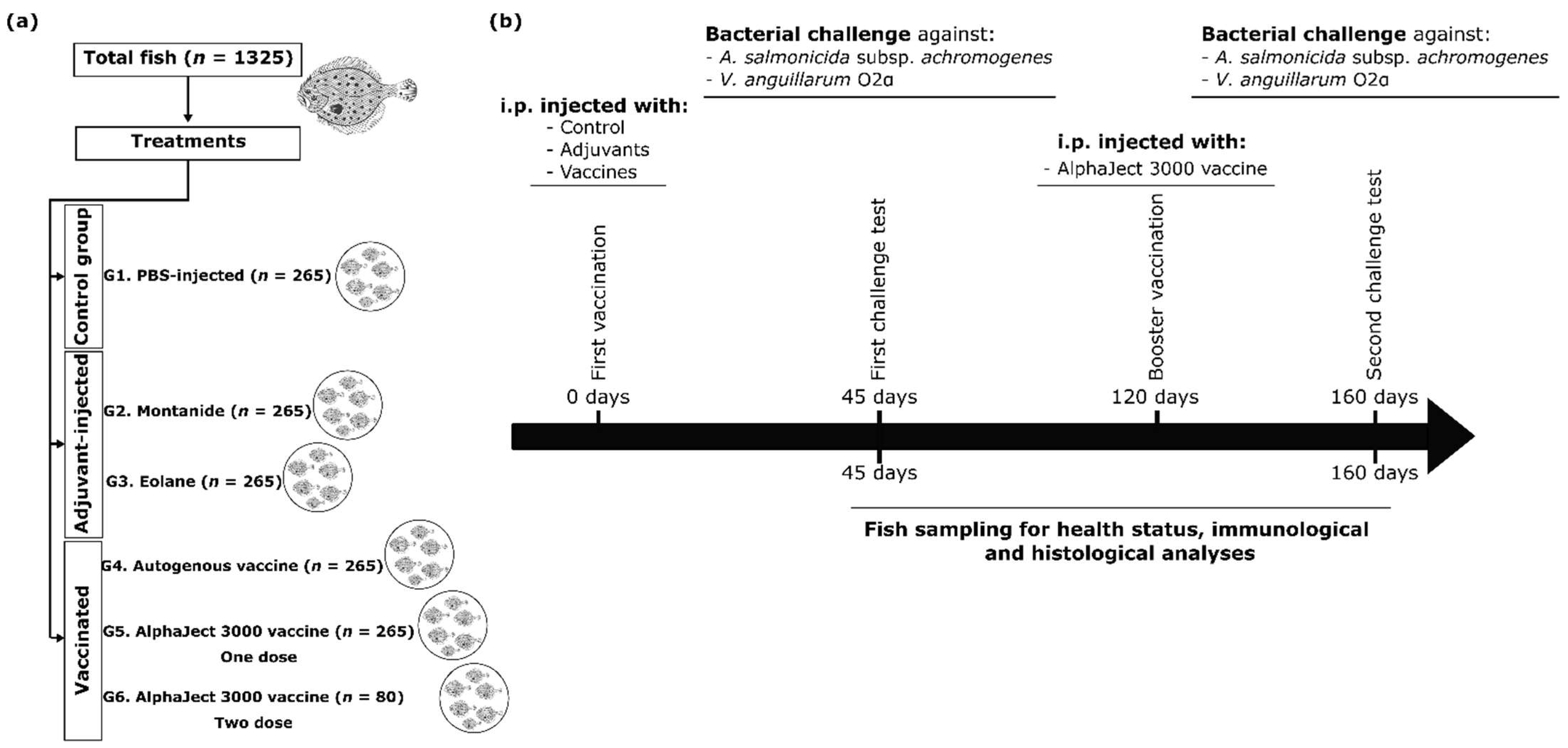
2.3. Experimental Fish, Rearing Conditions, and Vaccination Regimes
2.4. Sampling Procedures and Analysis of Fish Health Status
2.5. Efficacy of Vaccination
3. Results
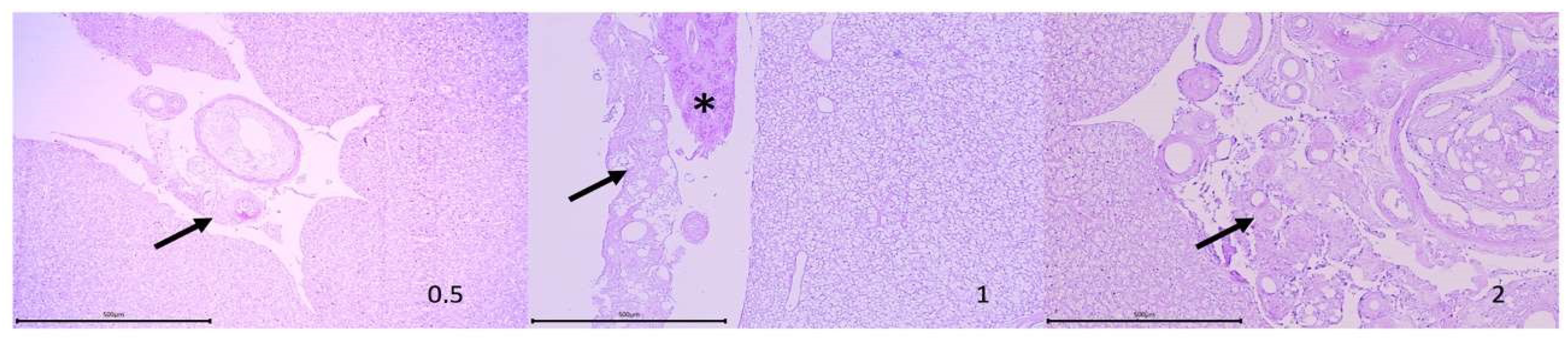
3.1. Macroscopic and Histological Effect of Vaccination
3.2. Evaluation of Physiological Parameters
3.3. Efficacy of Vaccines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. El Estado Mundial de la Pesca y la Acuicultura 2018. Cumplir los Objetivos de Desarrollo Sostenible; FAO: Rome, Italy, 2018; ISBN 978-92-5-130688-8. [Google Scholar]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens, 6th ed.; Springer: Chichester, UK, 2016; ISBN 978-3-319-32673-3. [Google Scholar]
- Novotny, A.J. Vibriosis and furunculosis in marine cultured salmon in Puget Sound, Washington. Mar. Fish. Rev. 1978, 40, 52–55. [Google Scholar]
- García Carballas, C.M. Respuesta Inmune en Rodaballo (Psetta máxima) y Lenguado (Solea sp). Efecto de la Vacunación. Ph.D. Thesis, Universidade de Santiago de Compostela, Santiago, Spain, 2018. [Google Scholar]
- Wiklund, T.; Dalsgaard, I. Occurrence, and significance of atypical Aeromonas salmonicida in non-salmonid and salmonid fish species: A review. Dis. Aquat. Organ. 1998, 32, 49–69. [Google Scholar] [CrossRef] [Green Version]
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Santos, Y.; Pazos, F.; Bandin, I.; Toranzo, A.E. Analysis of antigens present in the extracellular products and cell surface of Vibrio anguillarum serotypes O1, O2, and O3. Appl. Environ. Microbiol. 1995, 61, 2493–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondad-Reantaso, M.G.; Arthur, J.R.; Subasinghe, R.P. Improving biosecurity through prudent and responsible use of veterinary medicines in aquatic food production. In FAO Fisheries and Aquaculture Technical Paper; No. 547; FAO: Rome, Italy, 2012; ISBN 2070-7010. [Google Scholar]
- Håstein, T.; Gudding, R.; Evensen, O. Bacterial vaccines for fish—An update of the current situation worldwide. Dev. Biol. 2005, 121, 55–74. [Google Scholar]
- Björnsdóttir, B.; Gudmundsdóttir, S.; Bambir, S.H.; Gudmundsdóttir, B.K. Experimental infection of turbot, Scophthalmus maximus (L.), by Aeromonas salmonicida subsp. achromogenes and evaluation of cross protection induced by a furunculosis vaccine. J. Fish Dis. 2005, 28, 181–188. [Google Scholar] [CrossRef]
- Santos, Y.; García-Marquez, S.; Pereira, P.G.; Pazos, F.; Riaza, A.; Silva, R.; El Morabit, A.; Ubeira, F.M. Efficacy of furunculosis vaccines in turbot, Scophthalmus maximus (L.): Evaluation of immersion, oral and injection delivery. J. Fish Dis. 2005, 28, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Toranzo, A.E.; Santos, Y.; Barja, J.L. Immunization with bacterial antigens: Vibrio infections. Dev. Biol. Stand. 1997, 90, 90–105. [Google Scholar]
