
Long-Term SARS-CoV-2 Specific Immunity Is Affected by the Severity of Initial COVID-19 and Patient Age

 ,
,  , ,
, ,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects and Sampling
2.2. T Cell ELISpot Assays for S- and M-Derived SARS-CoV-2 Peptides
2.3. SARS-CoV-2 Specific Antibody Detection
2.4. Cells and Virus
2.5. Virus Neutralization Assay
2.6. Statistics and Data Analysis
3. Results
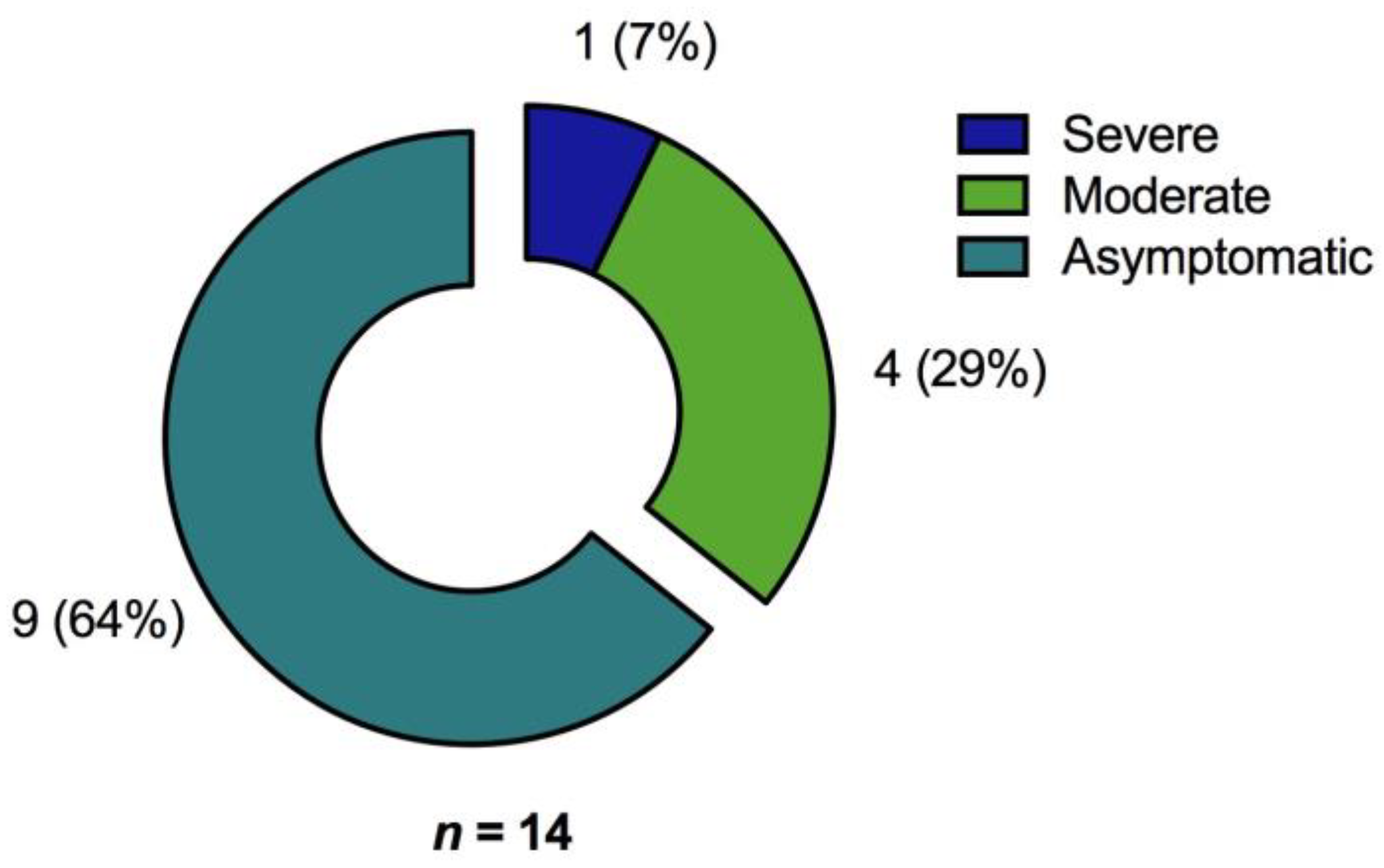
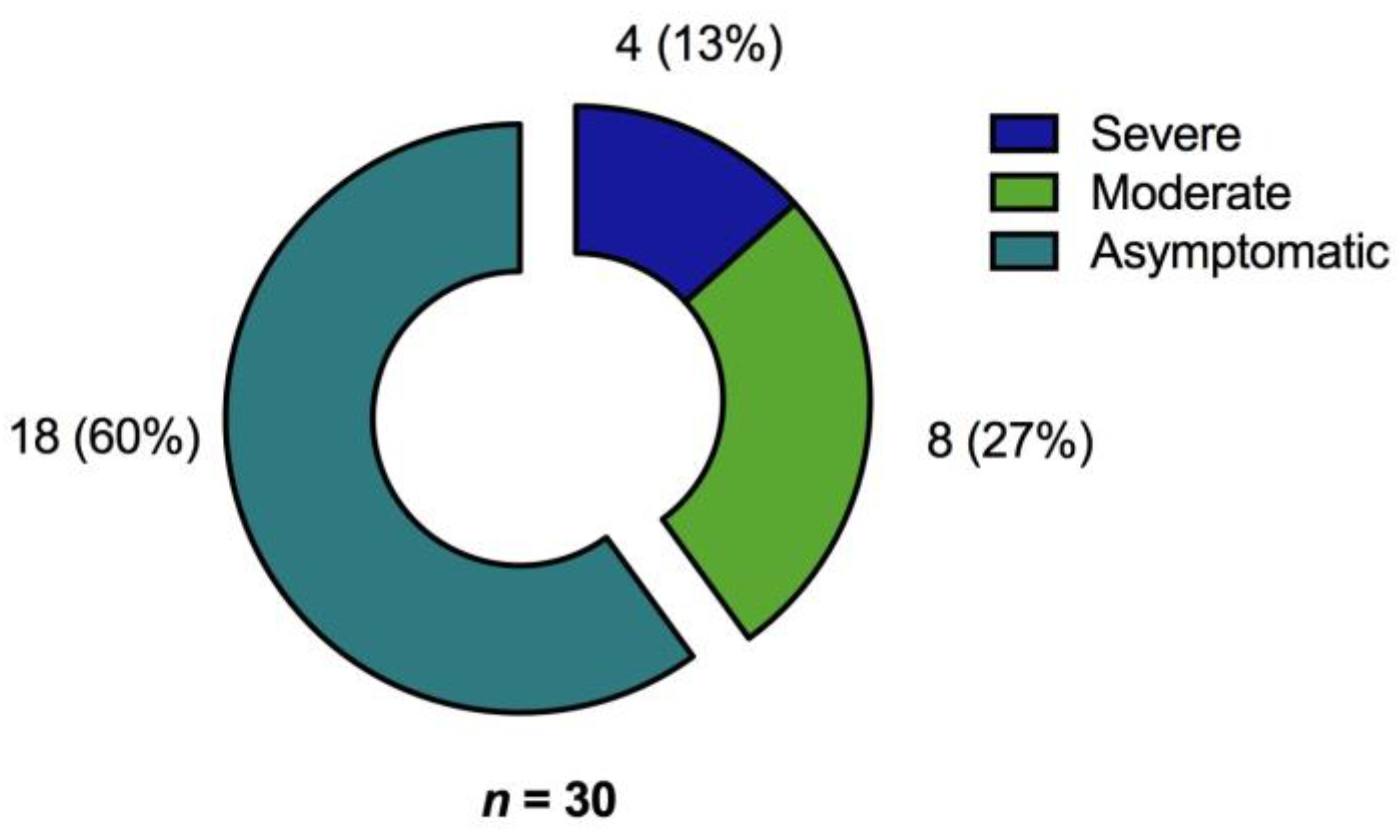
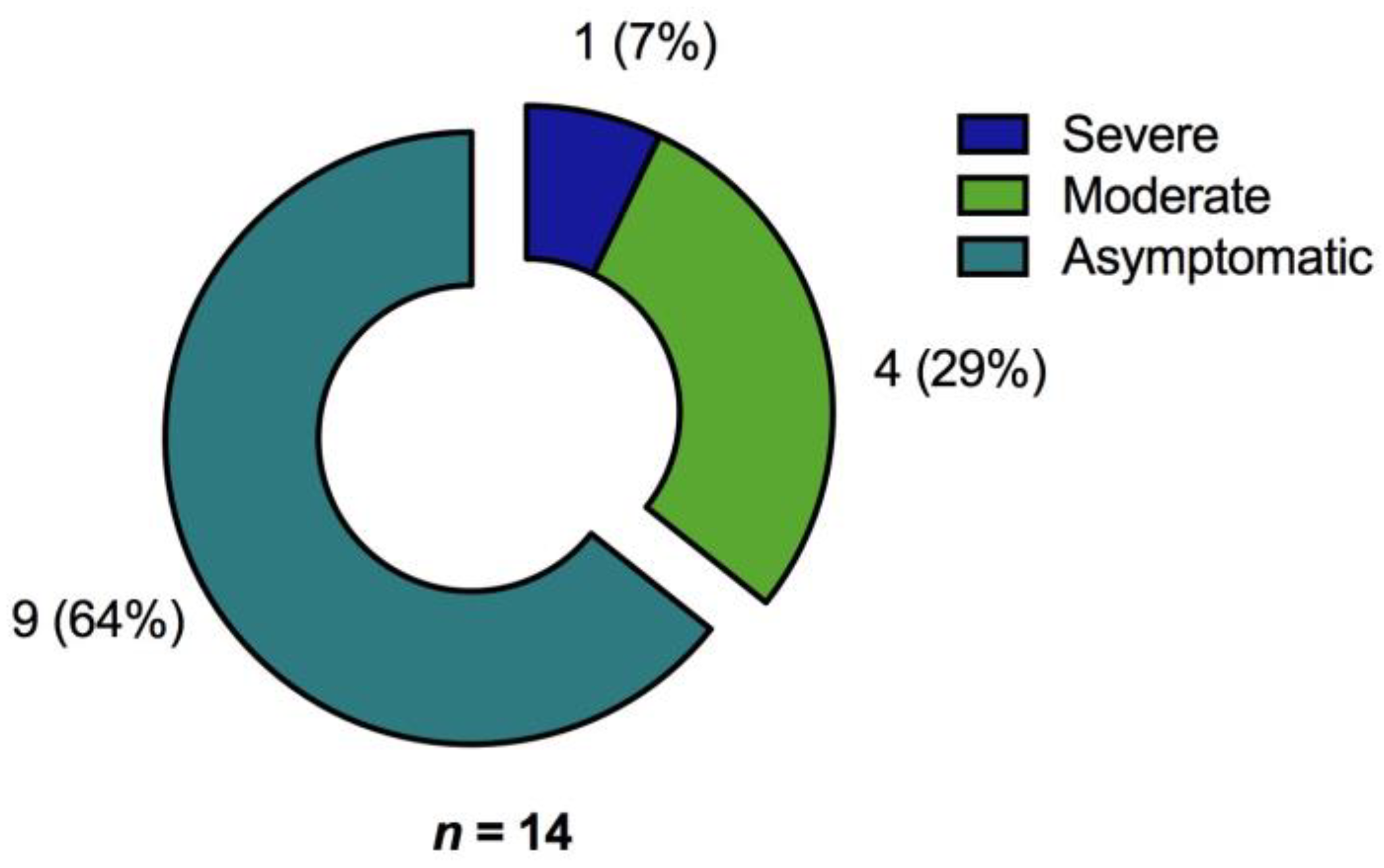
3.1. Participants
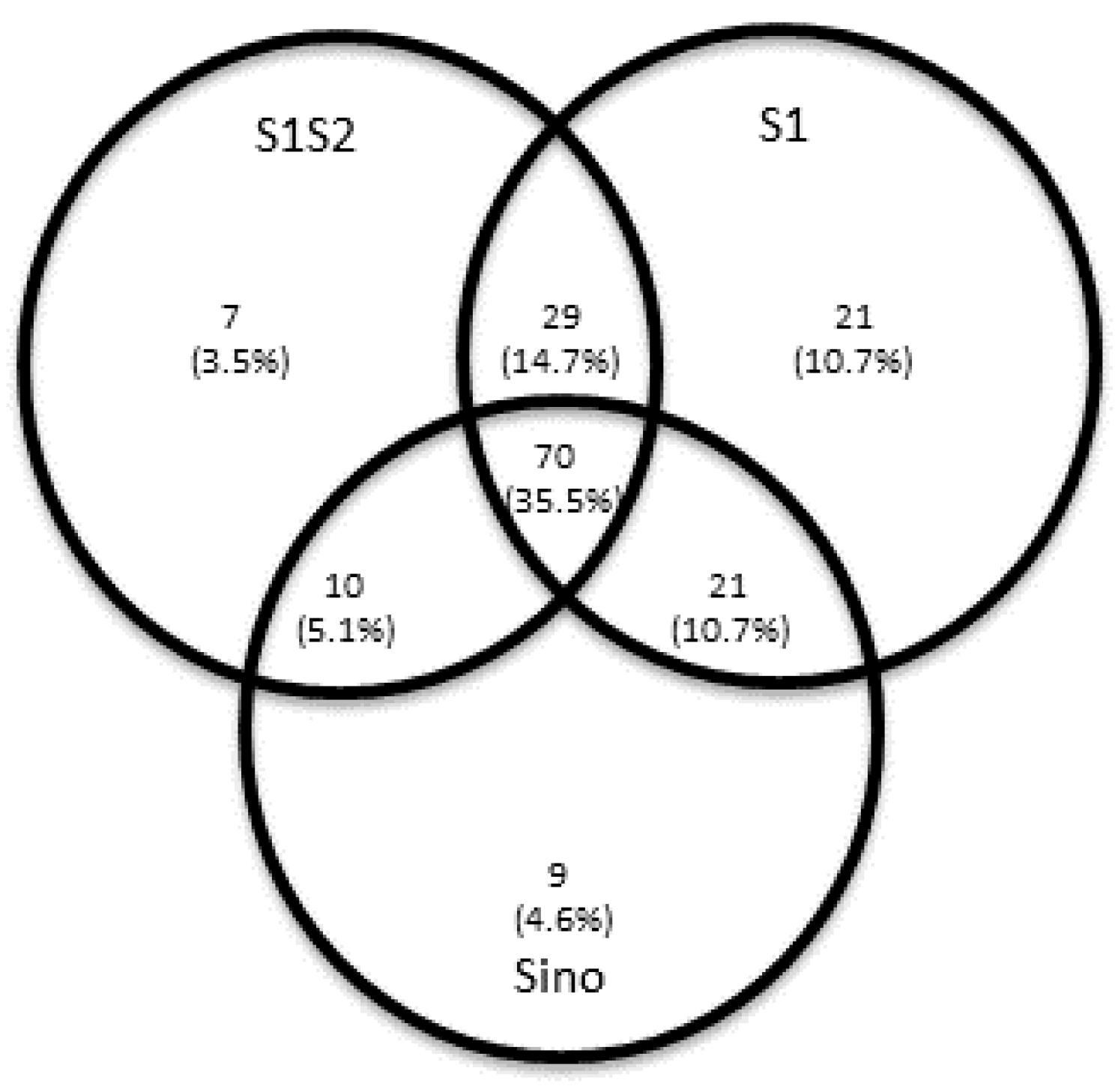
3.2. Simultaneous T Cell ELISpot Assays for S and M SARS-CoV-2 Peptides Increase the Probability of Detecting T Cell Immunity
3.2.1. Immunosuppression Is an Independent Risk Factor for Lack of Response to SARS-CoV-2 Antigen-Specific Stimulation in ELISpot Analyses
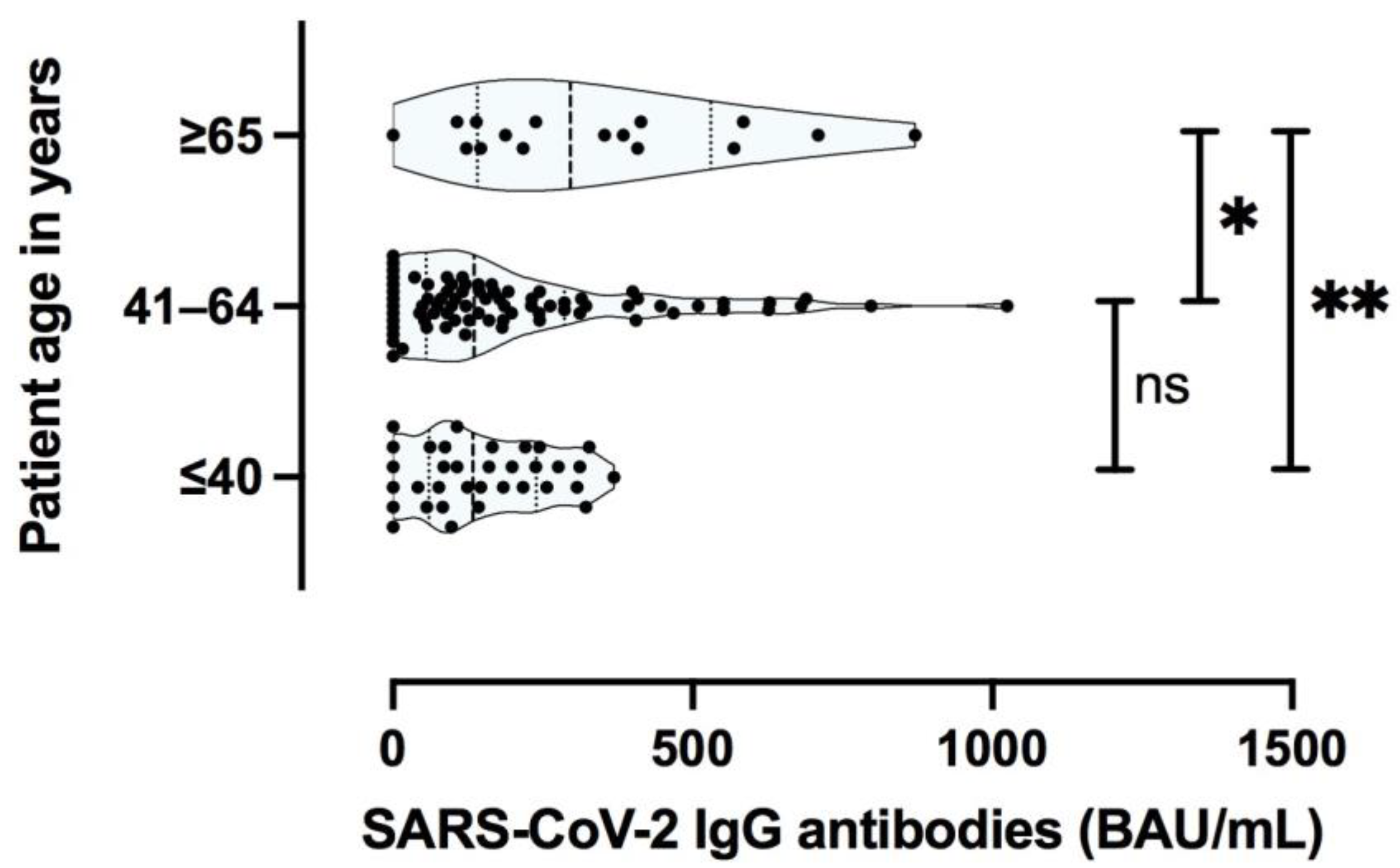
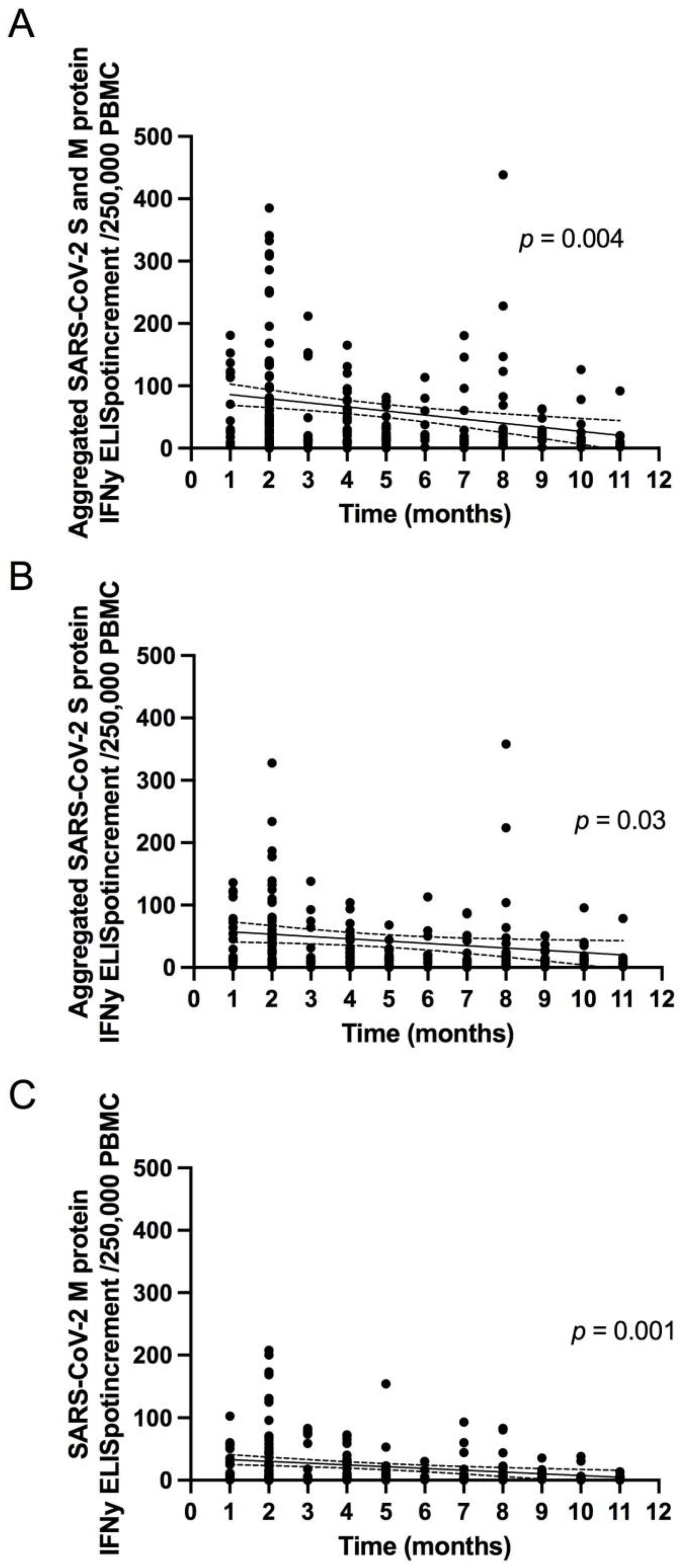
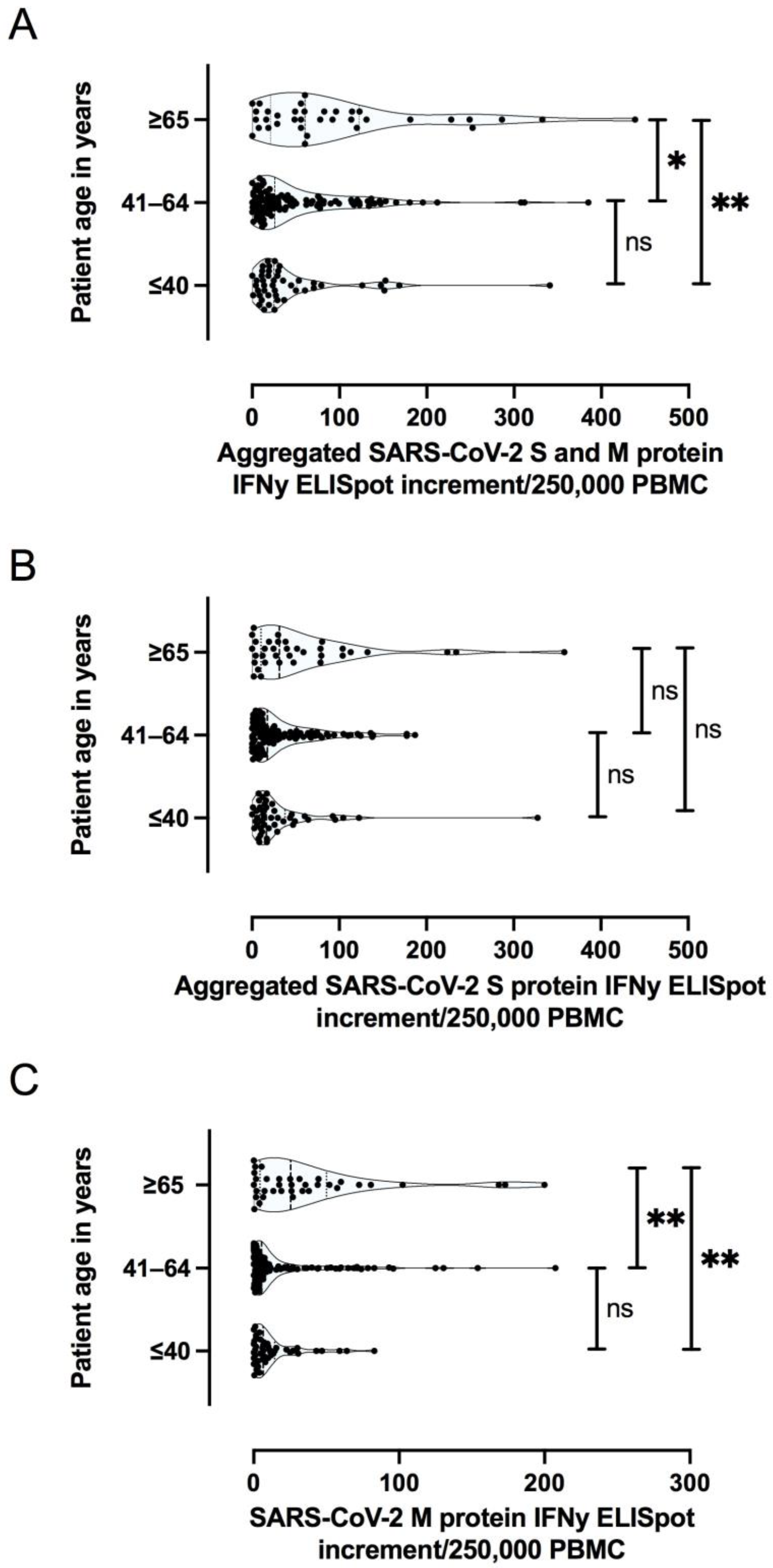
3.2.2. The Magnitude of SARS-CoV-2 Specific T Cell Response Is Dependent on the Severity of Primary COVID-19 and Patient Age
3.3. Specific Antibody Responses to SARS-CoV-2
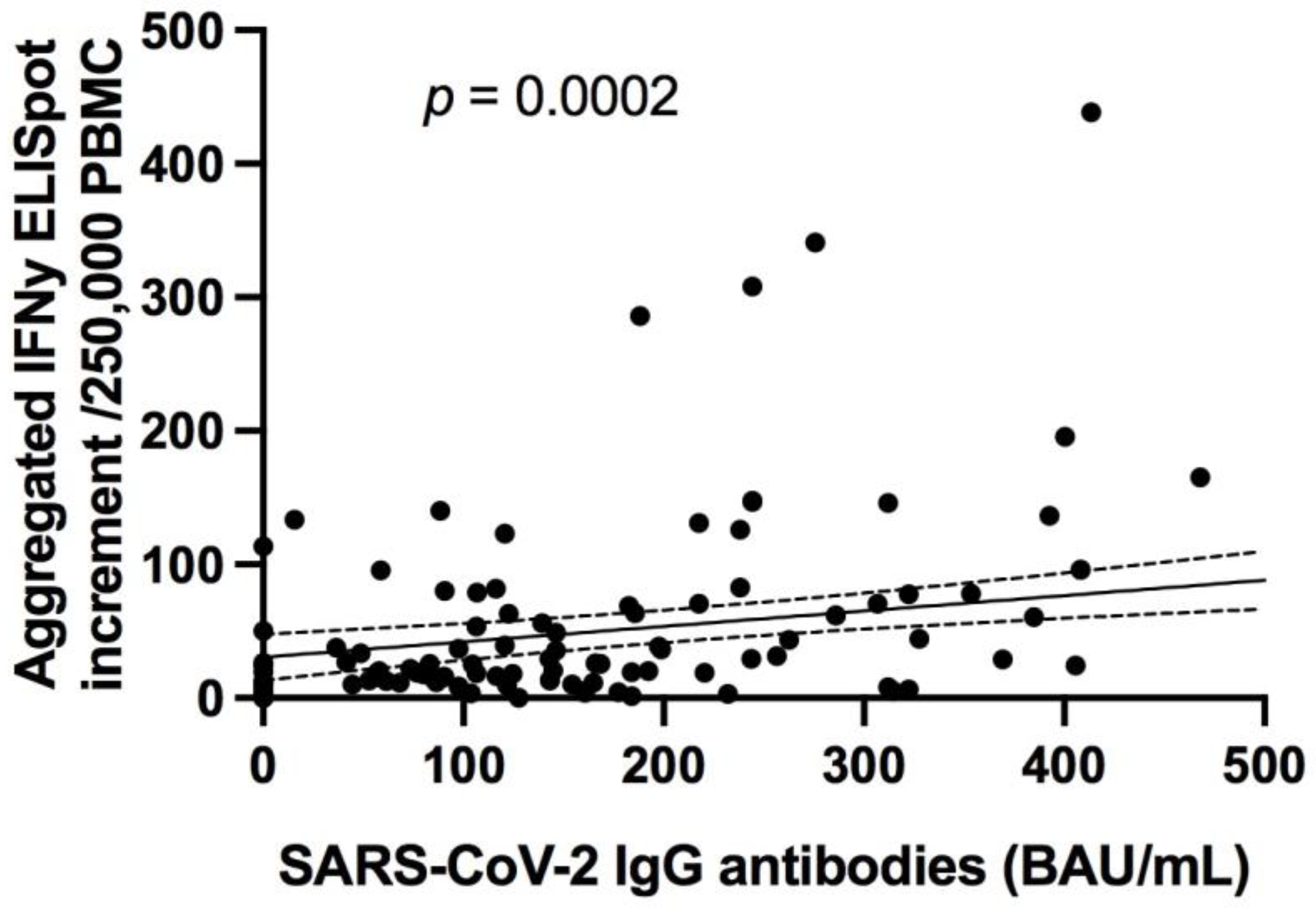
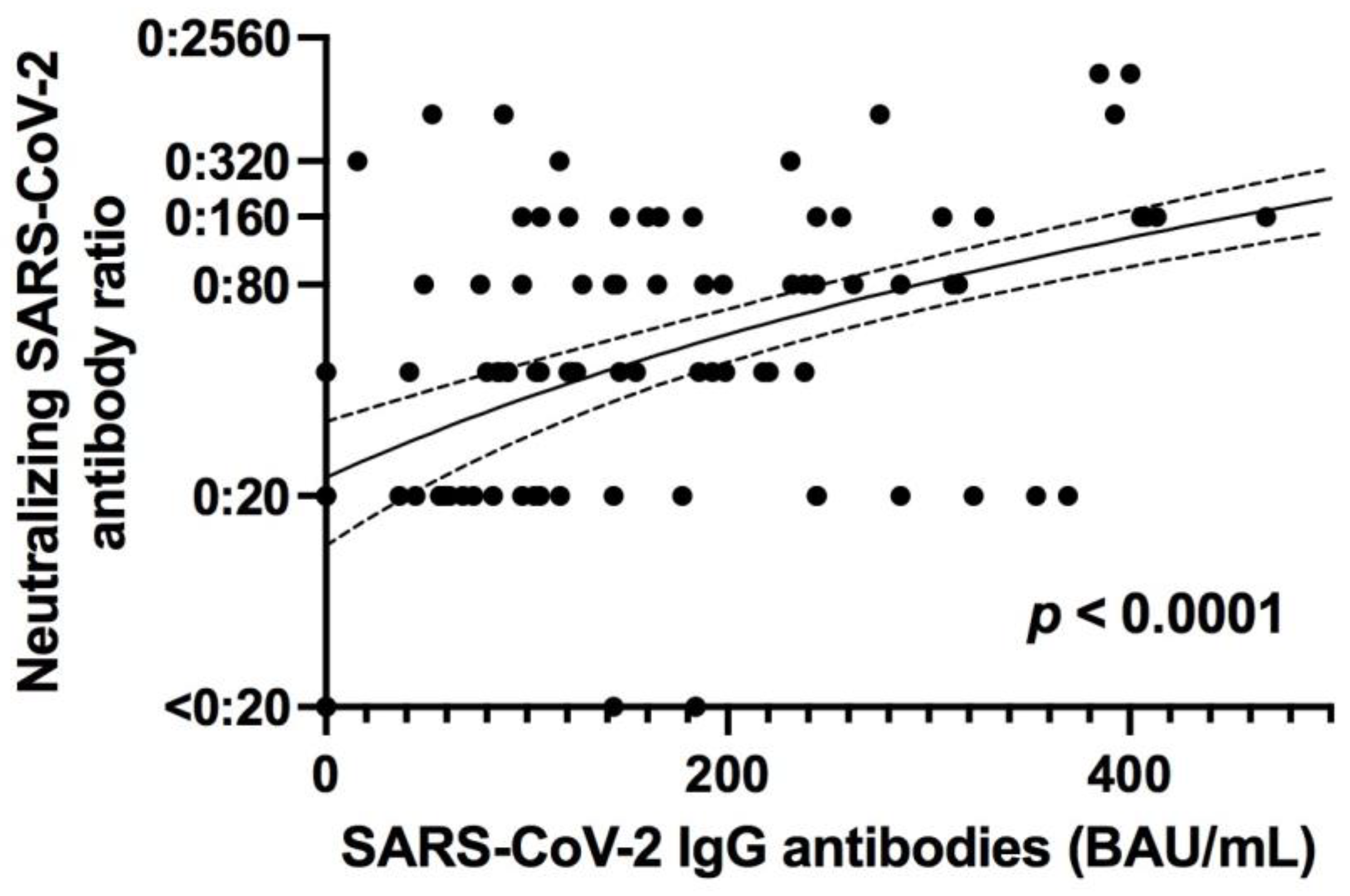
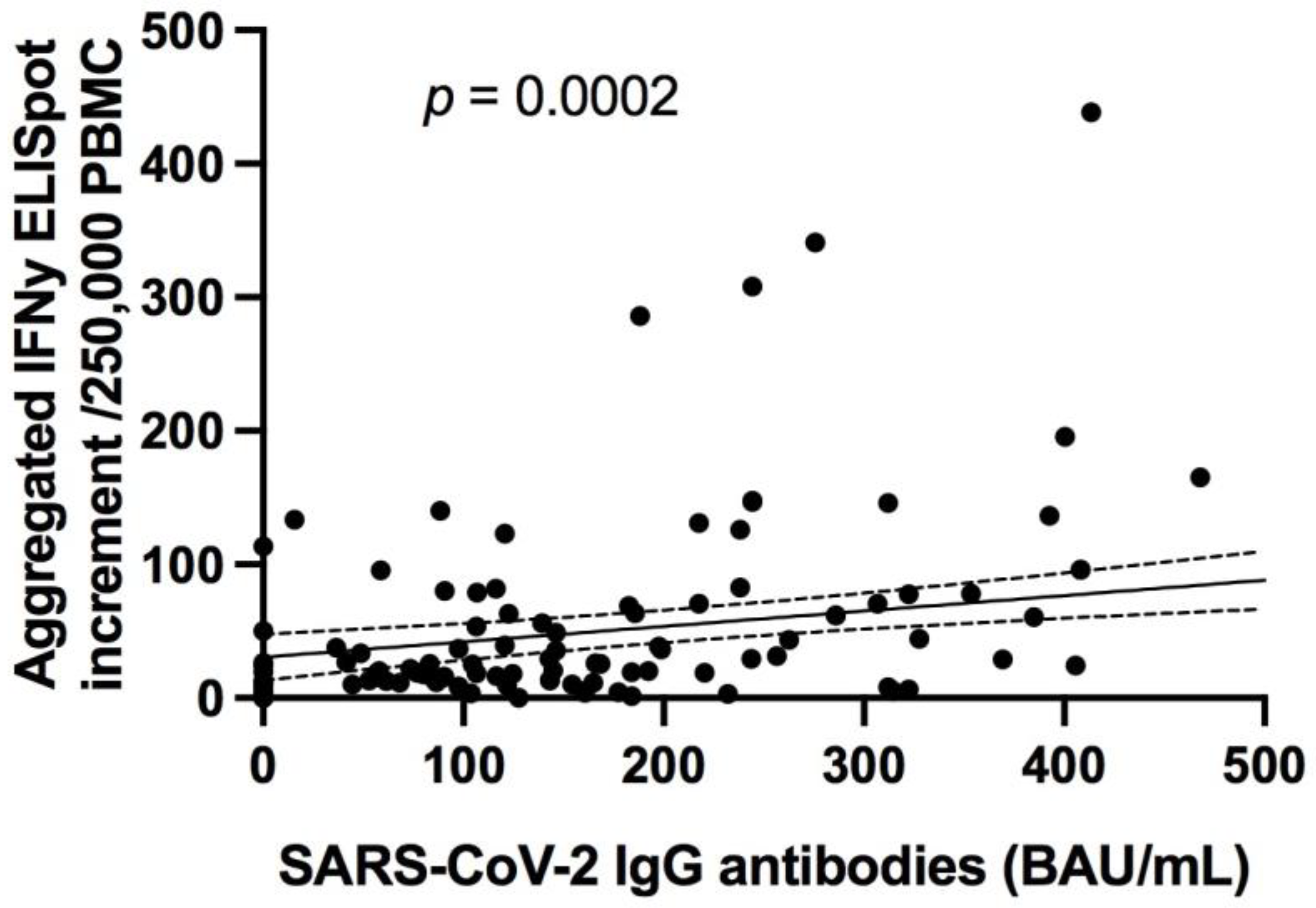
3.3.1. Specific Antibody Responses to SARS-CoV-2 Correlates with Neutralizing Antibodies in Addition to SARS-CoV-2 Specific T Cell Responses
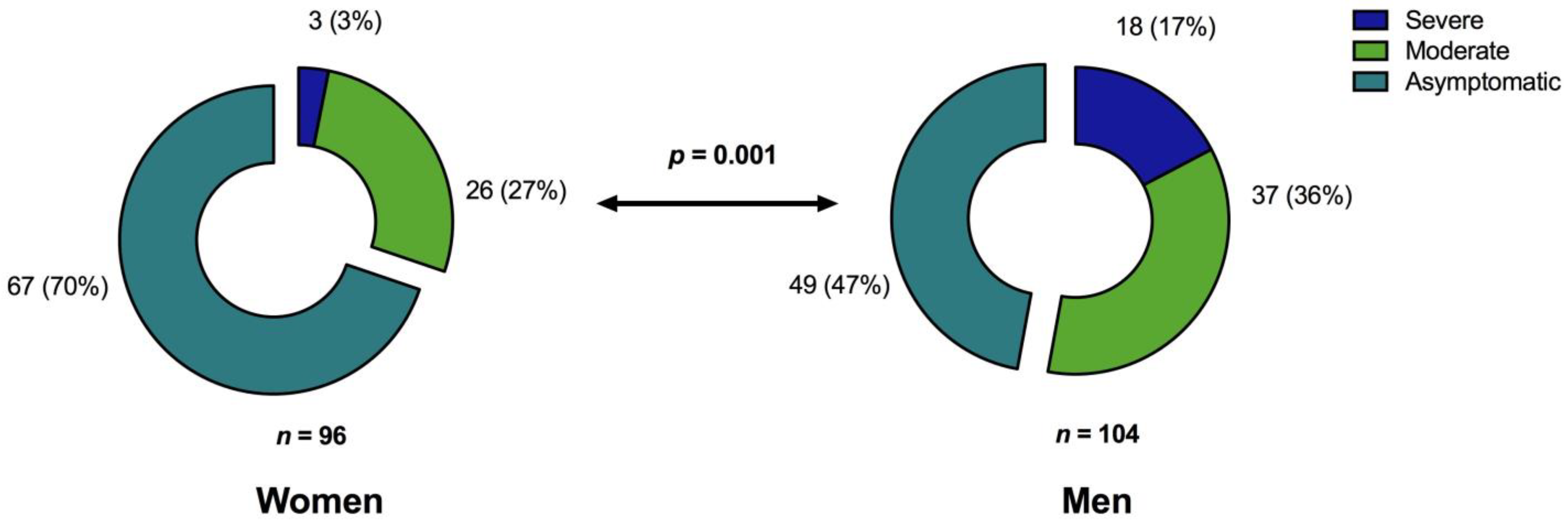
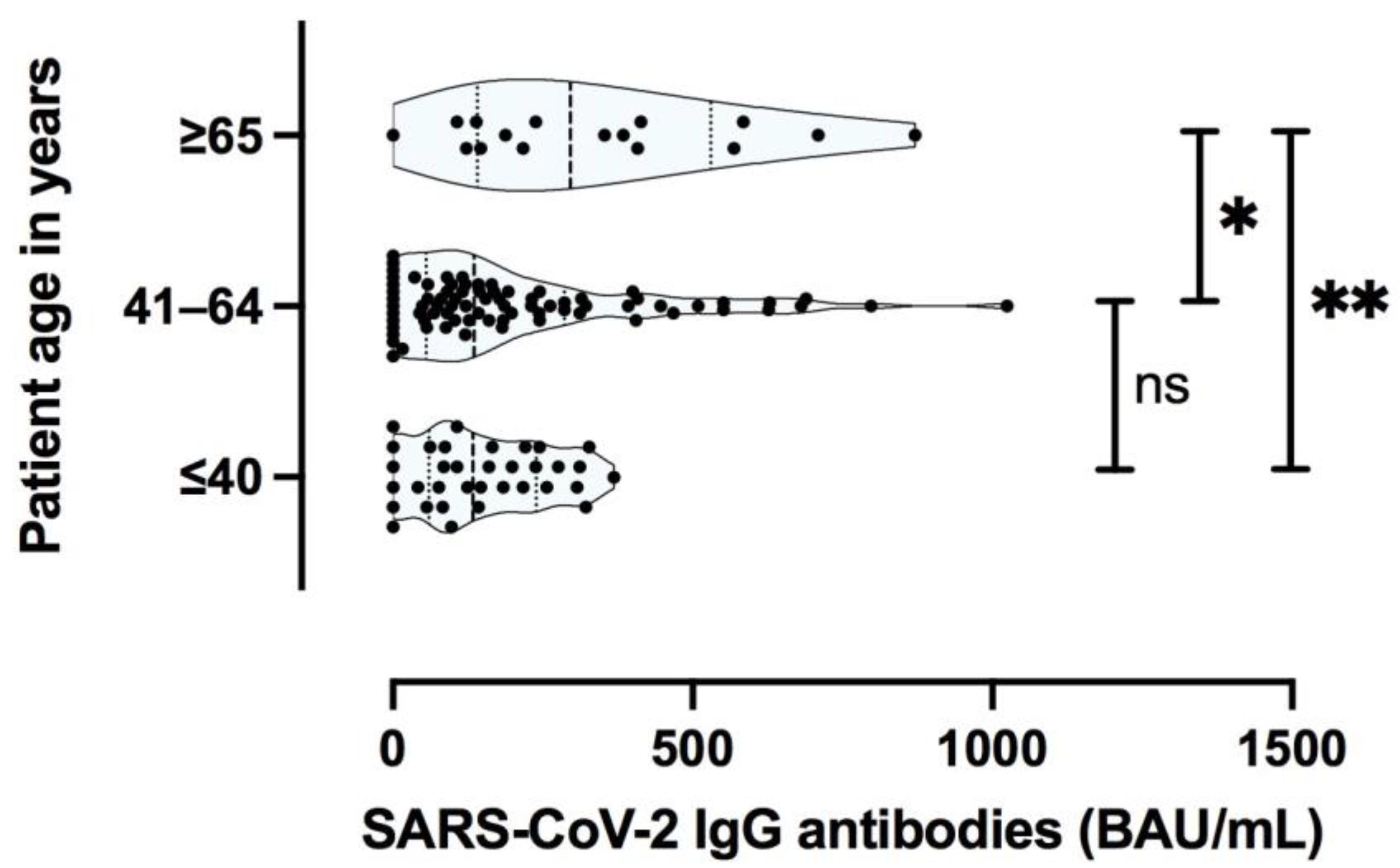
3.3.2. Specific Antibody Responses to SARS-CoV-2 Are Predominantly Influenced by the Severity of the Primary COVID-19
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Liang, B.; Chen, C.; Wang, H.; Fang, Y.; Shen, S.; Yang, X.; Wang, B.; Chen, L.; Chen, Q.; et al. SARS-CoV-2 infection induces sustained humoral immune responses in convalescent patients following symptomatic COVID-19. Nat. Commun. 2021, 12, 1813. [Google Scholar] [CrossRef]
- Wu, J.; Liang, B.Y.; Fang, Y.H.; Wang, H.; Yang, X.L.; Shen, S.; Chen, L.K.; Li, S.M.; Lu, S.H.; Xiang, T.D.; et al. Occurrence of COVID-19 Symptoms during SARS-CoV-2 Infection Defines Waning of Humoral Immunity. Front. Immunol. 2021, 12, 722027. [Google Scholar] [CrossRef]
- The Lancet Respiratory Medicine. Realising the potential of SARS-CoV-2 vaccines-a long shot? Lancet Respir. Med. 2021, 9, 117. [Google Scholar] [CrossRef]
