
Association of Genes of the NO Pathway with Altitude Disease and Hypoxic Pulmonary Hypertension

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants and Protocol
2.2. Selection of Single Nucleotide Polymorphisms (SNPs)
2.3. DNA Isolation from Whole Blood
2.4. Genotyping
2.5. Statistical Analyses
3. Results
3.1. Baseline Characteristics
3.2. Allele Frequencies of the Genetic Loci of Interest
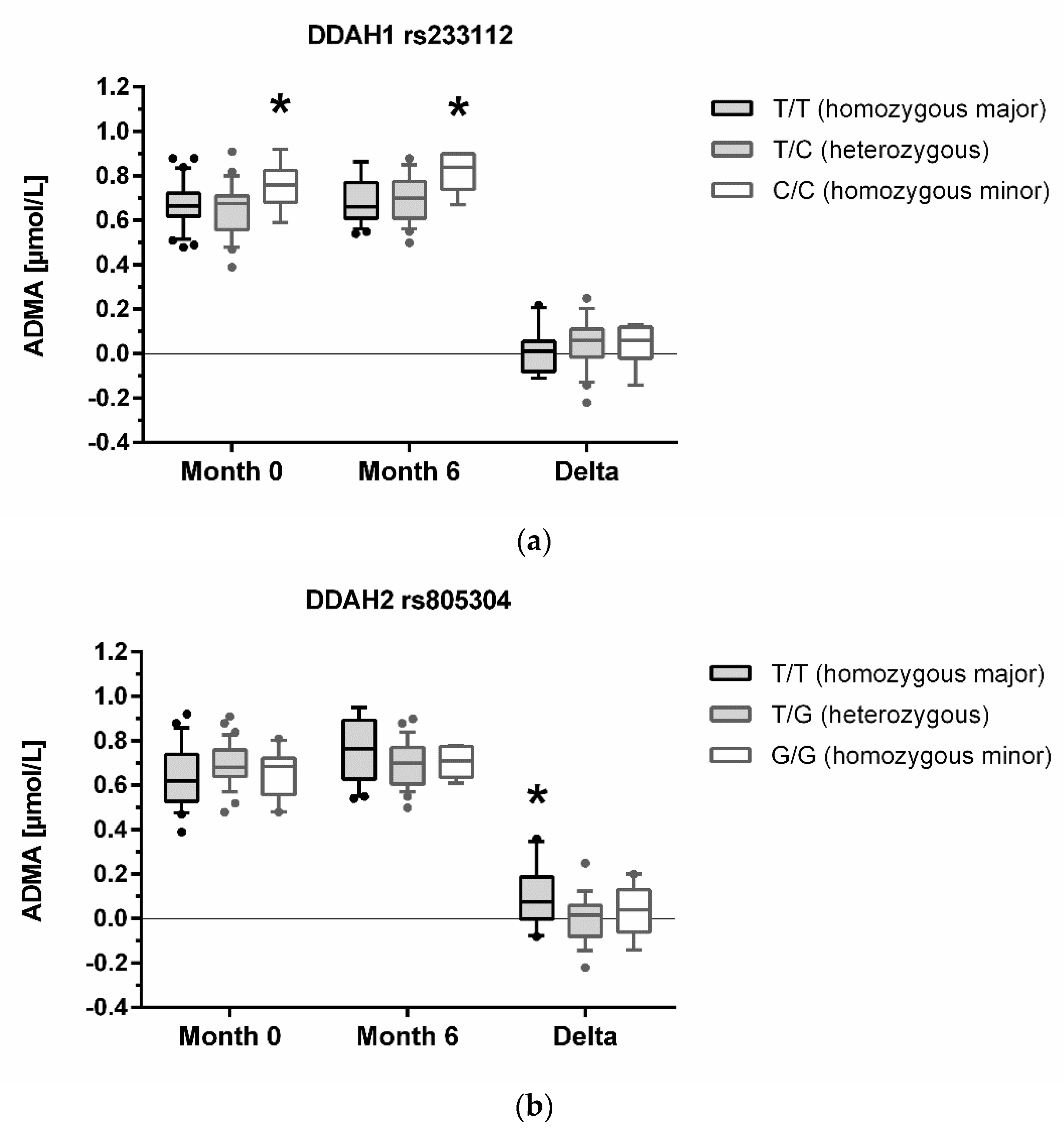
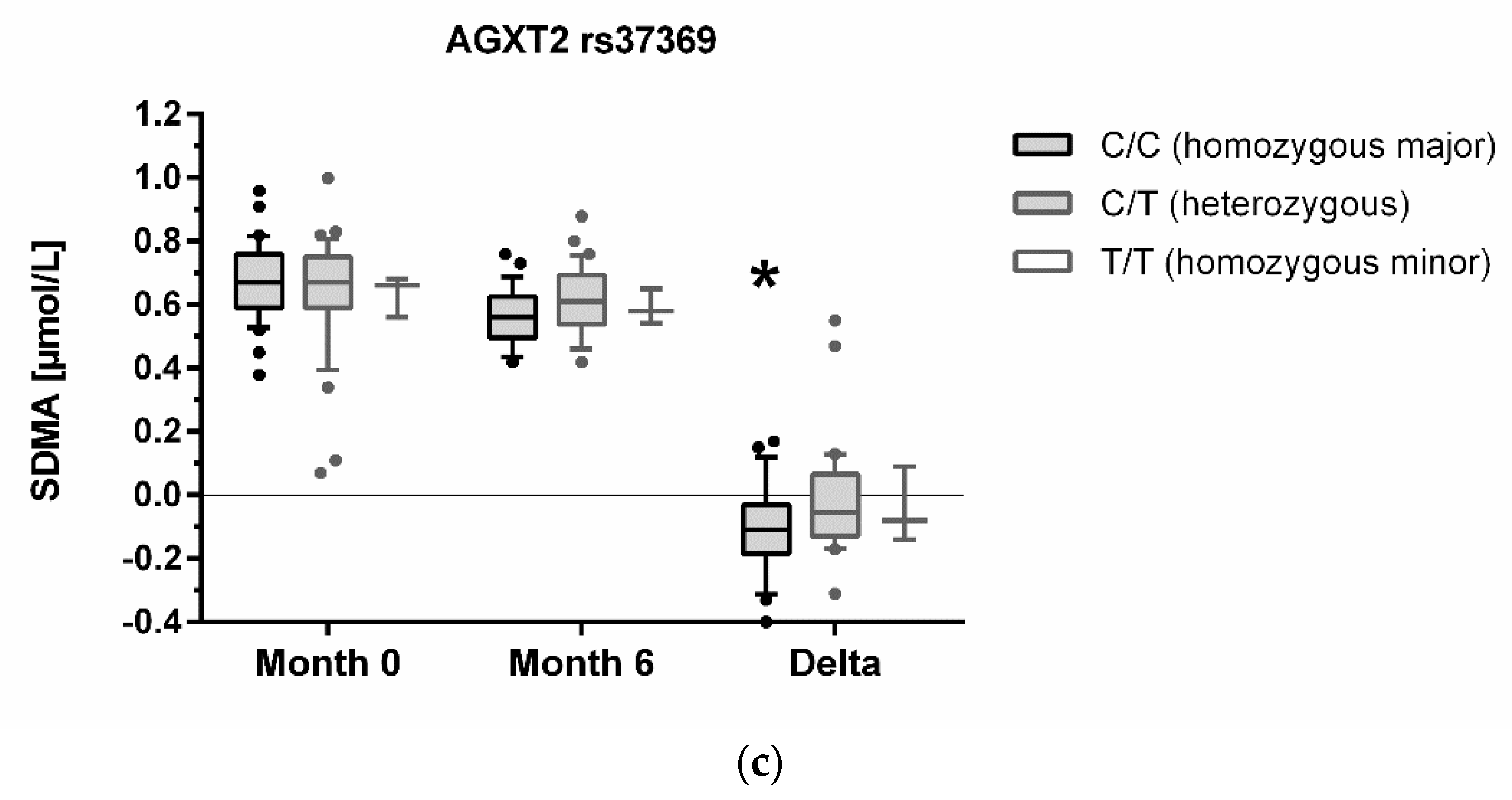
3.3. Association of Genotypes with Biomarker Concentrations
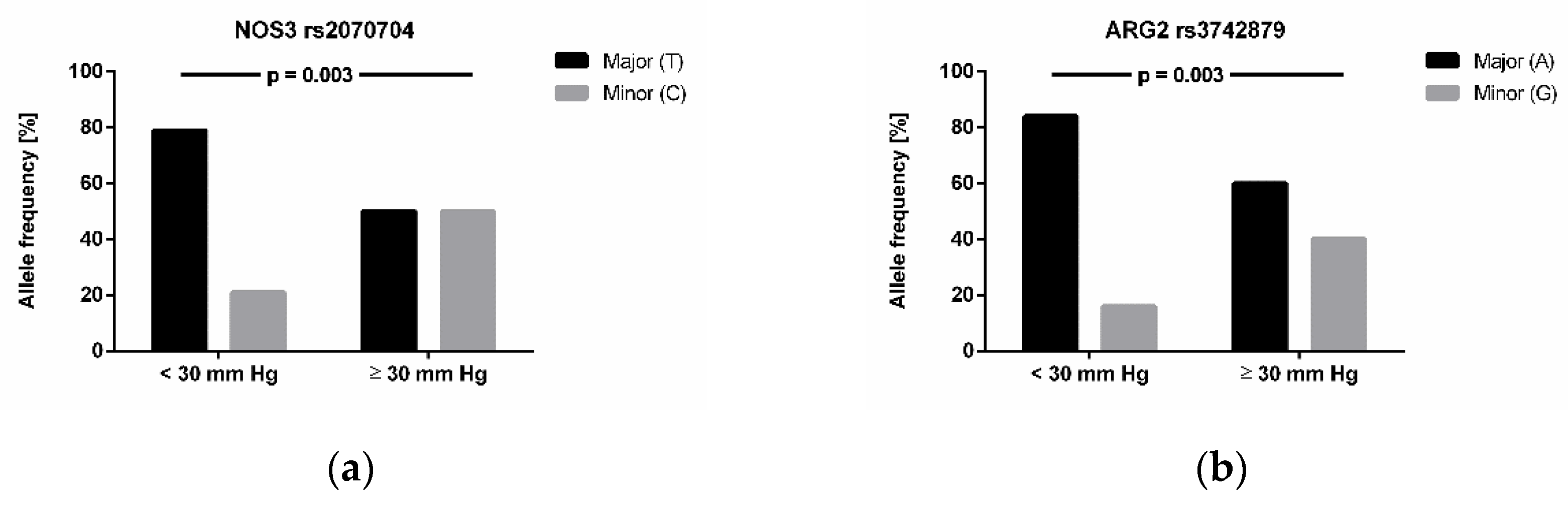
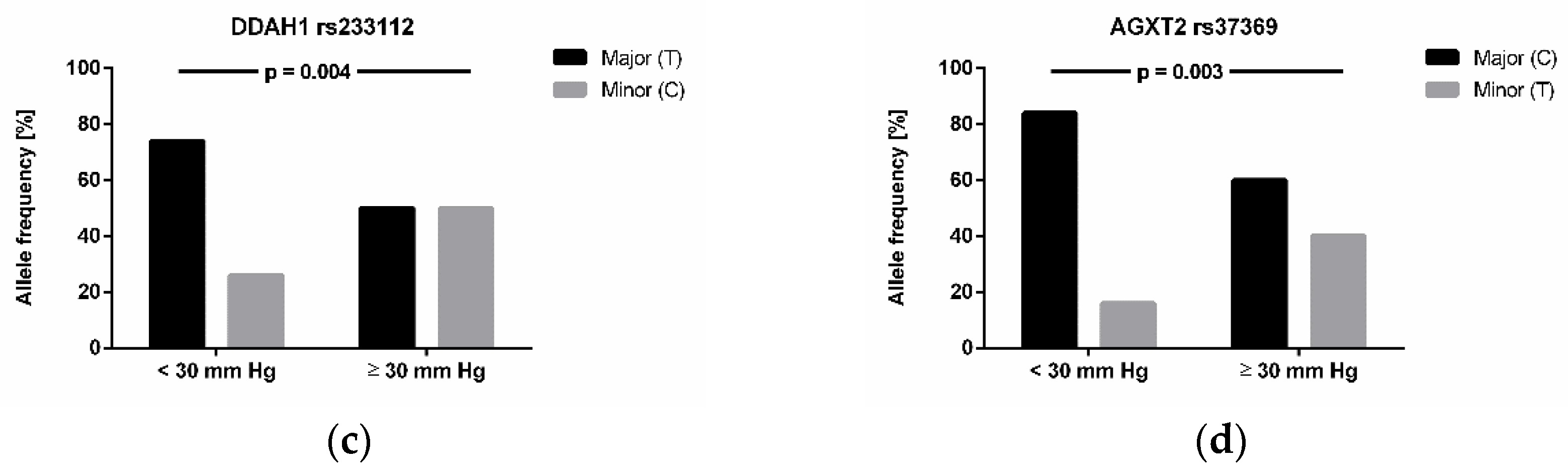
3.4. Association of Genotypes with High-Altitude Pulmonary Hypertension and Altitude Acclimatization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brito, J.; Siques, P.; López, R.; Romero, R.; León-Velarde, F.; Flores, K.; Lüneburg, N.; Hannemann, J.; Böger, R.H. Long-Term Intermittent Work at High Altitude: Right Heart Functional and Morphological Status and Associated Cardiometabolic Factors. Front. Physiol. 2018, 9, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, P.D.; Dantzker, D.R.; Dueck, R.; Clausen, J.L.; West, J.B. Ventilation-perfusion inequality in chronic obstructive pulmonary disease. J. Clin. Investig. 1977, 59, 203–216. [Google Scholar] [CrossRef]
- Calderón-Gerstein, W.S.; López-Peña, A.; Macha-Ramírez, R.; Bruno-Huamán, A.; Espejo-Ramos, R.; Vílchez-Bravo, S.; Ramírez-Breña, M.; Damián-Mucha, M.; Matos-Mucha, A. Endothelial dysfunction assessment by flow-mediated dilation in a high-altitude population. Vasc. Health Risk Manag. 2017, 13, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Hannemann, J.; Zummack, J.; Hillig, J.; Böger, R. Metabolism of asymmetric dimethylarginine in hypoxia: From bench to bedside. Pulm. Circ. 2020, 10, 2045894020918846. [Google Scholar] [CrossRef] [Green Version]