- European pharmacopoeia. Vibriosis (cold-water) vaccine (inactivated) for salmonids. In European pharmacopoeia; Council of Europe, European Directorate for the Quality of Medicines and Healthcare: Strasbourg, France, 2010; pp. 2–3. [Google Scholar]
- Mikkelsen, H.; Lund, V.; Larsen, R.; Seppola, M. Vibriosis vaccines based on various sero-subgroups of Vibrio anguillarum O2 induce specific protection in Atlantic cod (Gadus morhua L.) juveniles. Fish Shellfish Immunol. 2011, 30, 330–339. [Google Scholar] [CrossRef]
- Liu, X.; Jiao, C.; Ma, Y.; Wang, Q.; Zhang, Y. A live attenuated Vibrio anguillarum vaccine induces efficient immunoprotection in Tiger puffer (Takifugu rubripes). Vaccine 2018, 36, 1460–1466. [Google Scholar] [CrossRef]
- Vigneulle, M.; Laurencin, F.B. Uptake of Vibrio anguillarum bacterin in the posterior intestine of rainbow trout Oncorhynchus mykiss, sea bass Dicentrarchus labrax and turbot or anal intubation. Dis. Aquat. Organ. 1991, 11, 85–92. [Google Scholar] [CrossRef]
- Norqvist, A.; Hagström, A.; Wolf-Watz, H. Protection of rainbow trout against vibriosis and furunculosis by the use of attenuated strains of Vibrio anguillarum. Appl. Environ. Microbiol. 1989, 55, 1400–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villumsen, K.R.; Koppang, E.O.; Christensen, D.; Bojesen, A.M. Alternatives to mineral oil adjuvants in vaccines against Aeromonas salmonicida subsp. salmonicida in rainbow trout offer reductions in adverse effects. Sci. Rep. 2017, 7, 5930. [Google Scholar] [CrossRef] [Green Version]
- Bao, P.; Sun, X.; Liu, Q.; Zhang, Y.; Liu, X. Synergistic effect of a combined live Vibrio anguillarum and Edwardsiella piscicida vaccine in turbot. Fish Shellfish Immunol. 2019, 88, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Ellis, A.E. Immunization with bacterial antigens: Furunculosis. Dev. Biol. Stand. 1997, 90, 107–116. [Google Scholar] [PubMed]
- Buller, N. Bacteria and Fungi from Fish and Other Aquatic Animals, 2nd ed.; CABI Publishing: Wallingford, UK, 2014; ISBN 0851997384. [Google Scholar]
- Midtlyng, P.J.; Lillehaug, A. Growth of Atlantic salmon Salmo salar after intraperitoneal administration of vaccines containing adjuvants. Dis. Aquat. Organ. 1998, 32, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Morabit, A.; Carballas, C.G.; Seoane, R.; Riaza, A.; Santos, Y. Flow cytometric study of phagocytic and oxidative burst activities of turbot head-kidney leucocytes in response to Aeromonas salmonicida subspecies salmonicida and Tenacibacterium maritimum. In Recent Research Developments in Microbiology; Panadalai, S.G., Ed.; Research Signpost: New York, NY, USA, 2006; Volume 10, ISBN 81-308-0022-5. [Google Scholar]