- Kim, D.S.; Rowland-Jones, S.; Gea-Mallorqui, E. Will SARS-CoV-2 Infection Elicit Long-Lasting Protective or Sterilising Immunity? Implications for Vaccine Strategies (2020). Front. Immunol. 2020, 11, 571481. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.A.; Corman, V.M.; Echterhoff, A.K.C.; Muller, M.A.; Richter, A.; Schmandke, A.; Schmidt, M.L.; Schmidt, T.H.; de Vries, F.M.; Drosten, C.; et al. Seroprevalence and correlates of SARS-CoV-2 neutralizing antibodies from a population-based study in Bonn, Germany. Nat. Commun. 2021, 12, 2117. [Google Scholar] [CrossRef]
- Korth, J.; Wilde, B.; Dolff, S.; Anastasiou, O.E.; Krawczyk, A.; Jahn, M.; Cordes, S.; Ross, B.; Esser, S.; Lindemann, M.; et al. SARS-CoV-2-specific antibody detection in healthcare workers in Germany with direct contact to COVID-19 patients. J. Clin. Virol. 2020, 128, 104437. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.; Liang, B.; Fang, Y.; Lu, S.; Li, S.; Wang, H.; Li, H.; Yang, X.; Shen, S.; Zhu, B.; et al. Declining Levels of Neutralizing Antibodies against SARS-CoV-2 in Convalescent COVID-19 Patients One Year Post Symptom Onset. Front. Immunol. 2021, 12, 708523. [Google Scholar] [CrossRef]
- Jakob, C.E.M.; Borgmann, S.; Duygu, F.; Behrends, U.; Hower, M.; Merle, U.; Friedrichs, A.; Tometten, L.; Hanses, F.; Jung, N.; et al. First results of the "Lean European Open Survey on SARS-CoV-2-Infected Patients (LEOSS)". Infection 2021, 49, 63–73. [Google Scholar] [CrossRef]
- Schwarzkopf, S.; Krawczyk, A.; Knop, D.; Klump, H.; Heinold, A.; Heinemann, F.M.; Thummler, L.; Temme, C.; Breyer, M.; Witzke, O.; et al. Cellular Immunity in COVID-19 Convalescents with PCR-Confirmed Infection but with Undetectable SARS-CoV-2-Specific IgG. Emerg. Infect. Dis. 2021, 27, 122–129. [Google Scholar] [CrossRef]
- Lindemann, M.; Krawczyk, A.; Dolff, S.; Konik, M.; Rohn, H.; Platte, M.; Thummler, L.; Schwarzkopf, S.; Schipper, L.; Bormann, M.; et al. SARS-CoV-2-specific humoral and cellular immunity in two renal transplants and two hemodialysis patients treated with convalescent plasma. J. Med. Virol. 2021, 93, 3047–3054. [Google Scholar] [CrossRef]
- Brouwer, P.J.M.; Caniels, T.G.; van der Straten, K.; Snitselaar, J.L.; Aldon, Y.; Bangaru, S.; Torres, J.L.; Okba, N.M.A.; Claireaux, M.; Kerster, G.; et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science 2020, 369, 643–650. [Google Scholar] [CrossRef]
