
mTOR Inhibitor Treatment in Patients with Tuberous Sclerosis Complex Is Associated with Specific Changes in microRNA Serum Profile

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Molecular Methods
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Curatolo, P.; Moavero, R.; Roberto, D.; Graziola, F. Genotype/Phenotype Correlations in Tuberous Sclerosis Complex. Semin. Pediatr. Neurol. 2015, 22, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Manning, B.D. The TSC1–TSC2 complex: A molecular switchboard controlling cell growth. Biochem. J. 2008, 412, 179–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northrup, H.; Krueger, D.A.; Roberds, S.; Smith, K.; Sampson, J.; Korf, B.; Kwiatkowski, D.J.; Mowat, D.; Nellist, M.; Povey, S.; et al. Tuberous Sclerosis Complex Diagnostic Criteria Update: Recommendations of the 2012 International Tuberous Sclerosis Complex Consensus Conference. Pediatr. Neurol. 2013, 49, 243–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, D.N.; Belousova, E.D.; Sparagana, S.; Bebin, E.M.; Frost, M.D.; Kuperman, R.; Witt, O.; Kohrman, M.H.; Flamini, J.R.; Wu, J.Y.; et al. Long-Term Use of Everolimus in Patients with Tuberous Sclerosis Complex: Final Results from the EXIST-1 Study. PLoS ONE 2016, 11, e0158476. [Google Scholar] [CrossRef]
- Bissler, J.J.; Franz, D.N.; Frost, M.D.; Belousova, E.; Bebin, E.M.; Sparagana, S.; Berkowitz, N.; Ridolfi, A.; Kingswood, J.C. The effect of everolimus on renal angiomyolipoma in pediatric patients with tuberous sclerosis being treated for subependymal giant cell astrocytoma. Pediatr. Nephrol. 2017, 33, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Bissler, J.J.; McCormack, F.X.; Young, L.R.; Elwing, J.M.; Chuck, G.; Leonard, J.M.; Schmithorst, V.J.; Laor, T.; Brody, A.S.; Bean, J.; et al. Sirolimus for angiomyolipoma in tuberous sclerosis complex or lymphangioleiomyomatosis. N. Eng. J. Med. 2008, 358, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Mendell, J.T.; Olson, E.N. MicroRNAs in stress signaling and human disease. Cell 2012, 148, 1172–1187. [Google Scholar] [CrossRef] [Green Version]
- Santulli, G.; Totary-Jain, H. Tailoring mTOR-based therapy: Molecular evidence and clinical challenges. Pharmacogenomics 2013, 14, 1517–1526. [Google Scholar] [CrossRef] [Green Version]
- Trelinska, J.; Fendler, W.; Dachowska, I.; Kotulska, K.; Jozwiak, S.; Antosik, K.; Gnys, P.; Borowiec, M.; Mlynarski, W. Abnormal serum microRNA profiles in tuberous sclerosis are normalized during treatment with everolimus: Possible clinical implications. Orphanet J. Rare Dis. 2016, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Bąbol-Pokora, K.; Bielska, M.; Bobeff, K.; Jatczak-Pawlik, I.; Borkowska, J.; Kotulska, K.; Jóźwiak, S.; Młynarski, W.; Trelińska, J. A multistep approach to the genotype-phenotype analysis of Polish patients with tuberous sclerosis complex. Eur. J. Med. Genet. 2021, 64, 104309. [Google Scholar] [CrossRef]
- Blondal, T.; Jensby Nielsen, S.; Baker, A.; Andreasen, D.; Mouritzen, P.; Wrang Teilum, M.; Dahlsveen, I.K. Assessing sample and miRNA profile quality in serum and plasma or other biofluids. Methods 2013, 59, S1–S6. [Google Scholar] [CrossRef]
- Grabia, S.; Smyczynska, U.; Pagacz, K.; Fendler, W. NormiRazor: Tool applying GPU-accelerated computing for determination of internal references in microRNA transcription studies. BMC Bioinform. 2020, 21, 425. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Cai, Y.; Guo, H.; Li, H.Z.; Wang, W.D.; Zhang, Y.S. MicroRNA differential expression profile in tuberous sclerosis complex cell line TSC2−/− MEFs and normal cell line TSC2 +/+ MEFs. Beijing Da Xue Xue Bao 2017, 49, 580–584. [Google Scholar]
- Wang, W.X.; Prajapati, P.; Vekaria, H.; Spry, M.; Cloud, A.; Sullivan, P.; Springer, J. Temporal changes in inflammatory mitochondria-enriched microRNAs following traumatic brain injury and effects of miR-146a nanoparticle delivery. Neural Regen. Res. 2021, 16, 514. [Google Scholar] [CrossRef]
- Boese, A.S.; Saba, R.; Campbell, K.; Majer, A.; Medina, S.; Burton, L.; Booth, T.F.; Chong, P.; Westmacott, G.; Dutta, S.M.; et al. MicroRNA abundance is altered in synaptoneurosomes during prion disease. Mol. Cell. Neurosci. 2016, 71, 13–24. [Google Scholar] [CrossRef]
- Prajapati, P.; Wang, W.X.; Pesina, S.A.; Geleta, U.; Springer, J.E. Sex-Specific Alterations in Inflammatory MicroRNAs in Mouse Brain and Bone Marrow CD11b+ Cells Following Traumatic Brain Injury. Cell. Mol. Neurobiol. 2021. ahead-of-print. Available online: https://pubmed.ncbi.nlm.nih.gov/34761332/ (accessed on 19 April 2022).