- Melingen, G.O.; Wergeland, H.I. Physiological effects of an oil-adjuvanted vaccine on out-of-season Atlantic salmon (Salmo salar L.) smolt. Aquaculture 2002, 214, 397–409. [Google Scholar] [CrossRef]
- Durbin, A.P.; Cho, C.J.; Elkins, W.R.; Wyatt, L.S.; Moss, B.; Murphy, B.R. Comparison of the immunogenicity and efficacy of a replication-defective vaccinia virus expressing antigens of human parainfiuenza virus type 3 (HPIV3) with those of a live attenuated HPIV3 vaccine candidate in rhesus monkeys passively immunized with PIV3 antibodies. J. Infect. Dis. 1999, 179, 1345–1351. [Google Scholar] [CrossRef] [Green Version]
- Menanteau-Ledouble, S.; Kumar, G.; Saleh, M.; El-Matbouli, M. Aeromonas salmonicida: Updates on an old acquaintance. Dis. Aquat. Organ. 2016, 120, 49–68. [Google Scholar] [CrossRef] [Green Version]
- Midtlyng, P.J. Vaccinated fish welfare: Protection versus side-effects. Dev. Biol. Stand. 1997, 90, 371–379. [Google Scholar]
- Koppang, E.O.; Haugarvoll, E.; Hordvik, I.; Aune, L.; Poppe, T.T. Vaccine-associated granulomatous inflammation and melanin accumulation in Atlantic salmon, Salmo salar L., white muscle. J. Fish Dis. 2005, 28, 13–22. [Google Scholar] [CrossRef]
- Berg, A.; Rødseth, O.M.; Hansen, T. Fish size at vaccination influence the development of side-effects in Atlantic salmon (Salmo salar L.). Aquaculture 2007, 265, 9–15. [Google Scholar] [CrossRef]
- Coscelli, G.A.; Bermúdez, R.; Losada, A.P.; Santos, Y.; Quiroga, M.I. Vaccination against Aeromonas salmonicida in turbot (Scophthalmus maximus L.): Study of the efficacy, morphological changes, and antigen distribution. Aquaculture 2015, 445, 22–32. [Google Scholar] [CrossRef]
- Noia, M.; Domínguez, B.; Leiro, J.; Blanco-Méndez, J.; Luzardo-Álvarez, A.; Lamas, J. Inflammatory responses and side effects generated by several adjuvant-containing vaccines in turbot. Fish Shellfish Immunol. 2014, 38, 244–254. [Google Scholar] [CrossRef]
- Aucouturier, J.; Dupuis, L.; Ganne, V. Adjuvants designed for veterinary and human vaccines. Vaccine 2001, 19, 2666–2672. [Google Scholar] [CrossRef]
