
mRNA Vaccines Enhance Neutralizing Immunity against SARS-CoV-2 Variants in Convalescent and ChAdOx1-Primed Subjects

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vaccination and Control Cohorts
2.2. Serum and PBMC Isolation and Cryopreservation
2.3. Enzyme-Linked Immunosorbent Assays
2.4. Surrogate SARS-CoV-2 Neutralization Test (GenScript)
2.5. Interferon-Gamma (IFN-γ) Release Assay (IGRA)
2.6. SARS-CoV-2 Peptide Stimulation and FACS Analysis
2.7. Statistical Analysis
3. Results
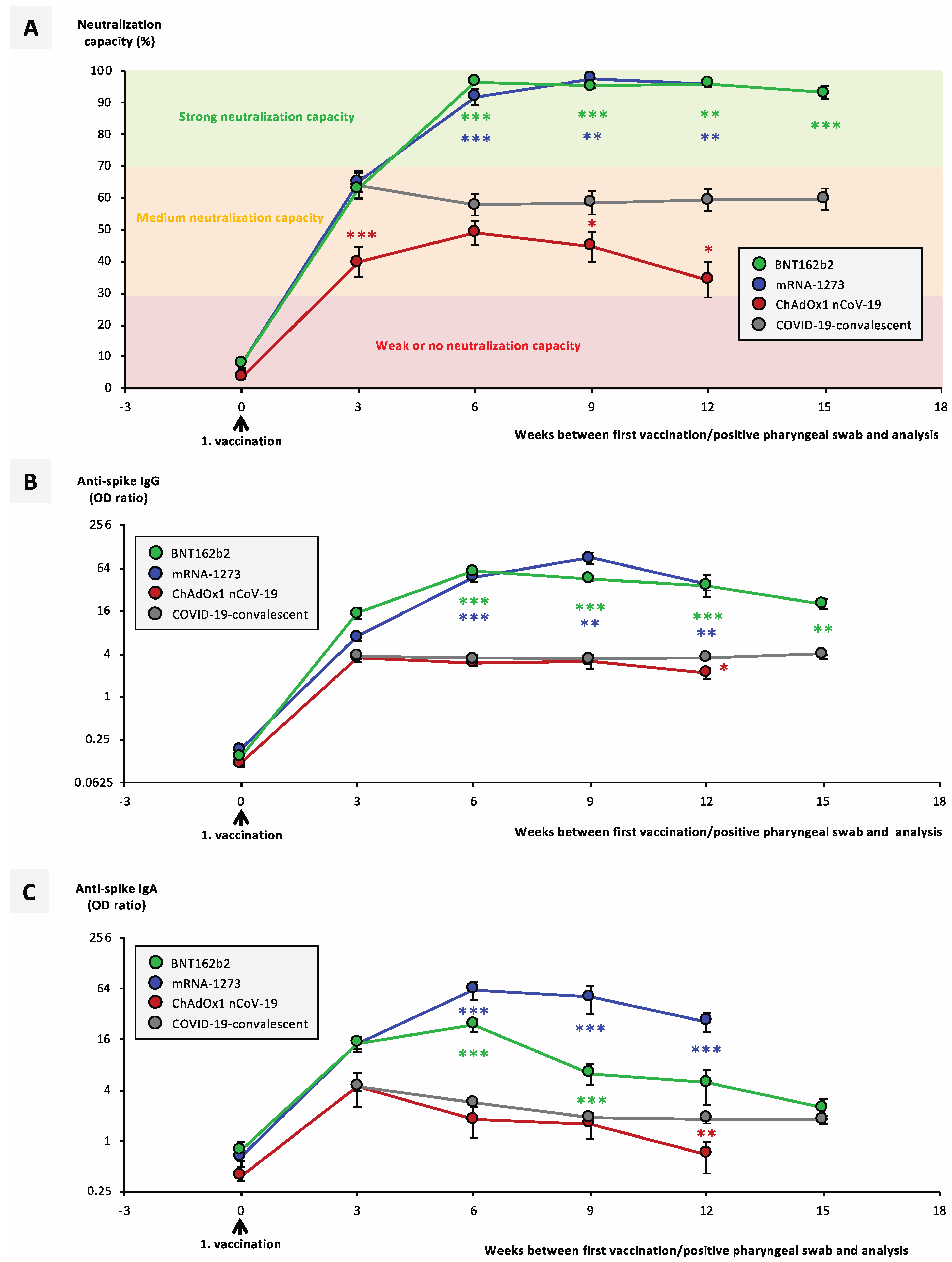
3.1. Temporal Course of Antibody Titers and Neutralization Capacity Significantly Differ between Vaccination Groups and COVID-19-Convalescent Individuals
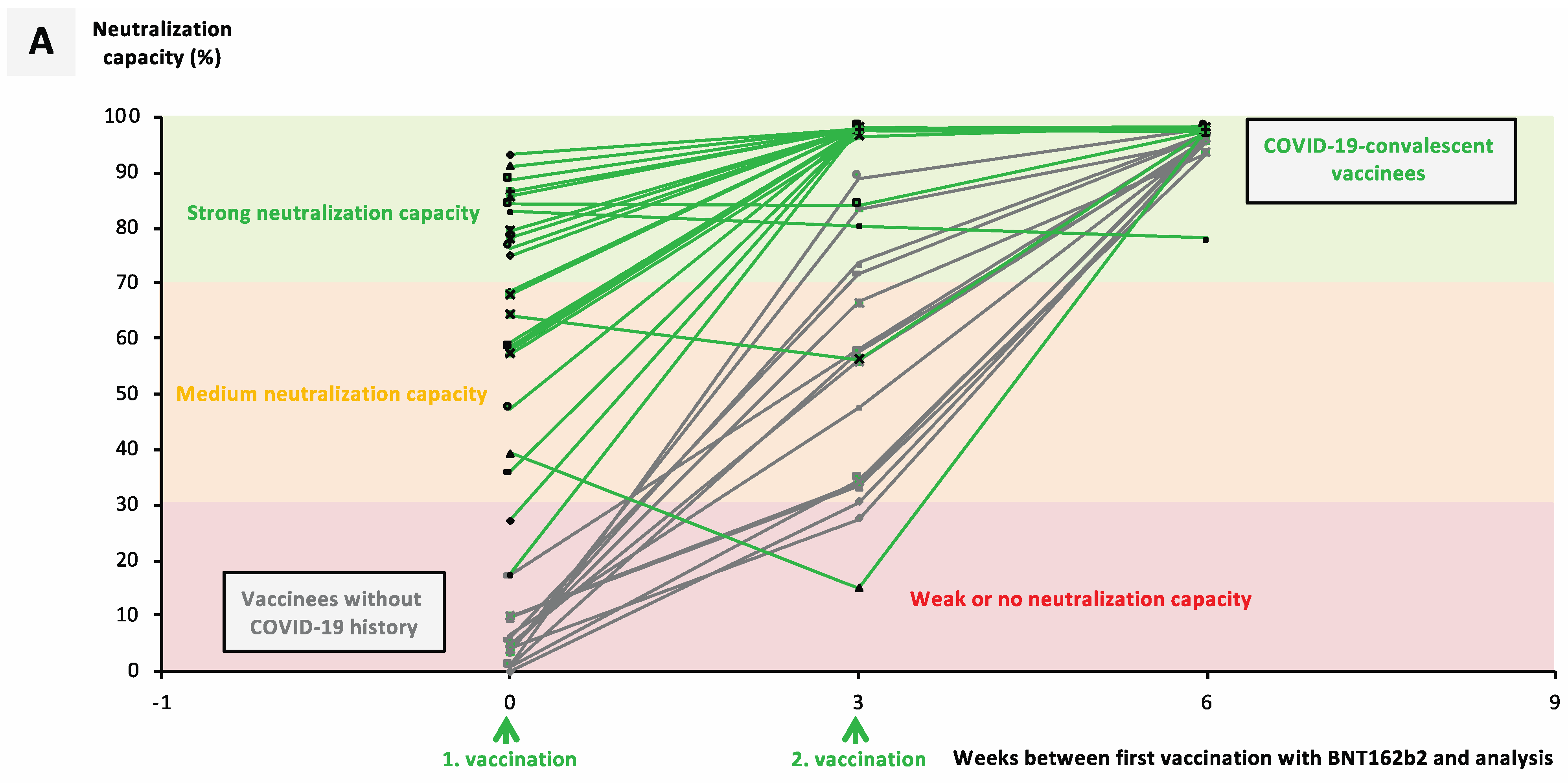
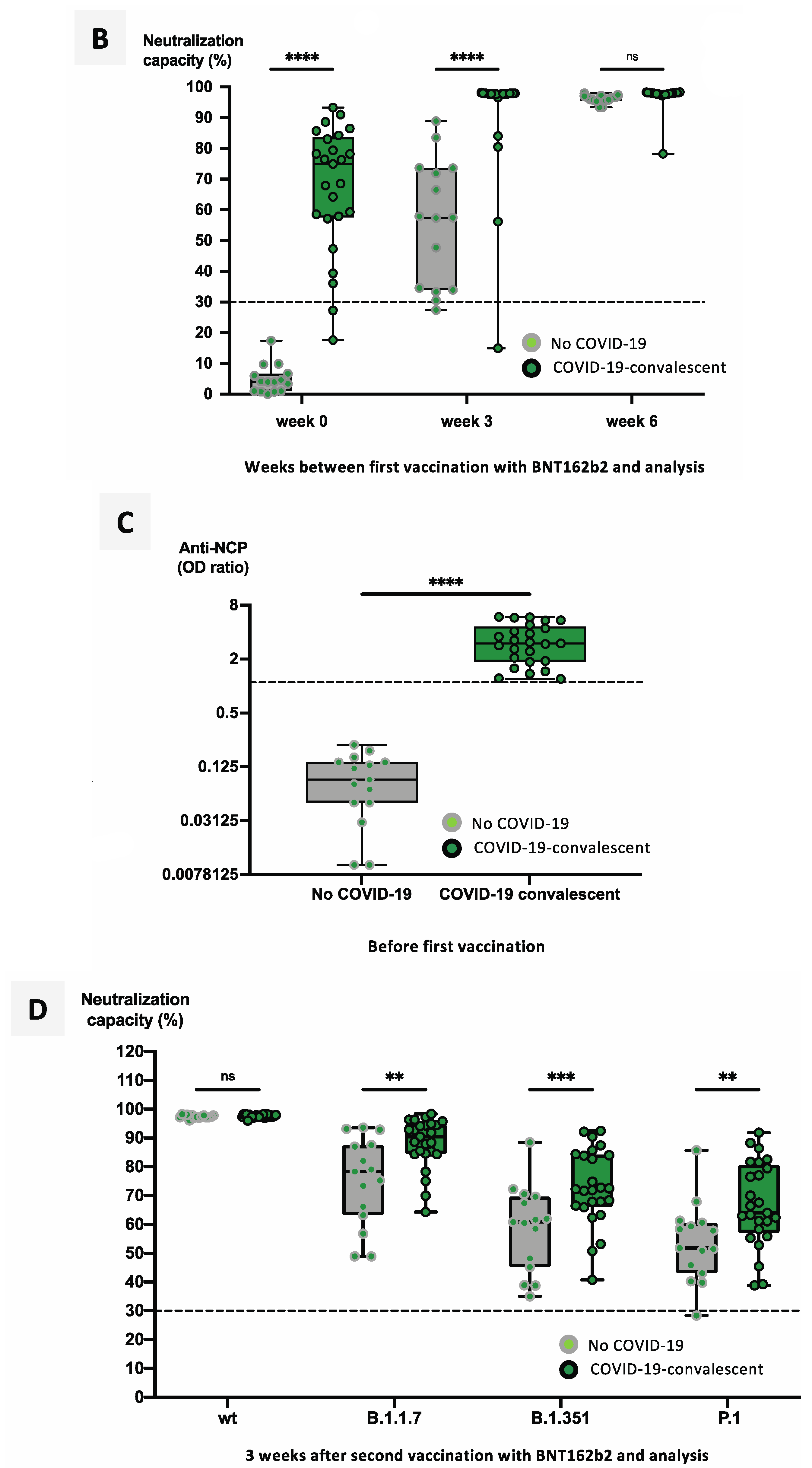
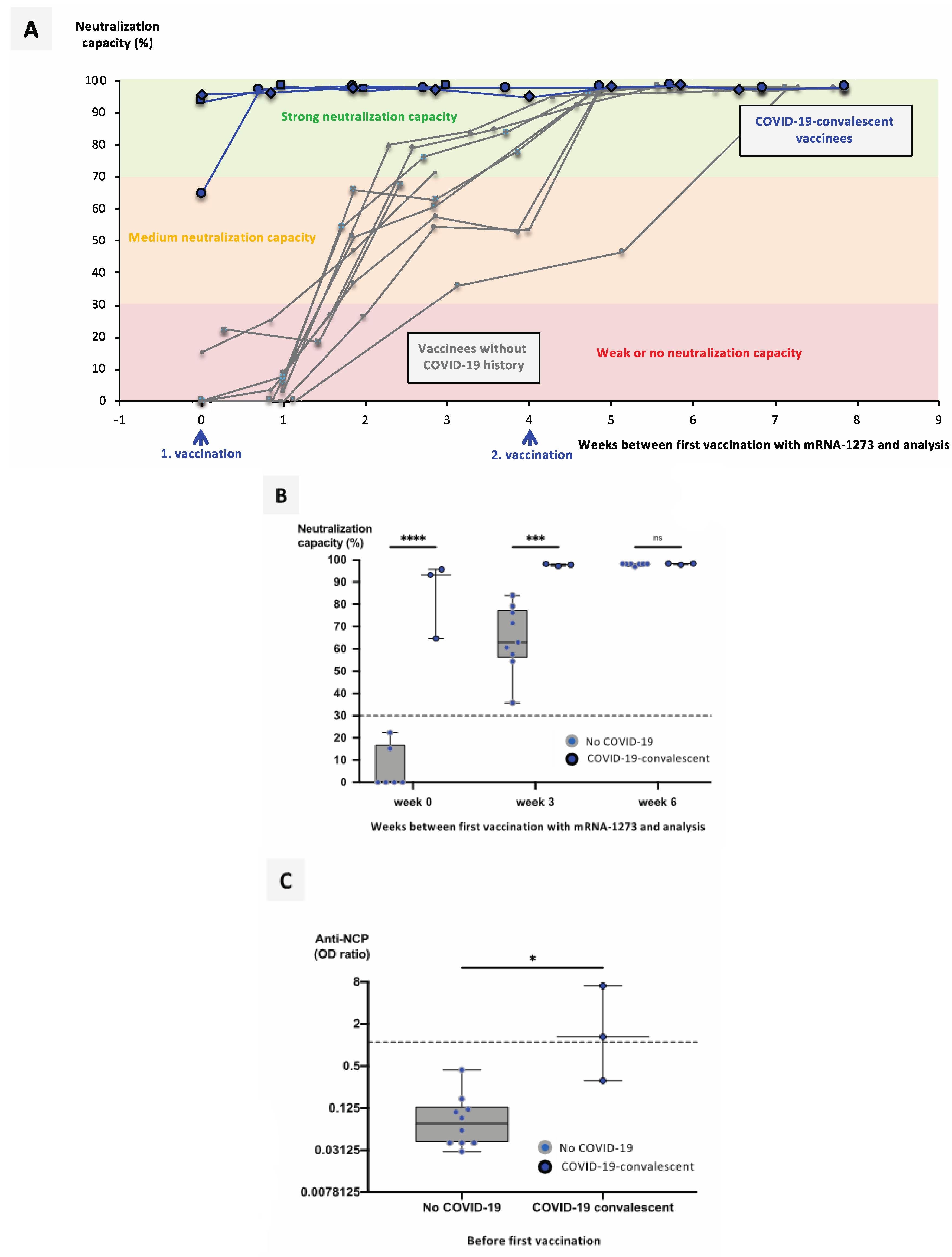
3.2. Previous SARS-CoV-2 Infection Significantly Facilitates Development of a Neutralizing Immune Response after Vaccination with mRNA Vaccines
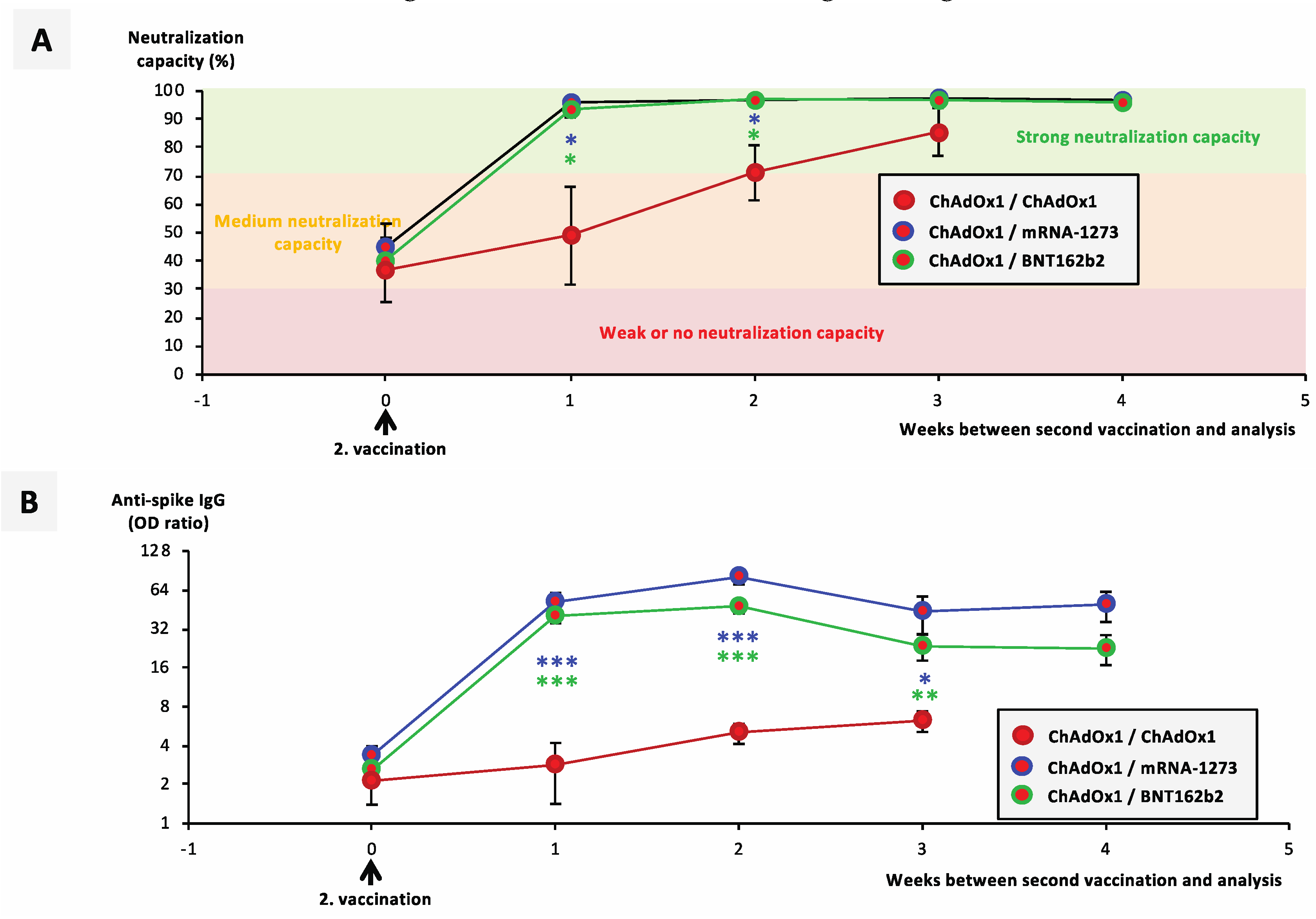
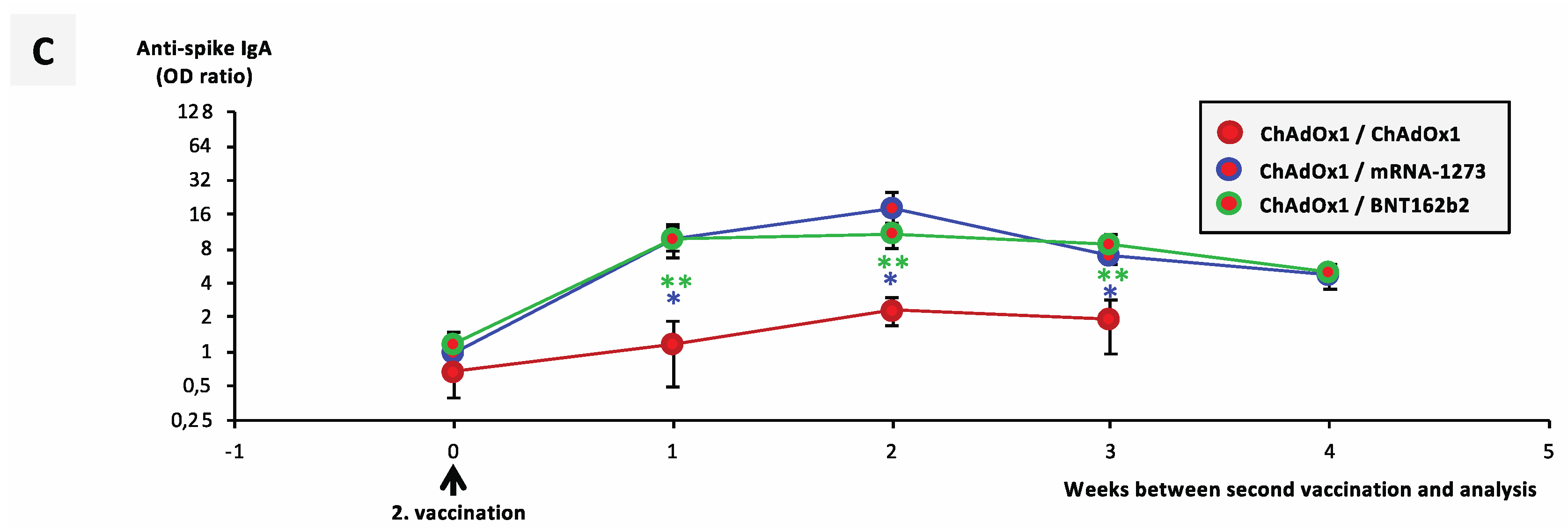
3.3. Booster Vaccination with an mRNA Vaccine Allows Rapid Development of a Neutralizing Immune Response in ChAdOx1-nCoV-19-Vaccinated individuals
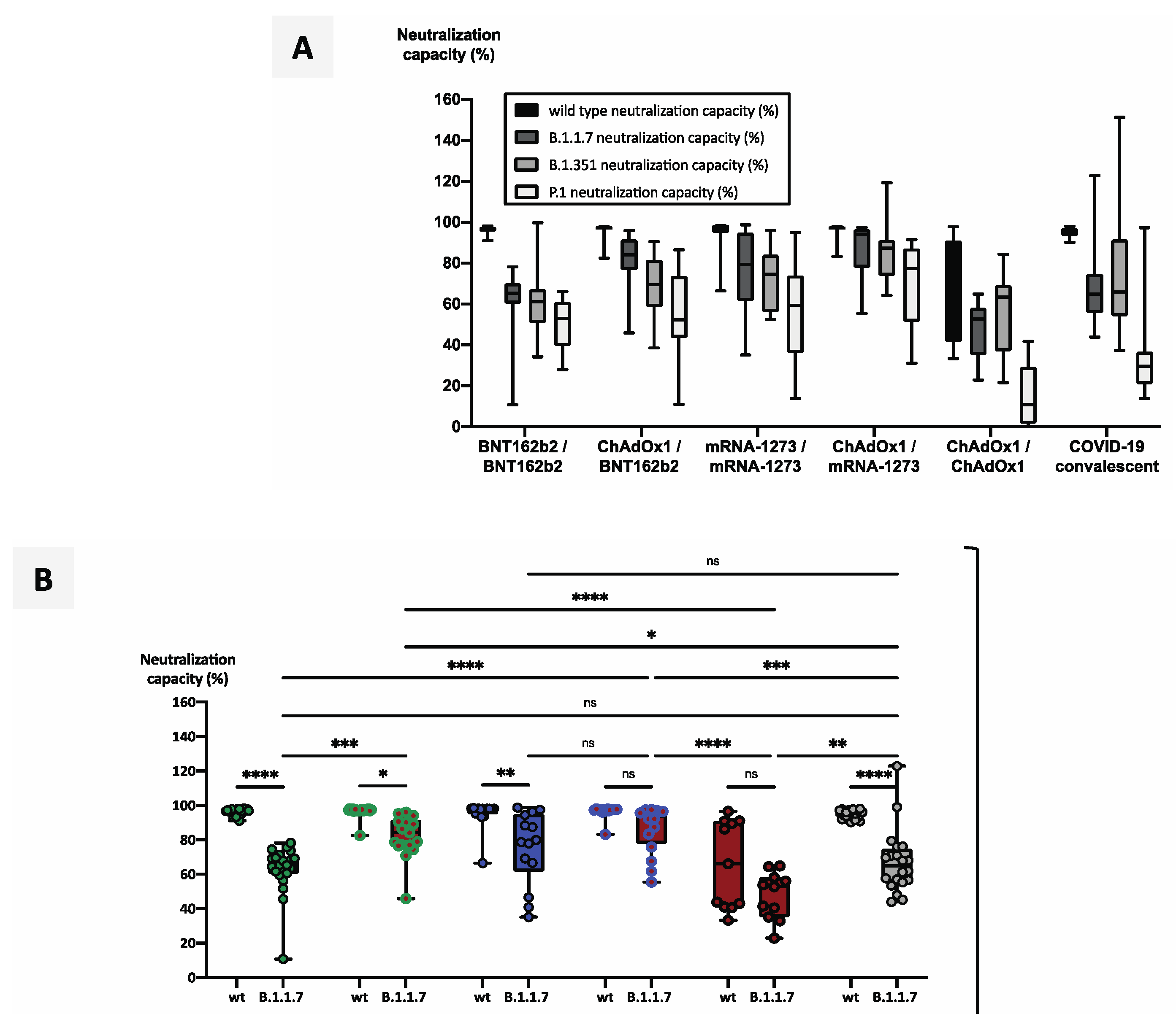