- Heilingloh, C.S.; Aufderhorst, U.W.; Schipper, L.; Dittmer, U.; Witzke, O.; Yang, D.; Zheng, X.; Sutter, K.; Trilling, M.; Alt, M.; et al. Susceptibility of SARS-CoV-2 to UV irradiation. Am. J. Infect. Control 2020, 48, 1273–1275. [Google Scholar] [CrossRef] [PubMed]
- Thieme, C.J.; Anft, M.; Paniskaki, K.; Blazquez-Navarro, A.; Doevelaar, A.; Seibert, F.S.; Hoelzer, B.; Konik, M.J.; Berger, M.M.; Brenner, T.; et al. Robust T Cell Response toward Spike, Membrane, and Nucleocapsid SARS-CoV-2 Proteins Is Not Associated with Recovery in Critical COVID-19 Patients. Cell Rep. Med. 2020, 1, 100092. [Google Scholar] [CrossRef]
- Jin, J.M.; Bai, P.; He, W.; Wu, F.; Liu, X.F.; Han, D.M.; Liu, S.; Yang, J.K. Gender Differences in Patients with COVID-19: Focus on Severity and Mortality. Front. Public Health 2020, 8, 152. [Google Scholar] [CrossRef]
- Ioannou, G.N.; Locke, E.; Green, P.; Berry, K.; O’Hare, A.M.; Shah, J.A.; Crothers, K.; Eastment, M.C.; Dominitz, J.A.; Fan, V.S. Risk Factors for Hospitalization, Mechanical Ventilation, or Death among 10131 US Veterans with SARS-CoV-2 Infection. JAMA Netw. Open 2020, 3, e2022310. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef] [PubMed]
- Westhoff, T.H.; Seibert, F.S.; Bauer, F.; Stervbo, U.; Anft, M.; Doevelaar, A.A.N.; Rohn, B.J.; Winnekendonk, G.; Dittmer, U.; Schenker, P.; et al. Allograft infiltration and meningoencephalitis by SARS-CoV-2 in a pancreas-kidney transplant recipient. Am. J. Transplant. 2020, 20, 3216–3220. [Google Scholar] [CrossRef]
- Ruthrich, M.M.; Giessen-Jung, C.; Borgmann, S.; Classen, A.Y.; Dolff, S.; Gruner, B.; Hanses, F.; Isberner, N.; Kohler, P.; Lanznaster, J.; et al. COVID-19 in cancer patients: Clinical characteristics and outcome-an analysis of the LEOSS registry. Ann. Hematol. 2021, 100, 383–393. [Google Scholar] [CrossRef]
- Candon, S.; Guerrot, D.; Drouot, L.; Lemoine, M.; Lebourg, L.; Hanoy, M.; Boyer, O.; Bertrand, D. T cell and antibody responses to SARS-CoV-2: Experience from a French transplantation and hemodialysis center during the COVID-19 pandemic. Am. J. Transplant. 2021, 21, 854–863. [Google Scholar] [CrossRef]
- Chavarot, N.; Ouedrani, A.; Marion, O.; Leruez-Ville, M.; Villain, E.; Baaziz, M.; Del Bello, A.; Burger, C.; Sberro-Soussan, R.; Martinez, F.; et al. Poor Anti-SARS-CoV-2 Humoral and T-cell Responses after 2 Injections of mRNA Vaccine in Kidney Transplant Recipients Treated with Belatacept. Transplantation 2021, 105, e94–e95. [Google Scholar] [CrossRef] [PubMed]
