
The New Status of Parasitic Diseases in the COVID-19 Pandemic—Risk Factors or Protective Agents?

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. COVID-19 Symptoms
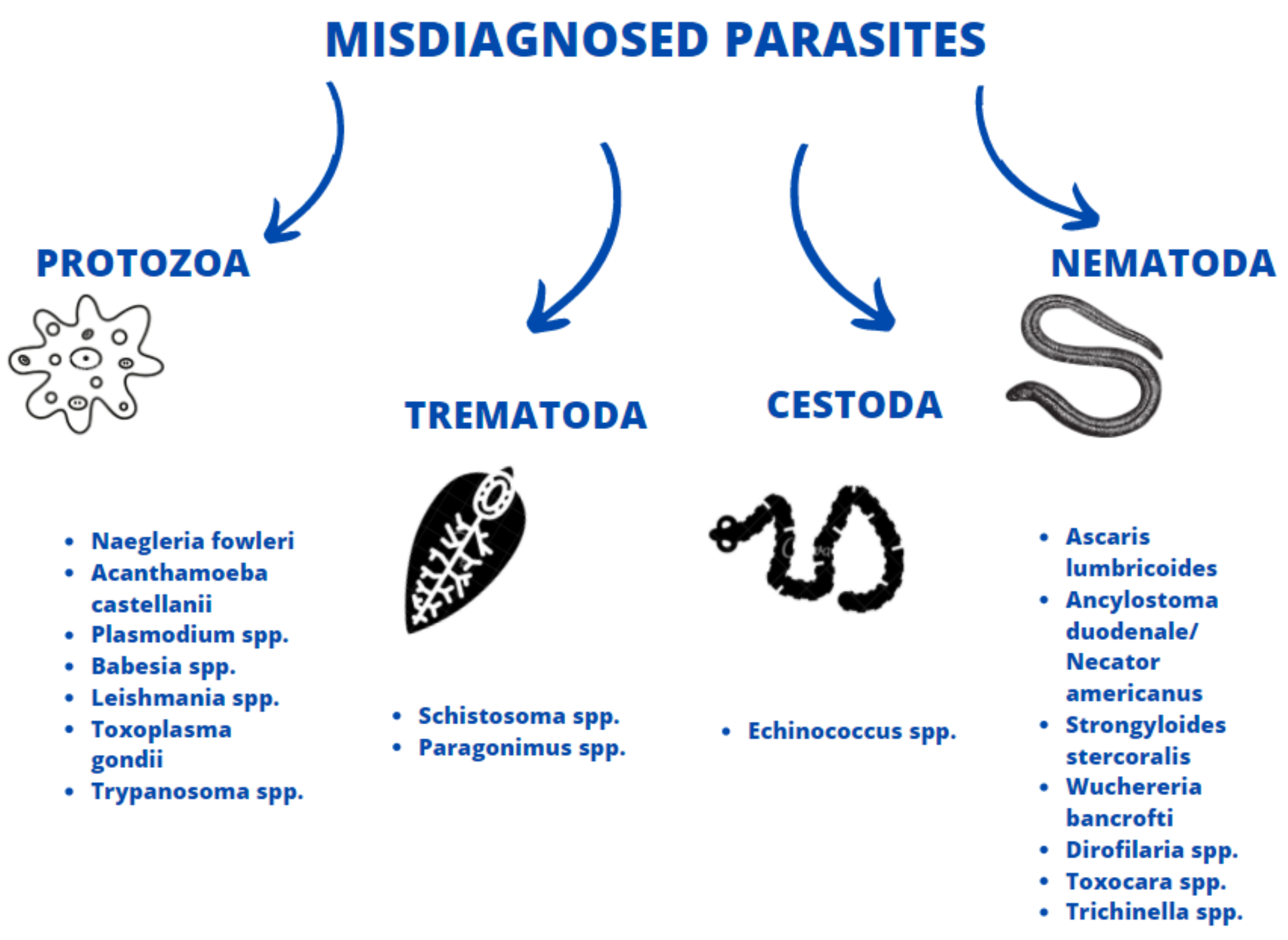
3. Comparison with Symptoms of Selected Parasitoses
3.1. Protozoans
3.2. Trematodes
3.3. Cestodes
3.4. Nematodes
4. Increased Risk of COVID Infection and Vaccination Efficacy
5. The COVID-19 Pandemic in Relation to Antiparasitic Prevention Programs and Parasitological Diagnostics
6. Can Parasites Protect Us from the COVID-19?
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsieh, S. Parasitic Disorders; Williamd&Wilkins: Baltimore, MD, USA, 1988. [Google Scholar]
- Bradbury, R.S.; Piedrafita, D.; Greenhill, A.; Mahanty, S. Will helminth co-infection modulate COVID-19 severity in endemic regions? Nat. Rev. Immunol. 2020, 20, 342. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, A.; Ardakani, H.M. Helminth infections and immunosenescence: The friend of my enemy. Exp. Gerontol. 2020, 133, 110852. [Google Scholar] [CrossRef]
- Abdoli, A. Helminths and COVID-19 Co-Infections: A Neglected Critical Challenge. Acs Pharm. Transl. Sci. 2020, 3, 1039–1041. [Google Scholar] [CrossRef]
- Fonte, L.; Acosta, A.; Sarmiento, M.E.; Ginori, M.; García, G.; Norazmi, M.N. COVID-19 Lethality in Sub-Saharan Africa and Helminth Immune Modulation. Front. Immunol. 2020, 11, 574910. [Google Scholar] [CrossRef]
- Mohamed, M.F.H.; Mohamed, S.F.; Yousaf, Z.; Kohla, S.; Howady, F.; Imam, Y. COVID-19 unfolding filariasis: The first case of SARS-CoV-2 and Wuchereria bancrofti coinfection. Plos Negl. Trop. Dis. 2020, 14, e0008853. [Google Scholar] [CrossRef] [PubMed]
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef]
- CDC. Symptoms of Coronavirus Disease 2019 (COVID-19). 2020. Available online: https://www.cdc.gov/coronavirus/2019-ncov/about/symptoms.html (accessed on 9 November 2020).
- Pan, L.; Mu, M.; Yang, P.; Sun, Y.; Wang, R.; Yan, J.; Li, P.; Hu, B.; Wang, J.; Hu, C.; et al. Clinical Characteristics of COVID-19 Patients With Digestive Symptoms in Hubei, China: A Descriptive, Cross-Sectional, Multicenter Study. Am. J. Gastroenterol. 2020, 115, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Pascarella, G.; Strumia, A.; Piliego, C.; Bruno, F.; Del Buono, R.; Costa, F.; Scarlata, S.; Agrò, F.E. COVID-19 diagnosis and management: A comprehensive review. J. Intern. Med. 2020, 288, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Confalonieri, M.; Salton, F.; Fabiano, F. Acute respiratory distress syndrome. Eur. Respir. Rev. 2017, 26, 160116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issa, R. Tropical parasitic lung diseases. Int. J. Pharm. Pharm. 2015, 7, 2–12. [Google Scholar]