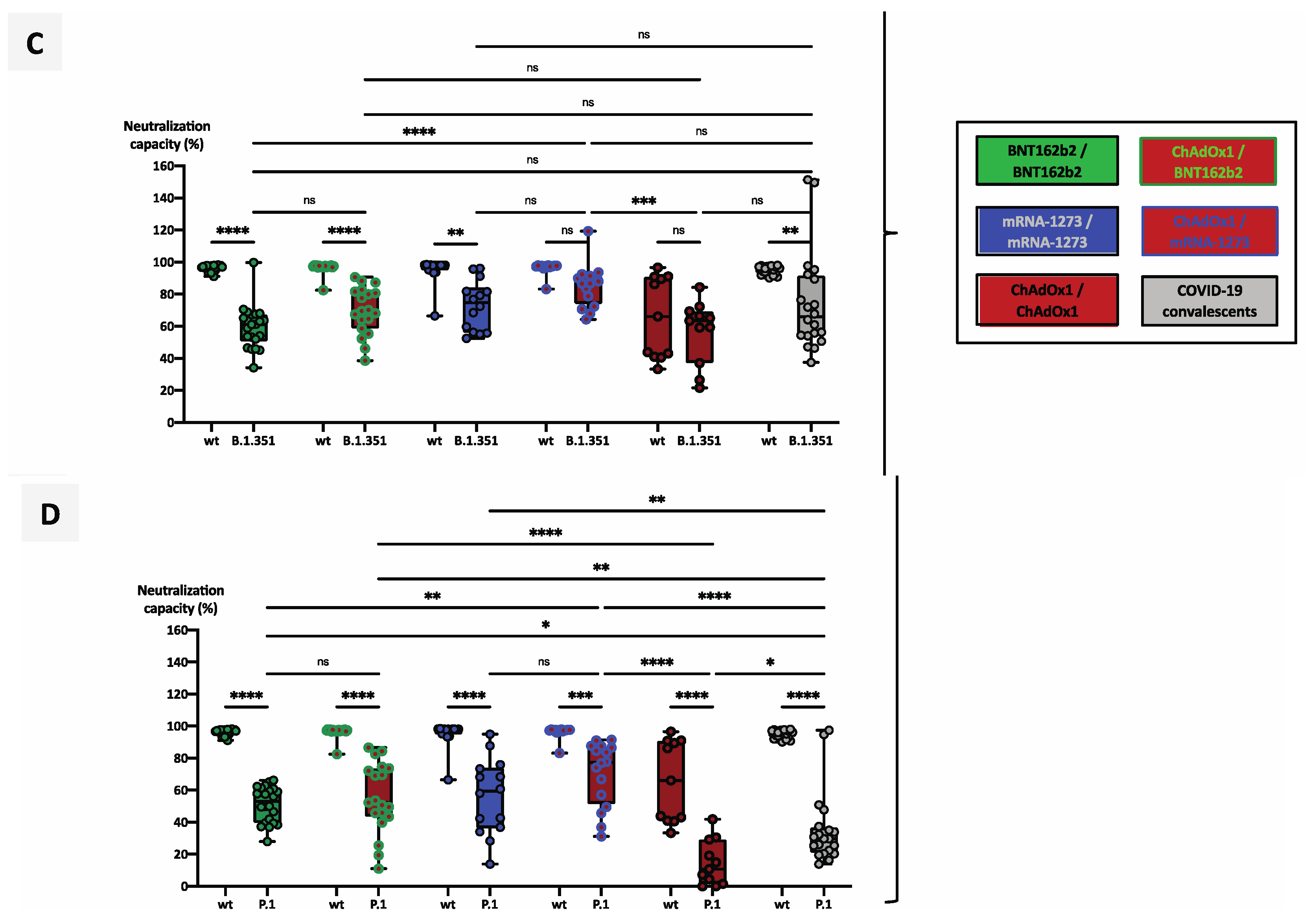
3.4. Heterologous Vaccination Regimes Induce Significantly Stronger Neutralizing Capacity against Variants of Concern than Homologous Schemes or Convalescence Alone
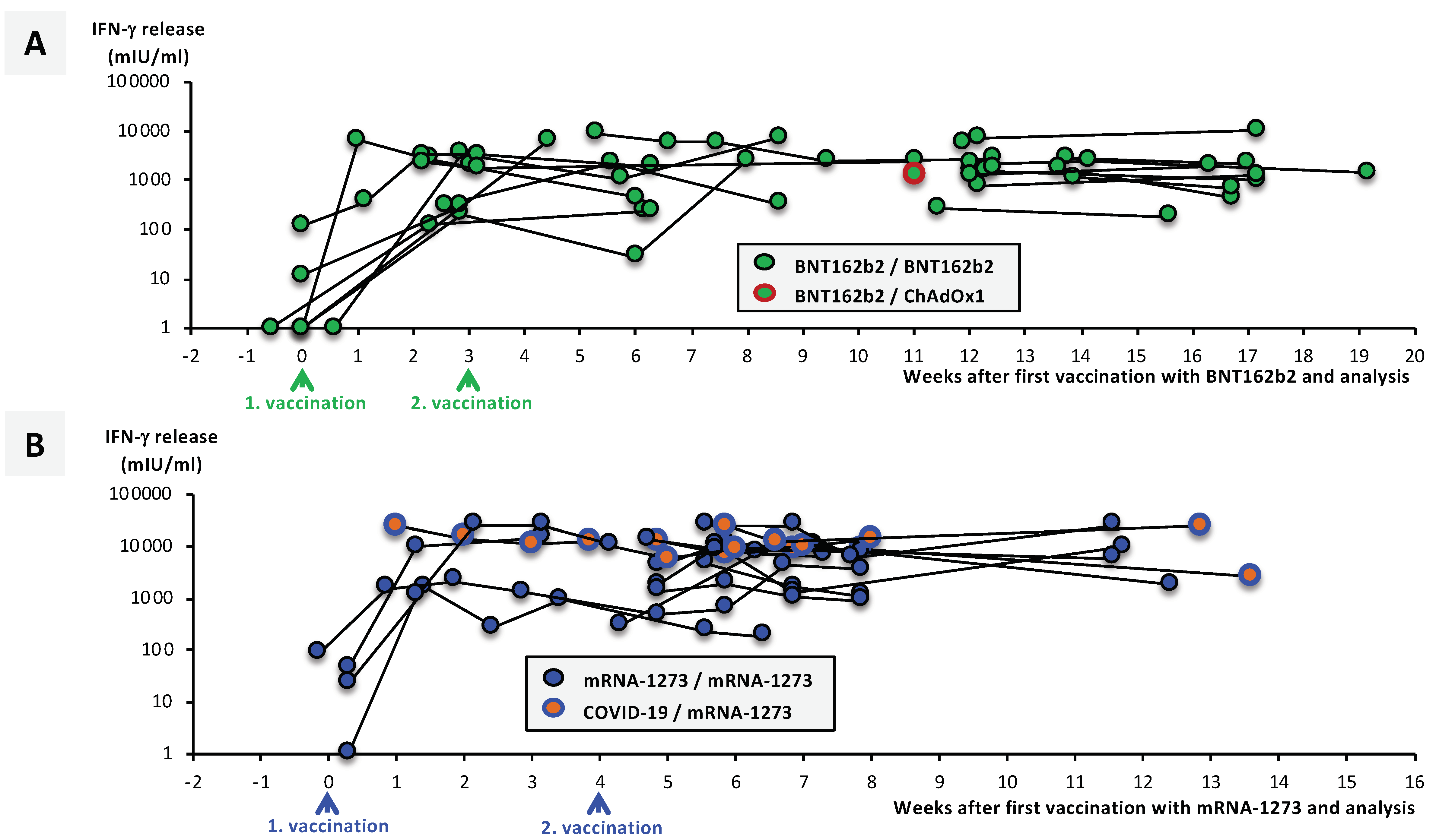
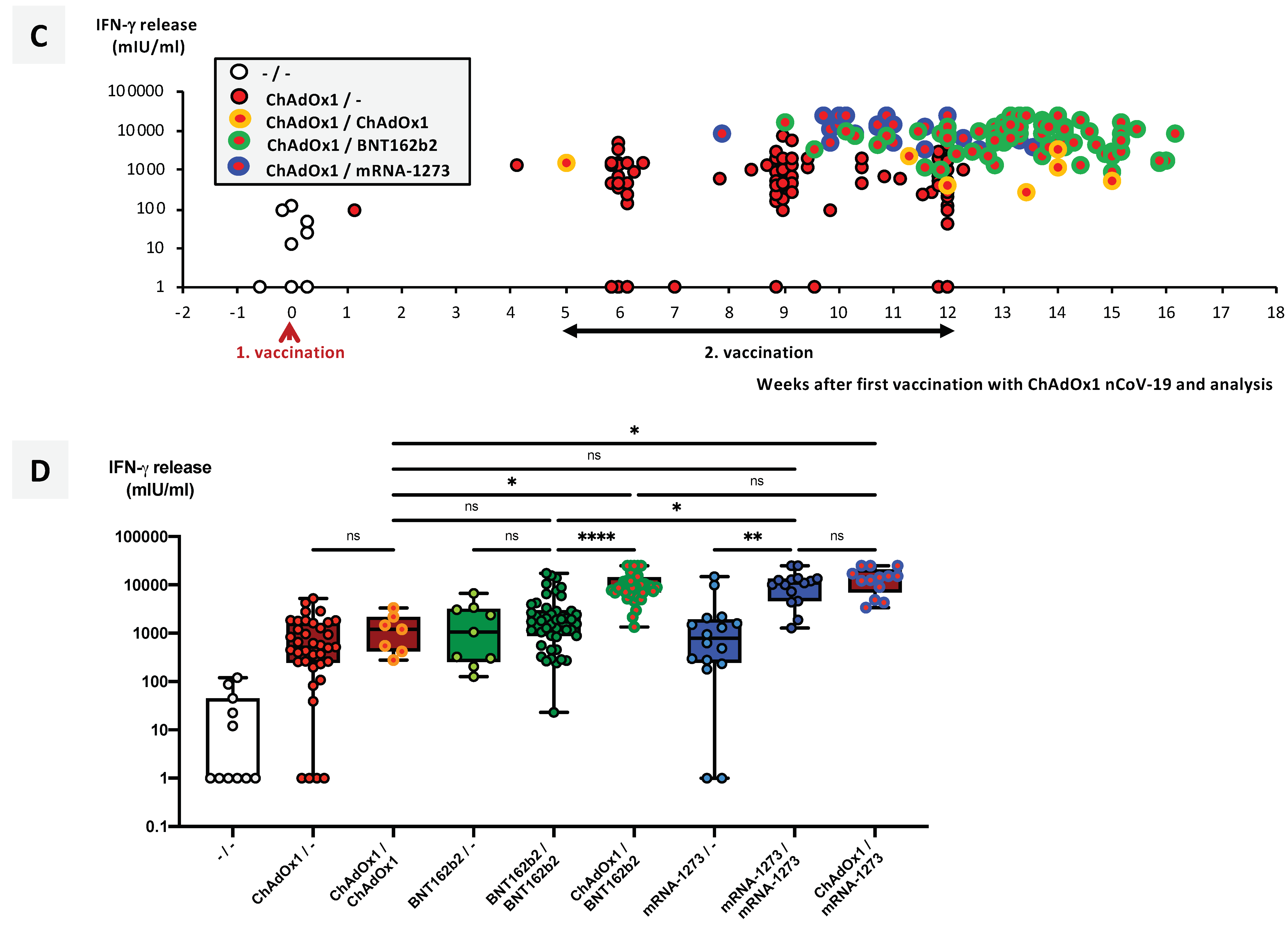
3.5. SARS-CoV-2-Specific T-Cell Responses Significantly Differ between Vaccination Groups
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Bernal, J.L.; Andrews, N.; Gower, C.; Stowe, J.; Robertson, C.; Tessier, E.; Simmons, R.; Cottrell, S.; Roberts, R.; O’Doherty, M.; et al. Early effectiveness of COVID-19 vaccination with BNT162b2 mRNA vaccine and ChAdOx1 adenovirus vector vaccine on symptomatic disease, hospitalisations and mortality in older adults in England. medRxiv 2021. [Google Scholar] [CrossRef]
- Jahrsdörfer, B.; Groß, R.; Seidel, A.; Wettstein, L.; Ludwig, C.; Schwarz, T.; Körper, S.; Rojewski, M.; Lotfi, R.; Weinstock, C.; et al. Characterization of the SARS-CoV-2 Neutralization Potential of COVID-19–Convalescent Donors. J. Immunol. 2021, 206, 2614–2622. [Google Scholar] [CrossRef]