- Lewis, N.C.S.; Bain, A.R.; Wildfong, K.W.; Green, D.J.; Ainslie, P.N. Acute hypoxaemia and vascular function in healthy humans. Exp. Physiol. 2017, 102, 1635–1646. [Google Scholar] [CrossRef] [Green Version]
- Hannemann, J.; Glatzel, A.; Hillig, J.; Zummack, J.; Schumacher, U.; Lüneburg, N.; Harbaum, L.; Böger, R. Upregulation of DDAH2 Limits Pulmonary Hypertension and Right Ventricular Hypertrophy During Chronic Hypoxia in Ddah1 Knockout Mice. Front. Physiol. 2020, 11, 597559. [Google Scholar] [CrossRef]
- Lüneburg, N.; Siques, P.; Brito, J.; Arriaza, K.; Pena, E.; Klose, H.; Leon-Velarde, F.; Böger, R.H. Long-Term Chronic Intermittent Hypobaric Hypoxia in Rats Causes an Imbalance in the Asymmetric Dimethylarginine/Nitric Oxide Pathway and ROS Activity: A Possible Synergistic Mechanism for Altitude Pulmonary Hypertension? Pulm. Med. 2016, 2016, 6578578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siques, P.; Brito, J.; Schwedhelm, E.; Pena, E.; León-Velarde, F.; De La Cruz, J.J.; Böger, R.H.; Hannemann, J. Asymmetric Dimethylarginine at Sea Level Is a Predictive Marker of Hypoxic Pulmonary Arterial Hypertension at High Altitude. Front. Physiol. 2019, 10, 651. [Google Scholar] [CrossRef] [PubMed]
- Fulton, M.D.; Brown, T.; Zheng, Y.G. The Biological Axis of Protein Arginine Methylation and Asymmetric Dimethylarginine. Int. J. Mol. Sci. 2019, 20, 3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüneburg, N.; Lieb, W.; Zeller, T.; Chen, M.H.; Maas, R.; Carter, A.M.; Xanthakis, V.; Glazer, N.L.; Schwedhelm, E.; Seshadri, S.; et al. Genome-wide association study of L-arginine and dimethylarginines reveals novel metabolic pathway for symmetric dimethylarginine. Circ. Cardiovasc. Genet. 