- Siddiqui, R.; Khan, N.A. Biology and pathogenesis of Acanthamoeba. Parasit Vectors 2012, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocięcka, W. Clinical parasitology. In Repertory of Selected Parasitic and Tropical Diseases; Scientific Publisher Medical University: Poznań, Poland, 2016. [Google Scholar]
- Jahangeer, M.; Mahmood, Z.; Munir, N.; Waraich, U.; Tahir, I.M.; Akram, M.; Shah, S.M.A.; Zulfqar, A.; Zainab, R. Naegleria fowleri: Sources of infection, pathophysiology, diagnosis, and management; a review. Clin. Exp. Pharmacol. Physiol. 2020, 47, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, R.; Khan, N.A. Primary amoebic meningoencephalitis caused by Naegleria fowleri: An old enemy presenting new challenges. PLoS Negl. Trop. Dis. 2014, 8, e3017. [Google Scholar] [CrossRef]
- Attia, J.; Hatala, R.; Cook, D.J.; Wong, J.G. Does This Adult Patient Have Acute Meningitis? JAMA 1999, 282, 175–181. [Google Scholar] [CrossRef]
- Van De Beek, D.; De Gans, J.; Spanjaard, L.; Weisfelt, M.; Reitsma, J.B.; Vermeulen, M. Clinical Features and Prognostic Factors in Adults with Bacterial Meningitis. New Engl. J. Med. 2004, 351, 1849–1859. [Google Scholar] [CrossRef] [Green Version]
- Kot, K.; Łanocha-Arendarczyk, N.A.; Kosik-Bogacka, D.I. Amoebas from the genus Acanthamoeba and their pathogenic properties. Ann. Parasitol. 2018, 64, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Maeda, T.; Kusakabe, S.; Arita, H.; Yagita, K.; Morii, E.; Kanakura, Y. A case report of granulomatous amoebic encephalitis by Group 1 Acanthamoeba genotype T18 diagnosed by the combination of morphological examination and genetic analysis. Diagn. Pathol. 2018, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- Manguin, S.; Bangs, M.; Pothikasikorn, J.; Chareonviriyaphap, T. Review on global co-transmission of human Plasmodium species and Wuchereria bancrofti by Anopheles mosquitoes. Infect. Genet. Evol. 2010, 10, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Talapko, J.; Škrlec, I.; Alebić, T.; Jukić, M.; Včev, A. Malaria: The Past and the Present. Microorganisms. 2019, 7, 179. [Google Scholar] [CrossRef] [Green Version]
- Jamil, M.D.; Kar, G.; Biswas, S.K.; Kaushik, P. Clinical manifestations of falciparum malaria in relation to parasitemia level in Southern Assam. J. Assoc. Phys. India 2009, 57, 4. [Google Scholar]
- Cabral, P.H.O.; Andrade, S.D.; Alecrim, W.D.; Alecrim, M.G.C.; Lacerda, M.V.G. Malaria and sickle cell anemia: Report of complications and clinical management of three patients in a highly endemic area FOR Plasmodium vivax malaria in the Brazilian Amazon. Case Rep. Clin. Pract. Rev. 2006, 7, 220–223. [Google Scholar]