- Jaafar, R.M.; Chettri, J.K.; Dalsgaard, I.; Al-Jubury, A.; Kania, P.W.; Skov, J.; Buchmann, K. Effects of adjuvant MontanideTM ISA 763 A VG in rainbow trout injection vaccinated against Yersinia ruckeri. Fish Shellfish Immunol. 2015, 47, 797–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romstad, A.B.; Reitan, L.J.; Midtlyng, P.; Gravningen, K.; Evensen, Ø. Antibody responses correlate with antigen dose and in vivo protection for oil-adjuvanted, experimental furunculosis (Aeromonas salmonicida subsp. salmonicida) vaccines in Atlantic salmon (Salmo salar L.) and can be used for batch potency testing of vaccines. Vaccine 2013, 31, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Mutoloki, S.; Alexandersen, S.; Gravningen, K.; Evensen, Ø. Time-course study of injection site inflammatory reactions following intraperitoneal injection of Atlantic cod (Gadus morhua L.) with oil-adjuvanted vaccines. Fish Shellfish Immunol. 2008, 24, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, D.; González, M.T. Evaluation of somatic indexes, hematology, and liver histopathology of the fish Labrisomus philippii from San Jorge Bay, northern Chile, as associated with environmental stress. Rev. Biol. Mar. Oceanogr. 2012, 47, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Rønsholdt, B.; McLean, E. The effect of vaccination and vaccine components upon short-term growth and feed conversion efficiency in rainbow trout. Aquaculture 1999, 174, 213–221. [Google Scholar] [CrossRef]
- Berg, A.; Rodseth, O.M.; Tangerás, A.; Hansen, T. Time of vaccination influences development of adhesion, growth, and spinal deformities in Atlantic salmon Salmo salar. Dis. Aquat. Org. 2006, 69, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Tang, L.; Li, S.; Li, G.; Mo, Z. The efficacy and side-effects of oil-based adjuvants emulsified Vibrio anguillarum bivalent inactivated vaccine in turbot (Scophthalmus maximus) under production mode. Aquaculture 2020, 524, 735259. [Google Scholar] [CrossRef]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Satheeshkumar, P.; Ananthan, G.; Kumar, D.S.; Jagadeesan, L. Haematology and biochemical parameters of different feeding behaviour of teleost fishes from Vellar estuary, India. Comp. Clin. Path. 2012, 21, 1187–1191. [Google Scholar] [CrossRef]
- Ivanc, A.; Haskovic, E.; Jeremic, S.; Dekic, R. Hematological evaluation of welfare and health of fish. Prax. Vet. 2016, 53, 191–202. [Google Scholar]
- Cazenave, J.; Wunderlin, D.A.; Hued, A.C.; Bistoni, M.D.L.Á. Haematological parameters in a neotropical fish, Corydoras paleatus (Jenyns, 1842) (Pisces, Callichthyidae), captured from pristine and polluted water. Hydrobiologia 2005, 537, 25–33. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeong, M.H.; Jun, J.C.; Kim, T.I. Changes in hematological, biochemical, and non-specific immune parameters of olive flounder, Paralichthys olivaceus, following starvation. Asian Australas. J. Anim. Sci. 2014, 27, 1360–1367. [Google Scholar] [CrossRef]
