
Evaluation of Radial Peripapillary Capillary Density in G6PD Deficiency: An OCT Angiography Pilot Study

 , ,
, ,
Abstract
:1. Introduction
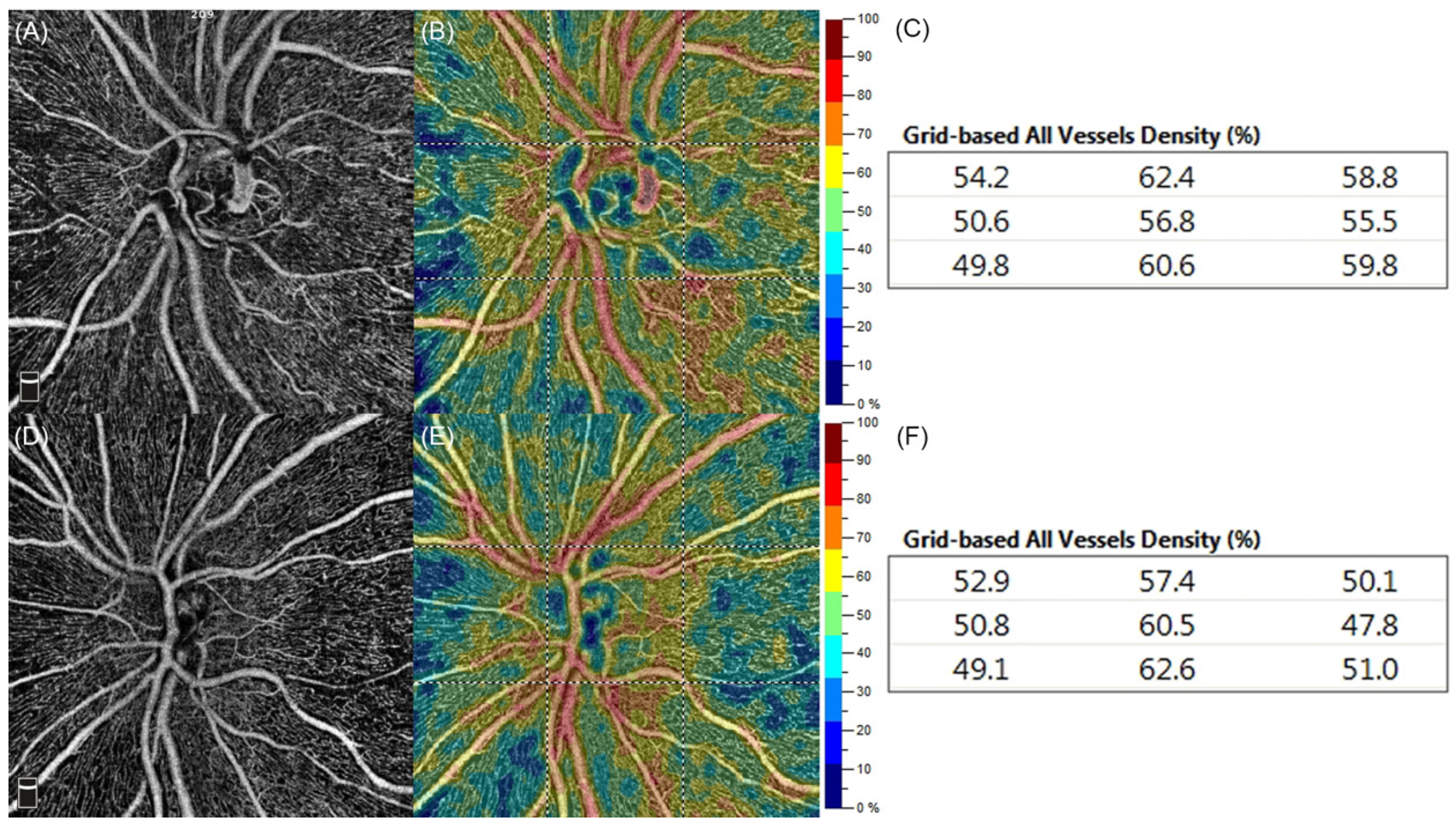
2. Materials and Methods
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luzzatto, L.; Poggi, V. Glucose-6-phosphate dehydrogenase deficiency. In Nathan and Oski’s Hematology of Infancy and Childhood, 7th ed.; Orkin, S.H., Nathan, D.G., Ginsburg, D., Look, A.T., Fisher, D.E., Lux, S.E., Eds.; Saunders Elsevier: Philadelphia, PA, USA, 2009; Volume 1, pp. 883–907. [Google Scholar]
- Cocco, P.; Todde, P.; Fornera, S.; Manca, M.B.; Manca, P.; Sias, A.R. Mortality in a cohort of men expressing the glucose-6-phosphate dehydrogenase deficiency. Blood 1998, 91, 706–709. [Google Scholar] [CrossRef] [Green Version]
- Beutler, E. G6PD deficiency. Blood 1994, 84, 3613. [Google Scholar] [CrossRef] [Green Version]
- Pinna, A.; Carru, C.; Solinas, G.; Zinellu, A.; Carta, F. Glucose-6-Phosphate Dehydrogenase Deficiency in Retinal Vein Occlusion. Investig. Opthalmol. Vis. Sci. 2007, 48, 2747–2752. [Google Scholar] [CrossRef] [Green Version]
- Siniscalco, M.; Bernini, L.; Latte, B.; Motulsky, A.G. Favism and Thalassæmia and their Relationship to Malaria. Nature 1961, 190, 1179–1180. [Google Scholar] [CrossRef]
- Dore, M.P.; Davoli, A.; Longo, N.; Marras, G.; Pes, G.M. Glucose-6-phosphate dehydrogenase deficiency and risk of colorectal cancer in Northern Sardinia: A retrospective observational study. Medicine 2016, 95, e5254. [Google Scholar] [CrossRef]
- Pinna, A.; Solinas, G.; Masia, C.; Zinellu, A.; Carru, C.; Carta, A. Glucose-6-Phosphate Dehydrogenase (G6PD) Deficiency in Nonarteritic Anterior Ischemic Optic Neuropathy in a Sardinian Population, Italy. Investig. Opthalmol. Vis. Sci. 2008, 49, 1328–1332. [Google Scholar] [CrossRef]
- Pinna, A.; Contini, E.L.; Carru, C.; Solinas, G. Glucose-6-Phosphate Dehydrogenase Deficiency and Diabetes Mellitus with Severe Retinal Complications in a Sardinian Population, Italy. Int. J. Med. Sci. 2013, 10, 1907–1913. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.M.; Geng, L.; Cahill-Smith, S.; Liu, F.; Douglas, G.; Mckenzie, C.A.; Smith, C.; Brooks, G.; Channon, K.M.; Li, J.M. Nox2 contributes to age-related oxidative damage to neurons and the cerebral vasculature. J Clin. Investig. 2022, 51, 3374–3386. [Google Scholar] [CrossRef]
- Serra, R.; Coscas, F.; Boulet, J.F.; Cabral, D.R.; Lupidi, M.; Coscas, G.J.; Souied, E.H. Predictive Activation Biomarkers of Treatment-Naive Asymptomatic Choroidal Neovascularization in Age-Related Macular Degeneration. Retina 2020, 40, 1224–1233. [Google Scholar] [CrossRef]