- Sanklecha, M.; Mehta, N.; Bagban, H. Varied presentation of complicated falciparum malaria in a family. Indian Pediatrics 2012, 49, 413–414. [Google Scholar]
- Hogan, A.B.; Jewell, B.L.; Sherrard-Smith, E.; Vesga, J.F.; Watson, O.J.; Whittaker, C.; Hamlet, A.; A Smith, J.; Winskill, P.; Verity, R.; et al. Potential impact of the COVID-19 pandemic on HIV, tuberculosis, and malaria in low-income and middle-income countries: A modelling study. Lancet Glob. Health 2020, 8, e1132–e1141. [Google Scholar] [CrossRef]
- Cheepsattayakorn, A.; Cheepsattayakorn, R. Parasitic pneumonia and lung involvement. Biomed. Res. Int. 2014, 2014, 874021. [Google Scholar] [CrossRef] [Green Version]
- Ord, R.L.; Lobo, C.A. Human Babesiosis: Pathogens, Prevalence, Diagnosis, and Treatment. Curr. Clin. Microbiol. Rep. 2015, 2, 173–181. [Google Scholar] [CrossRef]
- Hatcher, J.C.; Greenberg, P.D.; Antique, J.; Jimenez-Lucho, V.E. Severe Babesiosis in Long Island: Review of 34 Cases and Their Complications. Clin. Infect. Dis. 2001, 32, 1117–1125. [Google Scholar] [CrossRef]
- Hussein, M.I.H.; Albashir, A.A.D.; Elawad, O.A.M.A.; Homeida, A. Malaria and COVID-19: Unmasking their ties. Malar. J. 2020, 19, 457. [Google Scholar] [CrossRef]
- Alexandrino-De-Oliveira, P.; Santos-Oliveira, J.R.; Dorval, M.E.C.; Da-Costa, F.D.C.B.; Pereira, G.R.O.L.; Da Cunha, R.V.; Paniago, A.M.M.; Da-Cruz, A.M. HIV/AIDS-associated visceral leishmaniasis in patients from an endemic area in Central-west Brazil. Memórias Inst. Oswaldo Cruz 2010, 105, 692–697. [Google Scholar] [CrossRef] [Green Version]
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A review. F1000Research 2017, 6, 750. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.G.; Loaiza, J.R. American trypanosomiasis, or Chagas disease, in Panama: A chronological synopsis of ecological and epidemiological research. Parasites Vectors 2017, 10, 459. [Google Scholar] [CrossRef] [PubMed]
- Luintel, A.; Lowe, P.; Cooper, A.; MacLeod, A.; Büscher, P.; Brooks, T.; Brown, M. Case of Nigeria-Acquired Human African Trypanosomiasis in United Kingdom, 2016. Emerg. Infect. Dis. 2017, 23, 1225–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braakman, H.M.; Van De Molengraft, F.J.; Hubert, W.W.; Boerman, D.H. Lethal African trypanosomiasis in a traveler: MRI and neuropathology. Neurology 2006, 66, 1094–1096. [Google Scholar] [CrossRef] [PubMed]
- Butler, N.J.; Furtado, J.M.; Winthrop, K.L.; Smith, J.R. Ocular toxoplasmosis II: Clinical features, pathology and management. Clin. Exp. Ophthalmol. 2013, 41, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Basavaraju, A. Toxoplasmosis in HIV infection: An overview. Trop Parasitol. 2016, 6, 129–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, N.; Misra, B. Toxoplasma seropositivity and depression: A case report. BMC Psychiatry 2004, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E.; Rozenman, J.; Perelman, M. Pulmonary manifestations of early schistosome infection among nonimmune travelers. Am. J. Med. 2000, 109, 718–722. [Google Scholar] [CrossRef]