- Jahrsdörfer, B.; Kroschel, J.; Ludwig, C.; Corman, V.M.; Schwarz, T.; Körper, S.; Rojewski, M.; Lotfi, R.; Weinstock, C.; Drosten, C.; et al. Independent side-by-side validation and comparison of four serological platforms for SARS-CoV-2 antibody testing. J. Infect. Dis. 2020, 223, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Körper, S.; Jahrsdörfer, B.; Corman, V.M.; Pilch, J.; Wuchter, P.; Blasczyk, R.; Müller, R.; Tonn, T.; Bakchoul, T.; Schäfer, R.; et al. Donors for SARS-CoV-2 Convalescent Plasma for a Controlled Clinical Trial: Donor Characteristics, Content and Time Course of SARS-CoV-2 Neutralizing Antibodies. Transfus. Med. Hemother. 2021, 48, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Arora, P.; Gross, R.; Seidel, A.; Hörnich, B.F.; Hahn, A.S.; Krüger, N.; Graichen, L.; Hofmann-Winkler, H.; Kempf, A.; et al. SARS-CoV-2 variants B.1.351 and P.1 escape from neutralizing antibodies. Cell 2021, 184, 2384–2393.e12. [Google Scholar] [CrossRef]
- Vanshylla, K.; Di Cristanziano, V.; Kleipass, F.; Dewald, F.; Schommers, P.; Gieselmann, L.; Gruell, H.; Schlotz, M.; Ercanoglu, M.S.; Stumpf, R.; et al. Kinetics and correlates of the neutralizing antibody response to SARS-CoV-2 infection in humans. Cell Host Microbe. 2021, 29, 849–1030. [Google Scholar] [CrossRef]