- Dolff, S.; Korth, J.; Jahn, M.; Kribben, A.; Witzke, O.; Wilde, B. Anti-SARS-CoV-2 T-cell Responses after mRNA Vaccination in Belatacept Treated Renal Transplant Patients. Transplantation 2021, 105, e99. [Google Scholar] [CrossRef]
- Jahn, M.; Korth, J.; Dorsch, O.; Anastasiou, O.E.; Sorge-Hadicke, B.; Tyczynski, B.; Gackler, A.; Witzke, O.; Dittmer, U.; Dolff, S.; et al. Humoral Response to SARS-CoV-2-Vaccination with BNT162b2 (Pfizer-BioNTech) in Patients on Hemodialysis. Vaccines 2021, 9, 360. [Google Scholar] [CrossRef]
- Korth, J.; Jahn, M.; Dorsch, O.; Anastasiou, O.E.; Sorge-Hadicke, B.; Eisenberger, U.; Gackler, A.; Dittmer, U.; Witzke, O.; Wilde, B.; et al. Impaired Humoral Response in Renal Transplant Recipients to SARS-CoV-2 Vaccination with BNT162b2 (Pfizer-BioNTech). Viruses 2021, 13, 756. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Torre, A.; Bueno-Garcia, E.; Lopez-Martinez, R.; Rioseras, B.; Moro-Garcia, M.A.; Alonso-Alvarez, S.; Lluna-Gonzalez, A.; Sousa-Fernandez, A.; Fernandez-Gudin, M.; Campos-Riopedre, L.; et al. Surviving older patients show preserved cellular and humoral immunological memory several months after SARS-CoV-2 infection. J. Gerontol. A Biol. Sci. Med. Sci. 2021. [Google Scholar] [CrossRef]
- Zuo, J.; Dowell, A.C.; Pearce, H.; Verma, K.; Long, H.M.; Begum, J.; Aiano, F.; Amin-Chowdhury, Z.; Hoschler, K.; Brooks, T.; et al. Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following primary infection. Nat. Immunol. 2021, 22, 620–626. [Google Scholar] [CrossRef]
- Sherina, N.; Piralla, A.; Du, L.; Wan, H.; Kumagai-Braesch, M.; Andrell, J.; Braesch-Andersen, S.; Cassaniti, I.; Percivalle, E.; Sarasini, A.; et al. Persistence of SARS-CoV-2-specific B and T cell responses in convalescent COVID-19 patients 6–8 months after the infection. Med 2021, 2, 281–295.e4. [Google Scholar] [CrossRef] [PubMed]
- Gudbjartsson, D.F.; Norddahl, G.L.; Melsted, P.; Gunnarsdottir, K.; Holm, H.; Eythorsson, E.; Arnthorsson, A.O.; Helgason, D.; Bjarnadottir, K.; Ingvarsson, R.F.; et al. Humoral Immune Response to SARS-CoV-2 in Iceland. N. Engl. J. Med. 2020, 383, 1724–1734. [Google Scholar] [CrossRef]
- Wajnberg, A.; Amanat, F.; Firpo, A.; Altman, D.R.; Bailey, M.J.; Mansour, M.; McMahon, M.; Meade, P.; Mendu, D.R.; Muellers, K.; et al. Robust neutralizing antibodies to SARS-CoV-2 infection persist for months. Science 2020, 370, 1227–1230. [Google Scholar] [CrossRef]