- Yang, X.; Xie, J.; Jia, L.; Liu, N.; Liang, Y.; Wu, F.; Liang, B.; Li, Y.; Wang, J.; Sheng, C.; et al. Analysis of miRNAs Involved in Mouse Brain Damage upon Enterovirus 71 Infection. Front. Cell. Infect. Microbiol. 2017, 7, 133. [Google Scholar] [CrossRef]
- Goswami, S.; Banerjee, A.; Kumari, B.; Bandopadhyay, B.; Bhattacharya, N.; Basu, N.; Vrati, S.; Banerjee, A. Differential Expression and Significance of Circulating microRNAs in Cerebrospinal Fluid of Acute Encephalitis Patients Infected with Japanese Encephalitis Virus. Mol. Neurobiol. 2017, 54, 1541–1551. [Google Scholar] [CrossRef]
- Navarro-Villarán, E.; de la Cruz-Ojeda, P.; Contreras, L.; González, R.; Negrete, M.; Rodríguez-Hernández, M.A.; Marín-Gómez, L.M.; Álamo-Martínez, J.M.; Calvo, A.; Gómez-Bravo, M.A.; et al. Molecular Pathways Leading to Induction of Cell Death and Anti-Proliferative Properties by Tacrolimus and mTOR Inhibitors in Liver Cancer Cells. Cell. Physiol. Biochem. 2020, 54, 457–473. [Google Scholar]
- Zhang, Y.; Gao, M.; Chen, L.; Zhou, L.; Bian, S.; Lv, Y. Licochalcone A restrains microphthalmia-associated transcription factor expression and growth by activating autophagy in melanoma cells via miR-142-3p/Rheb/mTOR pathway. Phytother. Res. 2020, 34, 349–358. [Google Scholar] [CrossRef]
- Cukovic, D.; Bagla, S.; Ukasik, D.; Stemmer, P.M.; Jena, B.P.; Naik, A.R.; Sood, S.; Asano, E.; Luat, A.; Chugani, D.C.; et al. Exosomes in epilepsy of tuberous sclerosis complex: Carriers of pro-inflammatory micrornas. Non-Coding RNA 2021, 7, 40. [Google Scholar] [CrossRef]
- Zhong, Y.; Yu, C.; Qin, W. LncRNA SNHG14 promotes inflammatory response induced by cerebral ischemia/reperfusion injury through regulating miR-136-5p/ROCK1. Cancer Gene Ther. 2019, 26, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, D.N.; Lawson, J.A.; Yapici, Z.; Ikeda, H.; Polster, T.; Nabbout, R.; Curatolo, P.; de Vries, P.J.; Dlugos, D.J.; Voi, M.; et al. Everolimus for treatment-refractory seizures in TSC: Extension of a randomized controlled trial. Neurol. Clin. Pract. 2018, 8, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Tyburczy, M.E.; Kotulska, K.; Pokarowski, P.; Mieczkowski, J.; Kucharska, J.; Grajkowska, W.; Roszkowski, M.; Jozwiak, S.; Kaminska, B. Novel proteins regulated by mTOR in subependymal giant cell astrocytomas of patients with tuberous sclerosis complex and new therapeutic implications. Am. J. Pathol. 2010, 176, 1878–1890. [Google Scholar] [CrossRef]