- Cohen, K.W.; Linderman, S.L.; Moodie, Z.; Czartoski, J.; Lai, L.; Mantus, G.; Norwood, C.; Nyhoff, L.E.; Edara, V.V.; Floyd, K.; et al. Longitudinal analysis shows durable and broad immune memory after SARS-CoV-2 infection with persisting antibody responses and memory B and T cells. medRxiv 2021. [Google Scholar] [CrossRef]
- Hurlburt, N.K.; Seydoux, E.; Wan, Y.; Edara, V.V.; Stuart, A.B.; Feng, J.; Suthar, M.S.; McGuire, A.T.; Stamatatos, L.; Pancera, M. Structural basis for potent neutralization of SARS-CoV-2 and role of antibody affinity maturation. Nat. Commun. 2020, 11, 5413. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 182, 429–446.e14. [Google Scholar] [CrossRef]
- Casciola-Rosen, L.; Thiemann, D.R.; Andrade, F.; Zambrano, M.I.T.; Hooper, J.E.; Leonard, E.K.; Spangler, J.B.; Cox, A.L.; Machamer, C.E.; Sauer, L.; et al. IgM autoantibodies recognizing ACE2 are associated with severe COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. RNA-Based COVID-19 Vaccine BNT162b2 Selected for a Pivotal Efficacy Study. medRxiv 2020. [Google Scholar] [CrossRef]