- Channappanavar, R.; Zhao, J.; Perlman, S. T cell-mediated immune response to respiratory coronaviruses. Immunol. Res. 2014, 59, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Qiu, Z.; Zhang, L.; Han, Y.; He, W.; Liu, Z.; Ma, X.; Fan, H.; Lu, W.; Xie, J.; et al. Significant changes of peripheral T lymphocyte subsets in patients with severe acute respiratory syndrome. J. Infect. Dis. 2004, 189, 648–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Mentzer, A.J.; Liu, G.; Yao, X.; Yin, Z.; Dong, D.; Dejnirattisai, W.; Rostron, T.; Supasa, P.; Liu, C.; et al. Broad and strong memory CD4(+) and CD8(+) T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19. Nat. Immunol. 2020, 21, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Altmann, D.M. Adaptive immunity to SARS-CoV-2. Oxf. Open Immunol. 2020, 1, iqaa003. [Google Scholar] [CrossRef]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.C.; Wang, Z.; Cho, A.; Agudelo, M.; Barnes, C.O.; Gazumyan, A.; Finkin, S.; et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Sattler, A.; Angermair, S.; Stockmann, H.; Heim, K.M.; Khadzhynov, D.; Treskatsch, S.; Halleck, F.; Kreis, M.E.; Kotsch, K. SARS-CoV-2-specific T cell responses and correlations with COVID-19 patient predisposition. J. Clin. Investig. 2020, 130, 6477–6489. [Google Scholar] [CrossRef]
- Westmeier, J.; Paniskaki, K.; Karakose, Z.; Werner, T.; Sutter, K.; Dolff, S.; Overbeck, M.; Limmer, A.; Liu, J.; Zheng, X.; et al. Impaired Cytotoxic CD8(+) T Cell Response in Elderly COVID-19 Patients. mBio 2020, 11, e02243-20. [Google Scholar] [CrossRef]
- Nikolich-Zugich, J.; Knox, K.S.; Rios, C.T.; Natt, B.; Bhattacharya, D.; Fain, M.J. SARS-CoV-2 and COVID-19 in older adults: What we may expect regarding pathogenesis, immune responses, and outcomes. Geroscience 2020, 42, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.L.; Pekosz, A.; Park, H.S.; Ursin, R.L.; Shapiro, J.R.; Benner, S.E.; Littlefield, K.; Kumar, S.; Naik, H.M.; Betenbaugh, M.J.; et al. Sex, age, and hospitalization drive antibody responses in a COVID-19 convalescent plasma donor population. J. Clin. Investig. 2020, 130, 6141–6150. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Huang, B.; Wu, M.; Zhong, A.; Li, L.; Cai, Y.; Wang, Z.; Wu, L.; Zhu, M.; Li, J.; et al. Dynamic changes in anti-SARS-CoV-2 antibodies during SARS-CoV-2 infection and recovery from COVID-19. Nat. Commun. 2020, 11, 6044. [Google Scholar] [CrossRef]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef]