- Nelwan, M.L. Schistosomiasis: Life Cycle, Diagnosis, and Control. Curr. Ther. Res. 2019, 91, 5–9. [Google Scholar] [CrossRef]
- Niemann, T.; Marti, H.; Duhnsen, S.; Bongartz, G. Pulmonary Schistosomiasis—Imaging Features. J. Radiol. Case Rep. 2010, 4, 37–43. [Google Scholar] [CrossRef]
- Singh, T.S.; Sugiyama, H.; Rangsiruji, A. Paragonimus & paragonimiasis in India. Indian J. Med. Res. 2012, 136, 192–204. [Google Scholar]
- Kalhan, S.; Sharma, P.; Sharma, S.; Kakria, N.; Dudani, S.; Gupta, A. Paragonimus westermani infection in lung: A confounding diagnostic entity. Lung India 2015, 32, 265–267. [Google Scholar] [CrossRef]
- Peng, W.; Zhou, X.; Gasser, R.B. Ascaris egg profiles in human faeces: Biological and epidemiological implications. Parasitology 2003, 127, 283–290. [Google Scholar] [CrossRef]
- Lamberton, P.H.L.; Jourdan, P.M. Human Ascariasis: Diagnostics Update. Curr. Trop. Med. Rep. 2015, 2, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooker, S.; Bethony, J.; Hotez, P. Human Hookworm Infection in the 21st Century. Adv. Parasitol. 2004, 58, 197–288. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Cheng, M.; Zhang, J.; Chen, G.; Liu, D.; Liu, Y.; Liu, H. Hookworm Infection Caused Acute Intestinal Bleeding Diagnosed by Capsule: A Case Report and Literature Review. Korean J. Parasitol. 2017, 55, 417–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimino, R.O.; Fleitas, P.; Fernández, M.; Echazú, A.; Juarez, M.; Floridia-Yapur, N.; Cajal, P.; Seijo, A.; Abril, M.; Weinberg, D.; et al. Seroprevalence of the Strongyloides stercoralis Infection in Humans from Yungas Rainforest and Gran Chaco Region from Argentina and Bolivia. Pathogens 2020, 9, 394. [Google Scholar] [CrossRef]
- Dogan, C.; Gayaf, M.; Ozsoz, A.; Sahin, B.; Aksel, N.; Karasu, I.; Aydogdu, Z.; Turgay, N. Pulmonary Strongyloides stercoralis infection. Respir. Med. Case Rep. 2014, 11, 12–15. [Google Scholar] [CrossRef] [Green Version]
- Karanam, L.S.K.; Basavraj, G.K.; Papireddy, C.K.R. Strongyloides stercoralis Hyper infection Syndrome. Indian J. Surg. 2020, 12, 1–5. [Google Scholar] [CrossRef]
- Marchese, V.; Crosato, V.; Gulletta, M.; Castelnuovo, F.; Cristini, G.; Matteelli, A.; Castelli, F. Strongyloides infection manifested during immunosuppressive therapy for SARS-CoV-2 pneumonia. Infection 2020, 1–4. [Google Scholar] [CrossRef]