- Stamatatos, L.; Czartoski, J.; Wan, Y.H.; Homad, L.J.; Rubin, V.; Glantz, H.; Neradilek, M.; Seydoux, E.; Jennewein, M.F.; MacCamy, A.J.; et al. mRNA vaccination boosts cross-variant neutralizing antibodies elicited by SARS-CoV-2 infection. Science 2021, 372, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Libster, R.; Marc, G.P.; Wappner, D.; Coviello, S.; Bianchi, A.; Braem, V.; Esteban, I.; Caballero, M.T.; Wood, C.; Berrueta, M.; et al. Early High-Titer Plasma Therapy to Prevent Severe Covid-19 in Older Adults. N. Engl. J. Med. 2021, 384, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Simonovich, V.A.; Pratx, L.D.B.; Scibona, P.; Beruto, M.V.; Vallone, M.G.; Vázquez, C.; Savoy, N.; Giunta, D.H.; Pérez, L.G.; Sánchez, M.D.L.; et al. A Randomized Trial of Convalescent Plasma in Covid-19 Severe Pneumonia. N. Engl. J. Med. 2021, 384, 619–629. [Google Scholar] [CrossRef]
- Janiaud, P.; Axfors, C.; Schmitt, A.M.; Gloy, V.; Ebrahimi, F.; Hepprich, M.; Smith, E.R.; Haber, N.A.; Khanna, N.; Moher, D.; et al. Association of Convalescent Plasma Treatment With Clinical Outcomes in Patients With COVID-19: A Systematic Review and Meta-analysis. JAMA 2021, 325, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Greinacher, A.; Thiele, T.; Warkentin, T.E.; Weisser, K.; Kyrle, P.A.; Eichinger, S. Thrombotic Thrombocytopenia after ChAdOx1 nCov-19 Vaccination. N. Engl. J. Med. 2021, 384, 2092–2101. [Google Scholar] [CrossRef]