- Dugas, M.; Grote-Westrick, T.; Vollenberg, R.; Lorentzen, E.; Brix, T.; Schmidt, H.; Tepasse, P.R.; Kuhn, J. Less severe course of COVID-19 is associated with elevated levels of antibodies against seasonal human coronaviruses OC43 and HKU1 (HCoV OC43, HCoV HKU1). Int. J. Infect. Dis. 2021, 105, 304–306. [Google Scholar] [CrossRef]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Arndt, A.L.; Larson, B.J.; Hogue, B.G. A conserved domain in the coronavirus membrane protein tail is important for virus assembly. J. Virol. 2010, 84, 11418–11428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagliani, R.; Forni, D.; Clerici, M.; Sironi, M. Computational Inference of Selection Underlying the Evolution of the Novel Coronavirus, Severe Acute Respiratory Syndrome Coronavirus 2. J. Virol. 2020, 94, e00411-20. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Bard, J.D.; Triche, T.J.; Judkins, A.R.; Biegel, J.A.; Gai, X. Emerging variants of concern in SARS-CoV-2 membrane protein: A highly conserved target with potential pathological and therapeutic implications. Emerg. Microbes Infect. 2021, 10, 885–893. [Google Scholar] [CrossRef]
- Jorrissen, P.; Schutz, P.; Weiand, M.; Vollenberg, R.; Schrempf, I.M.; Ochs, K.; Frommel, C.; Tepasse, P.R.; Schmidt, H.; Zibert, A. Antibody Response to SARS-CoV-2 Membrane Protein in Patients of the Acute and Convalescent Phase of COVID-19. Front. Immunol. 2021, 12, 679841. [Google Scholar] [CrossRef] [PubMed]
- Ayyagari, V.S.; Venkateswarulu, T.C.; Abraham, P.K.; Srirama, K. Design of a multi-epitope-based vaccine targeting M-protein of SARS-CoV2: An immunoinformatics approach. J. Biomol. Struct. Dyn. 2020. [Google Scholar] [CrossRef]
- Bates, T.A.; Weinstein, J.B.; Farley, S.; Leier, H.C.; Messer, W.B.; Tafesse, F.G. Cross-reactivity of SARS-CoV structural protein antibodies against SARS-CoV-2. Cell Rep. 2021, 34, 108737. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Total Group (A) | Asymptomatic Disease (B) | Moderate Disease (C) | Severe Disease (D) | p Value |
|---|---|---|---|---|---|
| No. patients (%) | 200 (100) | 116 (58) | 63 (31.5) | 21 (10.5) | |