- Lier, A.J.; Tuan, J.J.; Davis, M.W.; Paulson, N.; McManus, D.; Campbell, S.; Peaper, D.R.; Topal, J.E. Case Report: Disseminated Strongyloidiasis in a Patient with COVID-19. Am. J. Trop. Med. Hyg. 2020, 103, 1590–1592. [Google Scholar] [CrossRef]
- Stauffer, W.M.; Alpern, J.D.; Walker, P.F. COVID-19 and Dexamethasone: A Potential Strategy to Avoid Steroid-Related Strongyloides Hyperinfection. JAMA 2020, 324, 623. [Google Scholar] [CrossRef]
- Mullerpattan, J.B.; Udwadia, Z.F.; Udwadia, F.E. Tropical pulmonary eosinophilia—A review. Indian J. Med Res. 2013, 138, 295–302. [Google Scholar]
- Sandhu, M.; Mukhopadhyay, S.; Sharma, S.K. Tropical pulmonary eosinophilia: A comparative evaluation of plain chest radiography and computed tomography. Australas. Radiol. 1996, 40, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Mulanovich, E.A.; Mulanovich, V.E.; Rolston, K.V. A case of Dirofilaria pulmonary infection coexisting with lung cancer. J. Infect. 2008, 56, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Simón, F.; Siles-Lucas, M.; Morchón, R.; González-Miguel, J.; Mellado, I.; Carretón, E.; Montoya-Alonso, J.A. Human and Animal Dirofilariasis: The Emergence of a Zoonotic Mosaic. Clin. Microbiol. Rev. 2012, 25, 507–544. [Google Scholar] [CrossRef] [Green Version]
- Mazur-Melewska, K.; Jończyk-Potoczna, K.; Kemnitz, P.; Mania, A.; Figlerowicz, M.; Służewski, W. Pulmonary presentation of Toxocara sp. infection in children. Pneumonol. Alergol. Pol. 2015, 83, 250–255. [Google Scholar] [CrossRef] [Green Version]
- Sakai, S.; Shida, Y.; Takahashi, N.; Yabuuchi, H.; Soeda, H.; Okafuji, T.; Hatakenaka, M.; Honda, H. Pulmonary lesions associated with visceral larva migrans due to Ascaris suum or Toxocara canis: Imaging of six cases. AJR Am. J. Roentgenol. 2006, 186, 1697–1702. [Google Scholar] [CrossRef]
- Roig, J.; Romeu, J.; Riera, C.; Texido, A.; Domingo, C.; Morera, J. Acute Eosinophilic Pneumonia due to Toxocariasis with Bronchoalveolar Lavage Findings. Chest 1992, 102, 294–296. [Google Scholar] [CrossRef]
- Taki, M.; Sugio, Y.; Furuta, K.; Nakaji, H.; Morita, K.; Ikeue, T.; Sugita, T.; Nishiyama, H. A case of toxocariasis that induced eosinophilic pneumonia with mechanical ventilation. Jpn. J. Chest Dis. 2013, 72, 1015–1020. [Google Scholar]
- Demirci, M.; Unlu, M.; Fidan, F.; Kaya, S. Eosinophilic pneumonia due to toxocariasis: An adult case report. Turk. Parazitol Derg 2012, 36, 258–259. [Google Scholar] [CrossRef]
- Despommier, D. Toxocariasis: Clinical Aspects, Epidemiology, Medical Ecology, and Molecular Aspects. Clin. Microbiol. Rev. 2003, 16, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Despommier, D. How Does Trichinella spiralis Make Itself at Home? Parasitol. Today 1998, 14, 318–323. [Google Scholar] [CrossRef]