- He, Q.; Mao, Q.; An, C.; Zhang, J.; Gao, F.; Bian, L.; Li, C.; Liang, Z.; Xu, M.; Wang, J. Heterologous prime-boost: Breaking the protective immune response bottleneck of COVID-19 vaccine candidates. Emerg. Microbes Infect. 2021, 10, 629–637. [Google Scholar] [CrossRef]
- Shaw, R.H.; Stuart, A.; Greenland, M.; Liu, X.; Van-Tam, J.S.N.; Snape, M.D. Heterologous prime-boost COVID-19 vaccination: Initial reactogenicity data. Lancet 2021, 397, 2043–2046. [Google Scholar] [CrossRef]
- Hillus, D.; Schwarz, T.; Tober-Lau, P.; Hastor, H.; Thibeault, C.; Kasper, S.; Helbig, E.T.; Lippert, L.J.; Tscheak, P.; Schmidt, M.L. Safety, reactogenicity, and immunogenicity of homologous and heterologous prime-boost immunisation with ChAdOx1-nCoV19 and BNT162b2: A prospective cohort study. medRxiv 2021. [Google Scholar] [CrossRef]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of ChAdOx1 nCoV-19 (AZD1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, A.; Di Lauria, N.; Maggi, L.; Salvati, L.; Vanni, A.; Capone, M.; Lamacchia, G.; Mantengoli, E.; Spinicci, M.; Zammarchi, L.; et al. First-dose mRNA vaccination is sufficient to reactivate immunological memory to SARS-CoV-2 in recovered COVID-19 subjects. J. Clin. Investig. 2021. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort Code | A | B | C | D | E | F | G | H | I |