2014, 7, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Rodionov, R.N.; Jarzebska, N.; Weiss, N.; Lentz, S.R. AGXT2: A promiscuous aminotransferase. Trends Pharmacol. Sci. 2014, 35, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Lin, Y.; Zhang, R. Associations between endothelial nitric oxide synthase gene polymorphisms and the risk of coronary artery disease: A systematic review and meta-analysis of 132 case-control studies. Eur. J. Prev. Cardiol. 2019, 26, 160–170. [Google Scholar] [CrossRef]
- Anderssohn, M.; McLachlan, S.; Lüneburg, N.; Robertson, C.; Schwedhelm, E.; Williamson, R.M.; Strachan, M.W.; Ajjan, R.; Grant, P.J.; Böger, R.H.; et al. Genetic and environmental determinants of dimethylarginines and association with cardiovascular disease in patients with type 2 diabetes. Diabetes Care 2014, 37, 846–854. [Google Scholar] [CrossRef] [Green Version]
- Maas, R.; Erdmann, J.; Lüneburg, N.; Stritzke, J.; Schwedhelm, E.; Meisinger, C.; Peters, A.; Weil, J.; Schunkert, H.; Böger, R.H.; et al. Polymorphisms in the promoter region of the dimethylarginine dimethylaminohydrolase 2 gene are associated with prevalence of hypertension. Pharmacol. Res. 2009, 60, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Testa, A.; Spoto, B.; Tripepi, G.; Mallamaci, F.; Malatino, L.; Fatuzzo, P.; Maas, R.; Boeger, R.; Zoccali, C. The GLU298ASP variant of nitric oxide synthase interacts with asymmetric dimethyl arginine in determining cardiovascular mortality in patients with end-stage renal disease. J. Hypertens. 2005, 23, 1825–1830. [Google Scholar] [CrossRef]
- Schwedhelm, E.; Maas, R.; Tan-Andresen, J.; Schulze, F.; Riederer, U.; Böger, R.H. High-throughput liquid chromatographic-tandem mass spectrometric determination of arginine and dimethylated arginine derivatives in human and mouse plasma. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 851, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Roach, R.C.; Hackett, P.H.; Oelz, O.; Bärtsch, P.; Luks, A.M.; MacInnis, M.J.; Baillie, J.K. The 2018 Lake Louise Acute Mountain Sickness Score. High Alt. Med. Biol. 2018, 19, 4–6. [Google Scholar] [CrossRef]