- Bruschi, F.; Murrell, K.D. New aspects of human trichinellosis: The impact of new Trichinella species. Postgrad. Med. J. 2002, 78, 15–22. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Gutman, J.R.; Lucchi, N.W.; Cantey, P.T.; Steinhardt, L.C.; Samuels, A.M.; Kamb, M.L.; Kapella, B.K.; McElroy, P.D.; Udhayakumar, V.; Lindblade, K.A. Malaria and Parasitic Neglected Tropical Diseases: Potential Syndemics with COVID-19? Am. J. Trop. Med. Hyg. 2020, 103, 572–577. [Google Scholar] [CrossRef]
- Allen, J.E.; Maizels, R.M. Diversity and dialogue in immunity to helminths. Nat. Rev. Immunol. 2011, 11, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Garvin, M.R.; Alvarez, C.; Miller, J.I.; Prates, E.T.; Walker, A.M.; Amos, B.K.; Mast, A.E.; Justice, A.; Aronow, B.; Jacobson, D.A. A mechanistic model and therapeutic interventions for COVID-19 involving a RAS-mediated bradykinin storm. eLife 2020, 9, e59177. [Google Scholar] [CrossRef] [PubMed]
- Vale, A.J.M.; Fernandes, A.C.L.; Guzen, F.P.; Pinheiro, F.I.; de Azevedo, E.P.; Cobucci, R.N. Susceptibility to COVID-19 in Pregnancy, Labor, and Postpartum Period: Immune System, Vertical Transmission, and Breastfeeding. Front. Glob. Women’s Health 2021, 2, 602572. [Google Scholar] [CrossRef]
- Wastnedge, E.A.N.; Reynolds, R.M.; Van Boeckel, S.R.; Stock, S.J.; Denison, F.C.; Maybin, J.A.; Critchley, H.O.D. Pregnancy and COVID-19. Physiol. Rev. 2021, 101, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Halle-Ekane, G.; Konje, J.C. Intestinal parasitic infections in pregnancy—A review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 254, 59–63. [Google Scholar] [CrossRef]
- Adepoju, P. NTDs in the time of COVID-19. Lancet Microbe 2020, 1, e244. [Google Scholar] [CrossRef]
- Dyer, O. African malaria deaths set to dwarf covid-19 fatalities as pandemic hits control efforts, WHO warns. BMJ 2020, 371. [Google Scholar] [CrossRef]
- Sherrard-Smith, E.; Hogan, A.B.; Hamlet, A.; Watson, O.J.; Whittaker, C.; Winskill, P.; Ali, F.; Mohammad, A.B.; Uhomoibhi, P.; Maikore, I.; et al. The potential public health consequences of COVID-19 on malaria in Africa. Nat. Med. 2020, 26, 1411–1416. [Google Scholar] [CrossRef] [PubMed]
- Malizia, V.; Giardina, F.; Vegvari, C.; Bajaj, S.; McRae-McKee, K.; Anderson, R.M.; de Vlas, S.J.; E Coffeng, L. Modelling the impact of COVID-19-related control programme interruptions on progress towards the WHO 2030 target for soil-transmitted helminths. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 253–260. [Google Scholar] [CrossRef]
- Napoli, P.E.; Nioi, M. Global Spread of Coronavirus Disease 2019 and Malaria: An Epidemiological Paradox in the Early Stage of A Pandemic. J. Clin. Med. 2020, 9, 1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://covid19.who.int/ (accessed on 4 December 2020).