|---|---|---|---|---|---|---|---|---|---|
| Vaccine Type/ COVID-19 Convalescence | BNT162b2/ BNT162b2 | BNT162b2/ BNT162b2 COVID-19-Naive | BNT162b2/ BNT162b2 COVID-19-Convalescent | mRNA-1273/ mRNA-1273 | ChAdOx1/- | ChAdOx1/ BNT162b2 | ChAdOx2/ mRNA-1273 | ChAdOx1/ ChAdOx1 | Unvaccinated Reference Cohort COVID-19-Convalescent |
| Number of Individuals | 21 | 15 | 25 | 13 | 29 | 26 | 10 | 5 | 162 |
| Number Females (%) | 17 (81.0%) | 12 (80.0%) | 21 (84.0%) | 8 (61.5%) | 25 (86.2%) | 2 (7.7%) | 7 (70.0%) | 3 (60.0%) | 76 (46.9%) |
| Number Males (%) | 4 (19.0%) | 3 (20.0%) | 4 (16.0%) | 5 (38.5%) | 4 (13.8%) | 24 (92.3%) | 3 (30.0%) | 2 (40.0%) | 86 (53.1%) |
| Average Age (Range) | 45 (26–65) | 47 (26–64) | 46 (21–73) | 51 (34–61) | 45 (24–64) | 44 (22–64) | 33 (21–47) | 53 (46–63) | 32 (19–61) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabricius, D.; Ludwig, C.; Scholz, J.; Rode, I.; Tsamadou, C.; Jacobsen, E.-M.; Winkelmann, M.; Grempels, A.; Lotfi, R.; Janda, A.; et al. mRNA Vaccines Enhance Neutralizing Immunity against SARS-CoV-2 Variants in Convalescent and ChAdOx1-Primed Subjects. Vaccines 2021, 9, 918. https://doi.org/10.3390/vaccines9080918