- León-Velarde, F.; Maggiorini, M.; Reeves, J.T.; Aldashev, A.; Asmus, I.; Bernardi, L.; Ge, R.L.; Hackett, P.; Kobayashi, T.; Moore, L.G.; et al. Consensus statement on chronic and subacute high altitude diseases. High Alt. Med. Biol. 2005, 6, 147–157. [Google Scholar] [CrossRef]
- Machiela, M.J.; Chanock, S.J. LDlink: A web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef] [PubMed]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Statist. 1979, 6, 65–70. [Google Scholar]
- Machiela, M.J.; Chanock, S.J. LDassoc: An online tool for interactively exploring genome-wide association study results and prioritizing variants for functional investigation. Bioinformatics 2018, 34, 887–889. [Google Scholar] [CrossRef] [PubMed]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, L.; Fairley, S.; Zheng-Bradley, X.; Streeter, I.; Perry, E.; Lowy, E.; Tasse, A.M.; Flicek, P. The international Genome sample resource (IGSR): A worldwide collection of genome variation incorporating the 1000 Genomes Project data. Nucleic Acids Res. 2017, 45, D854–D859. [Google Scholar] [CrossRef] [Green Version]
- Telo, S.; Kırkıl, G.; Kuluöztürk, M.; Balin, M.; Deveci, F. Can ADMA play a role in determining pulmonary hypertension related to chronic obstructive pulmonary disease? Clin. Respir. J. 2018, 12, 1433–1438. [Google Scholar] [CrossRef]
- Stautemas, J.; Van Kuilenburg, A.B.P.; Stroomer, L.; Vaz, F.; Blancquaert, L.; Lefevere, F.B.D.; Everaert, I.; Derave, W. Acute Aerobic Exercise Leads to Increased Plasma Levels of R- and S-β-Aminoisobutyric Acid in Humans. Front. Physiol. 2019, 10, 1240. [Google Scholar] [CrossRef] [Green Version]