- Njenga, M.K.; Dawa, J.; Nanyingi, M.; Gachohi, J.; Ngere, I.; Letko, M.; Otieno, C.F.; Gunn, B.M.; Osoro, E. Why is There Low Morbidity and Mortality of COVID-19 in Africa? Am. J. Trop. Med. Hyg. 2020, 103, 564–569. [Google Scholar] [CrossRef]
- Ssebambulidde, K.; Segawa, I.; Abuga, K.M.; Nakate, V.; Kayiira, A.; Ellis, J.; Tugume, L.; Kiragga, A.N.; Meya, D.B. Parasites and their protection against COVID-19- Ecology or Immunology? MedRxiv 2020. [Google Scholar] [CrossRef]
- Maizels, R.M. Regulation of immunity and allergy by helminth parasites. Allergy 2020, 75, 524–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siles-Lucas, M.; González-Miguel, J.; Geller, R.; Sanjuan, R.; Pérez-Arévalo, J.; Martínez-Moreno, Á. Potential Influence of Helminth Molecules on COVID-19 Pathology. Trends Parasitol. 2021, 37, 11–14. [Google Scholar] [CrossRef]
- Schwartz, C.; Hams, E.; Fallon, P.G. Helminth Modulation of Lung Inflammation. Trends Parasitol. 2018, 34, 388–403. [Google Scholar] [CrossRef] [Green Version]
- Furze, R.C.; Hussell, T.; Selkirk, M.E. Amelioration of Influenza-Induced Pathology in Mice by Coinfection with Trichinella spiralis. Infect. Immun. 2006, 74, 1924–1932. [Google Scholar] [CrossRef] [Green Version]
- Scheer, S.; Krempl, C.; Kallfass, C.; Frey, S.; Jakob, T.; Mouahid, G.; Moné, H.; Schmitt-Gräff, A.; Staeheli, P.; Lamers, M.C.S. mansoni Bolsters Anti-Viral Immunity in the Murine Respiratory Tract. PLoS ONE 2014, 9, e112469. [Google Scholar] [CrossRef] [PubMed]
- Wamae, C.N. Mass Drug Administration and Worms Experience in Africa: Envisage Repurposing Ivermectin for SARS-COV-2. Am. J. Trop. Med. Hyg. 2020, 103, 10–11. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Głuchowska, K.; Dzieciątkowski, T.; Sędzikowska, A.; Zawistowska-Deniziak, A.; Młocicki, D. The New Status of Parasitic Diseases in the COVID-19 Pandemic—Risk Factors or Protective Agents? J. Clin. Med. 2021, 10, 2533. https://doi.org/10.3390/jcm10112533
Głuchowska K, Dzieciątkowski T, Sędzikowska A, Zawistowska-Deniziak A, Młocicki D. The New Status of Parasitic Diseases in the COVID-19 Pandemic—Risk Factors or Protective Agents? Journal of Clinical Medicine. 2021; 10(11):2533. https://doi.org/10.3390/jcm10112533
Chicago/Turabian StyleGłuchowska, Kinga, Tomasz Dzieciątkowski, Aleksandra Sędzikowska, Anna Zawistowska-Deniziak, and Daniel Młocicki. 2021. "The New Status of Parasitic Diseases in the COVID-19 Pandemic—Risk Factors or Protective Agents?" Journal of Clinical Medicine 10, no. 11: 2533. https://doi.org/10.3390/jcm10112533
APA StyleGłuchowska, K., Dzieciątkowski, T., Sędzikowska, A., Zawistowska-Deniziak, A., & Młocicki, D. (2021). The New Status of Parasitic Diseases in the COVID-19 Pandemic—Risk Factors or Protective Agents? Journal of Clinical Medicine, 10(11), 2533. https://doi.org/10.3390/jcm10112533