- Faílde, L.D.; Losada, A.P.; Bermúdez, R.; Santos, Y.; Quiroga, M.I. Tenacibaculum maritimum infection: Pathology and immunohistochemistry in experimentally challenged turbot (Psetta maxima L.). Microb. Pathog. 2013, 65, 82–88. [Google Scholar] [CrossRef]
- Gudmundsdóttir, B.K.; Björnsdóttir, B. Vaccination against atypical furunculosis and winter ulcer disease of fish. Vaccine 2007, 25, 5512–5523. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Macroscopic Effects Spielberg Scale (Ss) | Microscopic Effects Semiquantitative Scale a | ||||||
|---|---|---|---|---|---|---|---|
| Fish Group | Days Post-Treatment | Liver | |||||
| 45 | 160 | Vacuolization | Adhesion on Serosa | Bile duct Fibriosis | Degeneration/Necrosis | Renal tubules Vacuolation | |
| G1 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.00 ± 0.38 | 0.00 ± 0.00 | 1.00 ± 0.00 | 1.13 ± 0.23 | 0.43 ± 0.19 |
| G2 | 2.60 ± 0.99 | 0.40 ± 0.97 | 1.94 ± 0.73 | 0.13 ± 0.23 | 1.13 ± 0.59 | 0.44 ± 0.42 | 0.69 ± 0.37 |
| G3 | 0.68 ± 0.95 | 0.40 ± 0.97 | 1.75 ± 0.53 | 0.19 ± 0.37 | 1.00 ± 0.00 | 0.19 ± 0.37 | 0.69 ± 0.59 |
| G4 | 5.65 ± 0.75 | 5.50 ± 0.53 | 1.81 ± 0.59 | 0.88 ± 0.79 | 1.00 ± 0.00 | 0.81 ± 0.59 | 0.88 ± 0.52 |
| G5 | 4.45 ± 1.05 | 2.50 ± 1.07 | 2.00 ± 0.65 | 0.94 ± 0.56 | 1.13 ± 0.59 | 0.69 ± 0.59 | 1.25 ± 0.93 |
| G6 | ND | 3.88 ± 0.35 | 1.81 ± 0.70 | 0.56 ± 0.56 | 1.19 ± 0.37 | 0.94 ± 0.68 | 0.75 ± 0.46 |
| Fish Group | 45 D after Treatment | 160 D after Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fish Weight (g) | VSI (%) | HSI (%) | Hematocrit | SI | Fish Weight (g) | VSI (%) | HSI (%) | Hematocrit | SI | |
| G1 | 94.4 ± 17.0 | 6.0 ± 1.1 b | 1.8 ± 0.3 b | 36.6 ± 13.0 | 2.7 ± 1.5 | 304.8 ± 55.0 b | 5.3 ± 0.2 b | 1.8 ± 0.4 | 26.8 ± 2.3 | 1.7 ± 0.1 |
| G2 | 94.0 ± 16.0 | 6.4 ± 08 b | 2.0 ± 0.3 b | 32.2 ± 11.2 | 1.0 ± 0.4 | 285.0 ± 81.0 | 5.7 ± 0.5 b | 1.6 ± 0.3 | 28.8 ± 4.9 | 2.0 ± 0.2 |
| G3 | 105.7 ± 17.0 | 6.4 ± 0.9 b | 2.0 ± 0.4 b | 42.3 ± 16.2 | ND | 339.5 ± 73.0 b | 5.6 ± 0.5 b | 1.9 ± 0.4 | 27.2 ± 3.1 | 1.8 ± 0.3 |
| G4 | 70.6 ± 14.0 | 10.7 ± 1.8 a | 0.7 ± 1.1 a | 36.4 ± 12.6 | 2.7 ± 0.6 | 205.2 ± 58.0 a | 6.8 ± 0.9 a | 1.7 ± 0.2 | 32.2 ± 6.2 | 2.4 ± 0.6 |
| G5 | 88.3 ± 16.0 | 7.6 ± 0.9 a | 2.2 ± 0.3 b | 41.0 ± 6.3 | 2.1 ± 0.5 | 300.6 ± 42.0 | 5.6 ± 0.3 b | 1.8 ± 0.2 | 32.6 ± 10.5 | 2.8 ± 0.3 |
| G6 | ND | ND | ND | ND | ND | 226.7 ± 46.0 | 5.7 ± 0.5 b | 1.6 ± 0.9 | 32.3 ± 5.8 | 2.2 ± 0.1 |
| Antigens | Fish Groups | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | G2 | G3 | G4 | G5 | G6 | ||||||