- Spaide, R.F.; Klancnik, J.M.; Cooney, M.J. Retinal Vascular Layers Imaged by Fluorescein Angiography and Optical Coherence Tomography Angiography. JAMA Ophthalmol. 2015, 133, 45–50. [Google Scholar] [CrossRef]
- Serra, R.; Sellam, A.; Coscas, F.; Bruyère, E.; Sieiro, A.; Coscas, G.J.; Souied, E.H. Evaluation of pseudophakic cystoid macular edema using optical coherence tomography angiography. Eur. J. Ophthalmol. 2018, 28, 234–240. [Google Scholar] [CrossRef]
- Scoles, D.; Gray, D.C.; Hunter, J.J.; Wolfe, R.; Gee, B.P.; Geng, Y.; Masella, B.D.; Libby, R.T.; Russell, S.; Williams, D.R.; et al. In-Vivo imaging of retinal nerve fiber layer vasculature: Imaging histology comparison. BMC Ophthalmol. 2009, 23, 9. [Google Scholar] [CrossRef] [Green Version]
- Alterman, M.; Henkind, P. Radial peripapillary capillaries of the retina. II. Possible role in Bjerrum scotoma. Br. J. Ophthalmol. 1968, 52, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Kornzweig, A.L.; Eliasoph, I.; Feldstein, M. Selective Atrophy of the Radial Peripapillary Capillaries in Chronic Glaucoma. Arch. Ophthalmol. 1968, 80, 696–702. [Google Scholar] [CrossRef]
- Mansoori, T.; Sivaswamy, J.; Gamalapati, J.S.; Balakrishna, N. Radial Peripapillary Capillary Density Measurement Using Optical Coherence Tomography Angiography in Early Glaucoma. J. Glaucoma 2017, 26, 438–443. [Google Scholar] [CrossRef]
- Michaelson, I. Retinal Circulation in Man and Animals; Charles C Thomas: Springfield, IL, USA, 1954. [Google Scholar]
- Henkind, P. Radial peripapillary capillaries of the retina. I. Anatomy: Human and comparative. Br. J. Ophthalmol. 1967, 51, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.K.; Balaratnasingam, C.; Xu, J.; Morgan, W.H.; Mammo, Z.; Han, S.; MacKenzie, P.; Merkur, A.; Kirker, A.; Albiani, D.; et al. Label-Free Density Measurements of Radial Peripapillary Capillaries in the Human Retina. PLoS ONE 2015, 10, e0135151. [Google Scholar] [CrossRef]
- Ip, M.; Hendrick, A. Retinal Vein Occlusion Review. Asia Pac. J. Ophthalmol. (Phila) 2018, 7, 40–45. [Google Scholar] [CrossRef]
- Clarkson, J.G. Central retinal vein occlusion. In Medical Retina, 3rd ed.; Ryan, S.J., Ed.; Mosby Inc.: St. Louis, MO, USA, 2001; Volume 2, pp. 1368–1375. [Google Scholar]
- Fekrat, S.; Finkelstein, D. Branch retinal vein occlusion. In Medical Retina, 3rd ed.; Ryan, S.J., Ed.; Mosby Inc.: St. Louis, MO, USA, 2001; Volume 2, pp. 1376–1381. [Google Scholar]
- Batetta, B.; Bonatesta, R.R.; Sanna, F.; Putzolu, M.; Mulas, M.F.; Collu, M.; Dessì, S. Cell growth and cholesterol metabolism in human glucose- 6-phosphate dehydrogenase deficient lymphomononuclear cells. Cell Prolif. 2002, 35, 143–154. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Radhakrishnan, R.; Mohammad, G. Regulation of Rac1 transcription by histone and DNA methylation in diabetic retinopathy. Sci. Rep. 2021, 11, 14097. [Google Scholar] [CrossRef]