- Suhre, K.; Wallaschofski, H.; Raffler, J.; Friedrich, N.; Haring, R.; Michael, K.; Wasner, C.; Krebs, A.; Kronenberg, F.; Chang, D.; et al. A genome-wide association study of metabolic traits in human urine. Nat. Genet. 2011, 43, 565–569. [Google Scholar] [CrossRef]
- Leiper, J.M.; Santa Maria, J.; Chubb, A.; MacAllister, R.J.; Charles, I.G.; Whitley, G.S.; Vallance, P. Identification of two human dimethylarginine dimethylaminohydrolases with distinct tissue distributions and homology with microbial arginine deiminases. Biochem. J. 1999, 343 Pt 1, 209–214. [Google Scholar] [CrossRef]
- Wang, D.; Gill, P.S.; Chabrashvili, T.; Onozato, M.L.; Raggio, J.; Mendonca, M.; Dennehy, K.; Li, M.; Modlinger, P.; Leiper, J.; et al. Isoform-specific regulation by N(G),N(G)-dimethylarginine dimethylaminohydrolase of rat serum asymmetric dimethylarginine and vascular endothelium-derived relaxing factor/NO. Circ. Res. 2007, 101, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Lambden, S.; Kelly, P.; Ahmetaj-Shala, B.; Wang, Z.; Lee, B.; Nandi, M.; Torondel, B.; Delahaye, M.; Dowsett, L.; Piper, S.; et al. Dimethylarginine dimethylaminohydrolase 2 regulates nitric oxide synthesis and hemodynamics and determines outcome in polymicrobial sepsis. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1382–1392. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, S.; Ishizaki, T.; Toga, H.; Sakai, A.; Isakova, J.; Taalaibekova, E.; Baiserkeev, Z.; Kojonazarov, B.; Aldashev, A. Endogenous Asymmetric Dimethylarginine Pathway in High Altitude Adapted Yaks. BioMed Res. Int. 2015, 2015, 196904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, V.; Moraga, F.A.; Llanos, A.J.; Ebensperger, G.; Taborda, M.I.; Uribe, E. Plasmatic Concentrations of ADMA and Homocystein in Llama (Lama glama) and Regulation of Arginase Type II: An Animal Resistent to the Development of Pulmonary Hypertension Induced by Hypoxia. Front. Physiol. 2018, 9, 606. [Google Scholar] [CrossRef] [PubMed]