- Mi, R.; Ma, J.; Zhang, D.; Li, L.; Zhang, H. Efficacy of combined inhibition of mTOR and ERK/MAPK pathways in treating a tuberous sclerosis complex cell model. J. Genet. Genom. 2009, 36, 355–361. [Google Scholar] [CrossRef]
- Nguyen, L.H.; Leiser, S.C.; Song, D.; Brunner, D.; Roberds, S.L.; Wong, M.B.A. Inhibition of MEK-ERK signaling reduces seizures in two mouse models of tuberous sclerosis complex. Epilepsy Res. 2022, 181, 106890. [Google Scholar] [CrossRef]
- Bongaarts, A.; Van Scheppingen, J.; Korotkov, A.; Mijnsbergen, C.; Anink, J.J.; Jansen, F.E.; Spliet, W.G.M.; Den Dunnen, W.F.A.; Gruber, V.E.; Scholl, T.; et al. The coding and non-coding transcriptional landscape of subependymal giant cell astrocytomas. Brain 2020, 143, 131–149. [Google Scholar] [CrossRef] [Green Version]

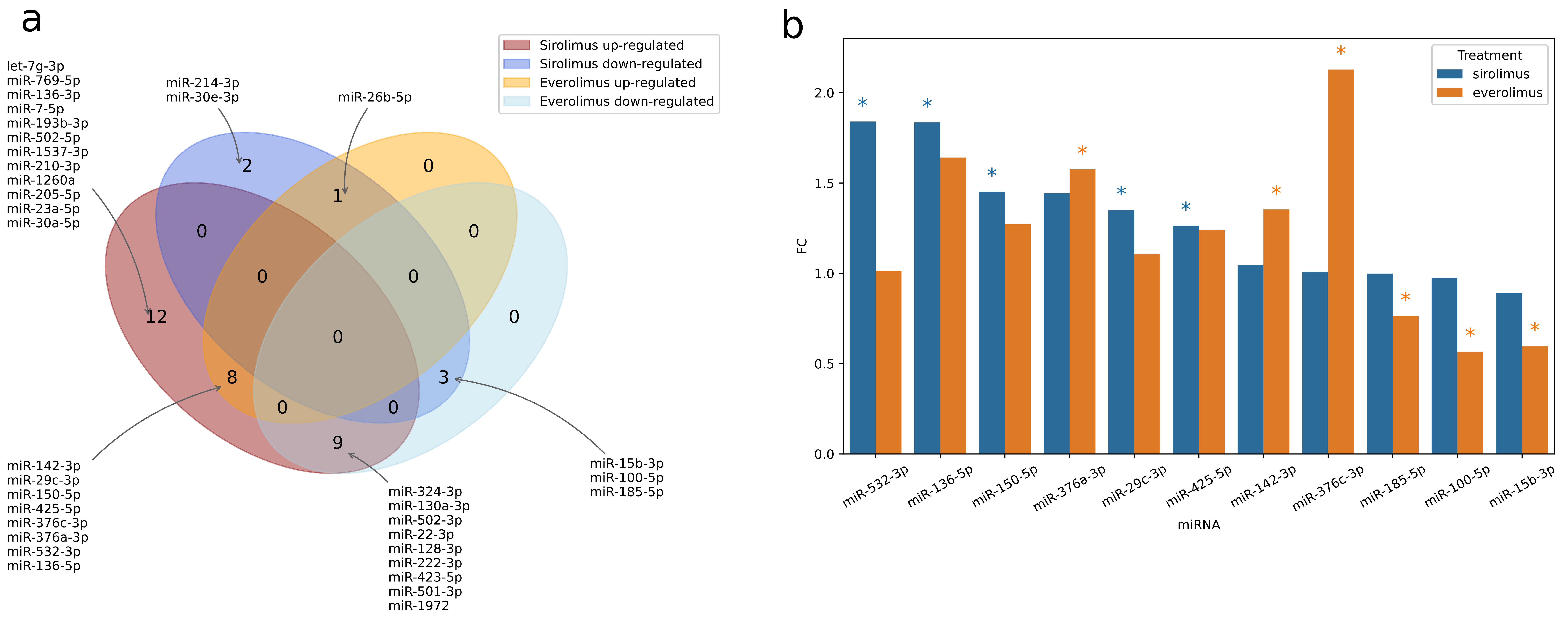


{kind=link}
{kind=link}
{kind=link}
| Whole Group (N = 10) | TSC2 (N = 7) | TSC1 (N = 2) | No Mutation (N = 1) | |
|---|---|---|---|---|
| Sex (M/F) | 5/5 | 6/1 | 1/1 | 1 |
| Age (years) | 10.4 ± 4.3 | 10.3 ± 4.8 | 8.6 ± 3.3 | 14.0 |
| SEGA tumor | 5/10 | 3/7 | 1/2 | 1 |
| SEN | 8/10 | 5/7 | 2/2 | 1 |
| Cortical dysplasia | 4/10 | 3/7 | 0/2 | 1 |
| ||||
| 6/10 | 4/7 | 2/2 | 0 |
| 0/10 | 0/7 | 0/2 | 0 |
| 7/10 | 5/7 | 2/2 | 0 |
| 3/10 | 3/7 | 0/2 | 0 |
| ||||
| 6/10 | 5/7 | 1/2 | 0 |
| 7/10 | 3/7 | 0/2 | 0 |
| 0/10 | 0/7 | 0/2 | 0 |
| 0/10 | 0/7 | 0/2 | 0 |
| TSC-associated neuropsychiatric disorders | 7/10 | 5/7 | 2/2 | 0 |
| Epilepsy | 4/10 | 4/7 | 0/2 | 0 |
| Target Prediction Method/Database | |||
|---|---|---|---|
| Gene | miR-136-5p | miR-150-5p | miR-376a-3p |
| LONP2 | - | - | - |
| DGKI | TargetScan | TargetScan | - |
| SMAD2 | - | - | - |
| UQCRB | - | - | - |
| PCSK5 | - | - | miRDB |
| BDNF | - | - | - |
| CCNA2 | - | - | - |
| AMOT | - | - | - |
| P2RY2 | TargetScan | - | - |
| FBXO22 | - | - | - |
| KEGG Pathway Name | p-Value | FDR-Adjusted p-Value |
|---|---|---|
| Ras signaling pathway | 0 | 0 |
| MAPK signaling pathway | 0.0006 | 0.0402 |
| Phospholipase D signaling pathway | 0.0005 | 0.0402 |
| Pathways in cancer | 0.0006 | 0.0402 |