- Wallace, M. NADPH diaphorase activity in activated astrocytes representing inducible nitric oxide synthase. Methods Enzym. 1996, 268, 497–503. [Google Scholar] [CrossRef]
- Li, H.; Poulos, T.L. Structure–function studies on nitric oxide synthases. J. Inorg. Biochem. 2005, 99, 293–305. [Google Scholar] [CrossRef]
- Stanton, R.C. Glucose-6-phosphate dehydrogenase, NADPH, and cell survival. IUBMB Life 2012, 64, 362–369. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| G6PD-Deficient Subjects | G6PD-Normal Controls | |
|---|---|---|
| Total eyes, n | 46 | 23 |
| Total patients, n | 46 | 23 |
| Age, mean ± SD (years) | 43.93 ± 15.79 | 45.30 ± 18.02 |
| BCVA, mean ± SD (LogMAR) | 0.07 ± 0.12 | −0.10 ± 0.10 |
| Refractive error, mean ± SD (spherical equivalent) | −0.69 ± 2.01 | −0.55 ± 1.05 |
| Peripapillary SECTORS | G6PD-Deficient Subjects | G6PD-Normal Controls | p Value |
|---|---|---|---|
| Supero-temporal | 57.71 ± 4.27 | 56.49 ± 4.29 | >0.05 |
| Supero-middle | 58.43 ± 4.08 | 58.08 ± 3.35 | >0.05 |
| Supero-nasal | 50.02 ± 4.68 | 49.33 ± 5.37 | >0.05 |
| Intermediate-temporal | 55.56 ± 3.99 | 55.30 ± 3.68 | >0.05 |
| Intermediate-nasal | 50.13 ± 4.50 | 50.10 ± 4.00 | >0.05 |
| Inferior-temporal | 58.75 ± 4.79 | 57.06 ± 4.27 | 0.03 |
| Inferior-middle | 59.79 ± 4.38 | 59.03 ± 4.14 | >0.05 |
| Inferior-nasal | 46.75 ± 4.53 | 45.79 ± 4.17 | >0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serra, R.; D’Amico Ricci, G.; Dore, S.; Coscas, F.; Pinna, A. Evaluation of Radial Peripapillary Capillary Density in G6PD Deficiency: An OCT Angiography Pilot Study. J. Clin. Med. 2022, 11, 3282. https://doi.org/10.3390/jcm11123282
Serra R, D’Amico Ricci G, Dore S, Coscas F, Pinna A. Evaluation of Radial Peripapillary Capillary Density in G6PD Deficiency: An OCT Angiography Pilot Study. Journal of Clinical Medicine. 2022; 11(12):3282. https://doi.org/10.3390/jcm11123282
Chicago/Turabian StyleSerra, Rita, Giuseppe D’Amico Ricci, Stefano Dore, Florence Coscas, and Antonio Pinna. 2022. "Evaluation of Radial Peripapillary Capillary Density in G6PD Deficiency: An OCT Angiography Pilot Study" Journal of Clinical Medicine 11, no. 12: 3282. https://doi.org/10.3390/jcm11123282
APA StyleSerra, R., D’Amico Ricci, G., Dore, S., Coscas, F., & Pinna, A. (2022). Evaluation of Radial Peripapillary Capillary Density in G6PD Deficiency: An OCT Angiography Pilot Study. Journal of Clinical Medicine, 11(12), 3282. https://doi.org/10.3390/jcm11123282