- Simonneau, G.; Montani, D.; Celermajer, D.S.; Denton, C.P.; Gatzoulis, M.A.; Krowka, M.; Williams, P.G.; Souza, R. Haemodynamic definitions and updated clinical classification of pulmonary hypertension. Eur. Respir. J. 2019, 53, 1801913. [Google Scholar] [CrossRef] [PubMed]
- Naeije, R. Pulmonary hypertension at high altitude. Eur. Respir. J. 2019, 53, 1900985. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Units | Baseline | 6 Months |
|---|---|---|---|
| Age | years | 18.0 (18.0–19.0) | n.a. |
| Male sex | n (%) | 69 (100) | n.a. |
| Smoker | n (%) | 35 (50.7) | n.a. |
| Height | m | 1.73 (1.68–1.76) | n.a. |
| Weight | kg | 69.0 (63.0–78.0) | n.a. |
| BMI | kg/m2 | 24.2 (21.6–26.1) | n.a. |
| Arterial oxygen saturation | % | 98.0 (98.0–98.5) | 91.0 (87.5–93.0) ** |
| Systolic blood pressure | mm Hg | 110 (105–120) | 110 (100–112.5) |
| Diastolic blood pressure | mm Hg | 70.0 (70.0–75.0) | 70.0 (65.0–80.0) |
| Heart rate | L/min | 71.5 (65.5–78.3) | 74.5 (68.5–82.0) |
| Haematocrit | % | 45.0 (44.0–46.0) | 48.5 (46.6.–49.9) ** |
| Haemoglobin | mg/dL | 14.9 (14.5–15.4) | 16.0 (15.7–16.4) ** |
| ADMA | µmol/L | 0.68 (0.60–0.75) | 0.72 (0.62–0.78) * |
| SDMA | µmol/L | 0.67 (0.59–0.75) | 0.58 (0.53–0.66) * |
| L-Arginine | µmol/L | 16.6 (11.9–23.4) | 29.6 (21.1–34.6) ** |
| Gene/SNP | Major/Minor Allele | Expected Allele Frequency # | Measured Allele Frequency | p |
|---|---|---|---|---|
| NOS3 | ||||
| rs1799983 | G/T | 0.785/0.215 | 0.790/0.210 | ns |
| rs2070744 | T/C | 0.742/0.258 | 0.717/0.283 | ns |
| rs891512 | G/A | 0.860/0.140 | 0.862/0.138 | ns |
| DDAH1 | ||||
| rs1241321 | A/G | 0.671/0.239 | 0.674/0.326 | ns |