| Rap1 signaling pathway | 0.0010 | 0.0439 |
| Longevity regulating pathway | 0.0010 | 0.0439 |
| Hedgehog signaling pathway | 0.0013 | 0.0439 |
| Neurotrophin signaling pathway | 0.0017 | 0.0439 |
| Glutamatergic synapse | 0.0016 | 0.0439 |
| GABAergic synapse | 0.0018 | 0.0439 |
| Thyroid hormone signaling pathway | 0.0017 | 0.0439 |
| EGFR tyrosine kinase inhibitor resistance | 0.0027 | 0.0441 |
| Longevity regulating pathway | 0.0026 | 0.0441 |
| Cholinergic synapse | 0.0021 | 0.0441 |
| Inflammatory mediator regulation of TRP channels | 0.0025 | 0.0441 |
| Transcriptional misregulation in cancer | 0.0022 | 0.0441 |
| Glioma | 0.0028 | 0.0441 |
| AMPK signaling pathway | 0.0032 | 0.0476 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pawlik, B.; Smyczyńska, U.; Grabia, S.; Fendler, W.; Dróżdż, I.; Bąbol-Pokora, K.; Kotulska, K.; Jóźwiak, S.; Borkowska, J.; Młynarski, W.; et al. mTOR Inhibitor Treatment in Patients with Tuberous Sclerosis Complex Is Associated with Specific Changes in microRNA Serum Profile. J. Clin. Med. 2022, 11, 3395. https://doi.org/10.3390/jcm11123395
Pawlik B, Smyczyńska U, Grabia S, Fendler W, Dróżdż I, Bąbol-Pokora K, Kotulska K, Jóźwiak S, Borkowska J, Młynarski W, et al. mTOR Inhibitor Treatment in Patients with Tuberous Sclerosis Complex Is Associated with Specific Changes in microRNA Serum Profile. Journal of Clinical Medicine. 2022; 11(12):3395. https://doi.org/10.3390/jcm11123395
Chicago/Turabian StylePawlik, Bartłomiej, Urszula Smyczyńska, Szymon Grabia, Wojciech Fendler, Izabela Dróżdż, Katarzyna Bąbol-Pokora, Katarzyna Kotulska, Sergiusz Jóźwiak, Julita Borkowska, Wojciech Młynarski, and et al. 2022. "mTOR Inhibitor Treatment in Patients with Tuberous Sclerosis Complex Is Associated with Specific Changes in microRNA Serum Profile" Journal of Clinical Medicine 11, no. 12: 3395. https://doi.org/10.3390/jcm11123395
APA StylePawlik, B., Smyczyńska, U., Grabia, S., Fendler, W., Dróżdż, I., Bąbol-Pokora, K., Kotulska, K., Jóźwiak, S., Borkowska, J., Młynarski, W., & Trelińska, J. (2022). mTOR Inhibitor Treatment in Patients with Tuberous Sclerosis Complex Is Associated with Specific Changes in microRNA Serum Profile. Journal of Clinical Medicine, 11(12), 3395. https://doi.org/10.3390/jcm11123395