| Median age, years (range) | 50 (19–87) | 48 (19–79) | 56 (24–87) | 56 (25–77) | <0.0001 |
| Sex, men/women (%) | 104/96 (52/48) | 49/67 (42.2/57.8) | 37/26 (58.7/41.3) | 18/3 (85.7/14.3) | 0.001 |
| Median BMI (range) | 26.5 (12.1–49.4) | 25.8 (12.1–45.2) | 27.5 (19.5–49.4) | 27.8 (20.5–40.8) | 0.017 |
| Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|
| Variable | Odds Ratio (95% Confidence Interval) | p Value | Odds Ratio (95% Confidence Interval) | p Value |
| Immunosuppressive condition | 7.42 (1.23–44.65) | 0.029 * | 12.53 (1.38–113.95) | 0.025 * |
| Anti-SARS-CoV-2 IgG antibodies positive | 0.056 (0.017–0.186) | <0.0001 * | 0.052 (0.014–0.183) | <0.0001 * |
| Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|
| Variable | Odds Ratio (95% Confidence Interval) | p Value | Odds Ratio (95% Confidence Interval) | p Value |
| Age, years | 1.4 (0.5–2.4) | 0.002 * | 1.1 (0.1–2.0) | 0.031 * |
| Sex (men/women) | 24.2 (−2.5–51.0) | 0.076 | 10.3 (−17.3–37.9) | 0.46 |
| Body mass index (kg/m²) | 1.9 (−0.1–3.9) | 0.056 | 1.1 (−0.9–3.1) | 0.29 |
| Anti-SARS-CoV-2 IgG antibodies positive | 55.2 (10.25–100.1) | 0.016 * | 31.0 (−12.9–74.9) | 0.17 |
| COVID-19 severity | 38.9 (19.4–58.4) | <0.0001 * | 23.85 (0.74–46.96) | 0.043 * |
| Time after SARS-CoV-2 infection (months) | −6.4 (−10.9–−2.08) | 0.004 * | −4.4 (−9.2–0.33) | 0.068 |
| Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|
| Variable | Odds Ratio (95% Confidence Interval) | p Value | Odds Ratio (95% Confidence Interval) | p Value |
| Age, years | 0.8 (0.1–1.5) | 0.03 * | 0.4 (−0.3–1.2) | 0.27 |
| Sex (men/women) | 16.1 (−4.0–36.1) | 0.1 | 4.3 (−16.6–25.1) | 0.7 |
| Body mass index (kg/m²) | 1.4 (−0.1–2.8) | 0.07 | 0.7 (−0.8–2.2) | 0.4 |
| Anti-SARS-CoV-2 IgG antibodies positive | 37.3 (3.6–71.0) | 0.03 * | 22.0 (−11.2–55.3) | 0.19 |
| COVID-19 severity | 31.3 (19.4–58.4) | <0.0001 * | 26.4 (8.9–43.8) | 0.003 * |
| Time after SARS-CoV-2 infection (months) | −3.7 (−6.9–−0.4) | 0.03 * | −1.6 (−5.2–1.9) | 0.4 |
| Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|
| Variable | Odds Ratio (95% Confidence Interval) | p Value | Odds Ratio (95% Confidence Interval) | p Value |
| Age, years | 0.7 (0.3–1.0) | <0.0001 * | 0.7 (0.3–1.0) | <0.0001 * |
| Sex (men/women) | 8.4 (−1.7–18.5) | 0.1 | 6.0 (−4.4–16.4) | 0.26 |
| Body mass index (kg/m²) | 0.6 (−0.2–1.3) | 0.13 | 0.4 (−0.4–1.2) | 0.29 |