| rs480414 | G/A | 0.735/0.265 | 0.812/0.188 | ns |
| rs233112 | T/C | 0.666/0.334 | 0.652/0.348 | ns |
| DDAH2 | ||||
| rs805304 | T/G | 0.549/0.451 | 0.601/0.399 | ns |
| rs2272592 | C/T | 0.925/0.075 | 0.913/0.087 | ns |
| ARG1 | ||||
| rs2246012 | T/C | 0.751/0.249 | 0.775/0.225 | ns |
| rs2781667 | C/T | 0.524/0.475 | 0.580/0.420 | ns |
| ARG2 | ||||
| rs3742879 | A/G | 0.767/0.233 | 0.775/0.225 | ns |
| rs3759757 | G/C | 0.635/0.365 | 0.659/0.341 | ns |
| AGXT2 | ||||
| rs37369 | C/T | 0.676/0.324 | 0.703/0.297 | ns |
| rs16899974 | C/A | 0.739/0.261 | 0.710/0.290 | ns |
| PRMT1 | ||||
| rs10415880 | G/A | 0.801/0.199 | 0.833/0.167 | ns |
| rs975484 | C/G | 0.682/0.318 | 0.768/0.232 | 0.0434 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hannemann, J.; Siques, P.; Schmidt-Hutten, L.; Zummack, J.; Brito, J.; Böger, R. Association of Genes of the NO Pathway with Altitude Disease and Hypoxic Pulmonary Hypertension. J. Clin. Med. 2021, 10, 5761. https://doi.org/10.3390/jcm10245761
Hannemann J, Siques P, Schmidt-Hutten L, Zummack J, Brito J, Böger R. Association of Genes of the NO Pathway with Altitude Disease and Hypoxic Pulmonary Hypertension. Journal of Clinical Medicine. 2021; 10(24):5761. https://doi.org/10.3390/jcm10245761
Chicago/Turabian StyleHannemann, Juliane, Patricia Siques, Lena Schmidt-Hutten, Julia Zummack, Julio Brito, and Rainer Böger. 2021. "Association of Genes of the NO Pathway with Altitude Disease and Hypoxic Pulmonary Hypertension" Journal of Clinical Medicine 10, no. 24: 5761. https://doi.org/10.3390/jcm10245761
APA StyleHannemann, J., Siques, P., Schmidt-Hutten, L., Zummack, J., Brito, J., & Böger, R. (2021). Association of Genes of the NO Pathway with Altitude Disease and Hypoxic Pulmonary Hypertension. Journal of Clinical Medicine, 10(24), 5761. https://doi.org/10.3390/jcm10245761