Fabricius D, Ludwig C, Scholz J, Rode I, Tsamadou C, Jacobsen E-M, Winkelmann M, Grempels A, Lotfi R, Janda A, et al. mRNA Vaccines Enhance Neutralizing Immunity against SARS-CoV-2 Variants in Convalescent and ChAdOx1-Primed Subjects. Vaccines. 2021; 9(8):918. https://doi.org/10.3390/vaccines9080918
Chicago/Turabian StyleFabricius, Dorit, Carolin Ludwig, Judith Scholz, Immanuel Rode, Chrysanthi Tsamadou, Eva-Maria Jacobsen, Martina Winkelmann, Aline Grempels, Ramin Lotfi, Aleš Janda, and et al. 2021. "mRNA Vaccines Enhance Neutralizing Immunity against SARS-CoV-2 Variants in Convalescent and ChAdOx1-Primed Subjects" Vaccines 9, no. 8: 918. https://doi.org/10.3390/vaccines9080918
APA StyleFabricius, D., Ludwig, C., Scholz, J., Rode, I., Tsamadou, C., Jacobsen, E.-M., Winkelmann, M., Grempels, A., Lotfi, R., Janda, A., Körper, S., Adler, G., Debatin, K.-M., Schrezenmeier, H., & Jahrsdörfer, B. (2021). mRNA Vaccines Enhance Neutralizing Immunity against SARS-CoV-2 Variants in Convalescent and ChAdOx1-Primed Subjects. Vaccines, 9(8), 918. https://doi.org/10.3390/vaccines9080918