| 45d | 160d | 45d | 160d | 45d | 160d | 45d | 160d | 45d | 160d | 160d | |
| A. salmonicida subsp. achromogenes | <2 b | <2 b | <2 b | <2 b | <2 b | <2 b | 1468 ± 878 a | 200 ± 0 b | 174 ± 68 b | 12 ± 10 b | 200 ± 0 b |
| V. anguillarum O2α | 2 ± 0 b | <2 b | <2 b | <2 b | <2 b | <2 b | 637 ± 934 b | 140 ± 104 b | 406 ± 708 b | 16 ± 9 b | 650 ± 900 b |
| Fish Group | Vaccination Procedure | Infectious Dose Cells/Fish | %Mortality | RPS60 | |
|---|---|---|---|---|---|
| Primary Dose | Booster Dose | ||||
| Protection against V. anguillarum 45 d Postvaccination | |||||
| G1 | i.p. injection | 1 × 109 | 60 a | ||
| G2 | i.p. injection | 37 b | 38 | ||
| G3 | i.p. injection | 35 b | 42 | ||
| G4 | i.p. injection | 0 b | 100 | ||
| G5 | i.p. injection | 0 b | 100 | ||
| Protection against A. salmonicida subsp. achromogenes 45 d postvaccination | |||||
| G1 | i.p. injection | 9.9 × 108 | 60 a | ||
| G2 | i.p. injection | 4 b | 93 | ||
| G3 | i.p. injection | 4 b | 93 | ||
| G4 | i.p. injection | 10 b | 83 | ||
| G5 | i.p. injection | 30 b | 50 | ||
| Protection against V. anguillarum 160 d postvaccination | |||||
| G1 | i.p. injection | 6.6 × 1010 | 60 a | ||
| G2 | i.p. injection | 57 | 5 | ||
| G3 | i.p. injection | 56 | 7 | ||
| G4 | i.p. injection | 29 b | 52 | ||
| G5 | i.p. injection | 20 b | 67 | ||
| G6 | i.p. injection | i.p. injection | 24 b | 60 | |
| Protection against A. salmonicida subsp. achromogenes 160 d postvaccination | |||||
| G1 | i.p. injection | 1.4 × 1010 | 60 a | ||
| G2 | i.p. injection | 78 | 0 | ||
| G3 | i.p. injection | 27 b | 55 | ||
| G4 | i.p. injection | 0 b | 100 | ||
| G5 | i.p. injection | 14 b | 77 | ||
| G6 | i.p. injection | i.p. injection | 0 b | 100 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Corral, Y.; Girons, A.; González-Barreiro, O.; Seoane, R.; Riaza, A.; Santos, Y. Effect of Bivalent Vaccines against Vibrio anguillarum and Aeromonas salmonicida Subspecie achromogenes on Health and Survival of Turbot. Vaccines 2021, 9, 906. https://doi.org/10.3390/vaccines9080906
Torres-Corral Y, Girons A, González-Barreiro O, Seoane R, Riaza A, Santos Y. Effect of Bivalent Vaccines against Vibrio anguillarum and Aeromonas salmonicida Subspecie achromogenes on Health and Survival of Turbot. Vaccines. 2021; 9(8):906. https://doi.org/10.3390/vaccines9080906
Chicago/Turabian StyleTorres-Corral, Yolanda, Albert Girons, Oscar González-Barreiro, Rafael Seoane, Ana Riaza, and Ysabel Santos. 2021. "Effect of Bivalent Vaccines against Vibrio anguillarum and Aeromonas salmonicida Subspecie achromogenes on Health and Survival of Turbot" Vaccines 9, no. 8: 906. https://doi.org/10.3390/vaccines9080906