| Anti-SARS-CoV-2 IgG antibodies positive | 18.0 (1.0–35.0) | 0.04 * | 9.0 (−7.5–25.6) | 0.28 |
| COVID-19 severity | 7.9 (0.4–15.3) | 0.04 * | −2.5 (−11.2–6.2) | 0.6 |
| Time after SARS-CoV-2 infection (months) | −2.8 (−4.5–1.2) | 0.001 * | −2.8 (−4.5–1.0) | 0.003 * |
| Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|
| Variable | Odds Ratio (95% Confidence Interval) | p Value | Odds Ratio (95% Confidence Interval) | p Value |
| Age, years | 4.1 (1.7–6.5) | 0.001 * | 2.3 (−0.02–4.6) | 0.052 |
| Sex (men/women) | 73.9 (3.8–143.9) | 0.039 * | 20.6 (−42.5–83.7) | 0.52 |
| Body mass index (kg/m²) | 4.9 (−0.2–10.0) | 0.059 | 3.7 (−0.7–8.2) | 0.09 |
| SARS-CoV-2 specific T cells present | 168.9 (45.6–292.2) | 0.008 * | 157.3 (48.7–265.8) | 0.005 * |
| COVID-19 severity | 141.8 (93.6–190.2) | <0.0001 * | 113.8 (63.8–163.8) | <0.0001 * |
| Time after SARS-CoV-2 infection (months) | −5.2 (−16.7–6.4) | 0.3 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konik, M.; Lindemann, M.; Zettler, M.; Meller, L.; Dolff, S.; Rebmann, V.; Horn, P.A.; Dittmer, U.; Krawczyk, A.; Schipper, L.; et al. Long-Term SARS-CoV-2 Specific Immunity Is Affected by the Severity of Initial COVID-19 and Patient Age. J. Clin. Med. 2021, 10, 4606. https://doi.org/10.3390/jcm10194606
Konik M, Lindemann M, Zettler M, Meller L, Dolff S, Rebmann V, Horn PA, Dittmer U, Krawczyk A, Schipper L, et al. Long-Term SARS-CoV-2 Specific Immunity Is Affected by the Severity of Initial COVID-19 and Patient Age. Journal of Clinical Medicine. 2021; 10(19):4606. https://doi.org/10.3390/jcm10194606
Chicago/Turabian StyleKonik, Margarethe, Monika Lindemann, Markus Zettler, Lara Meller, Sebastian Dolff, Vera Rebmann, Peter A. Horn, Ulf Dittmer, Adalbert Krawczyk, Leonie Schipper, and et al. 2021. "Long-Term SARS-CoV-2 Specific Immunity Is Affected by the Severity of Initial COVID-19 and Patient Age" Journal of Clinical Medicine 10, no. 19: 4606. https://doi.org/10.3390/jcm10194606
APA StyleKonik, M., Lindemann, M., Zettler, M., Meller, L., Dolff, S., Rebmann, V., Horn, P. A., Dittmer, U., Krawczyk, A., Schipper, L., Trilling, M., Anastasiou, O. E., Schwarzkopf, S., Thümmler, L., Taube, C., Schöbel, C., Brenner, T., Skoda, E.-M., Wilde, B., ... Rohn, H. (2021). Long-Term SARS-CoV-2 Specific Immunity Is Affected by the Severity of Initial COVID-19 and Patient Age. Journal of Clinical Medicine, 10(19), 4606. https://doi.org/10.3390/jcm10194606