Promising Advances in Pharmacotherapy for Patients with Spinal Cord Injury—A Review of Studies Performed In Vivo with Modern Drugs

 , , ,
, , , {kind=link}
{kind=link}
Abstract
:1. Introduction
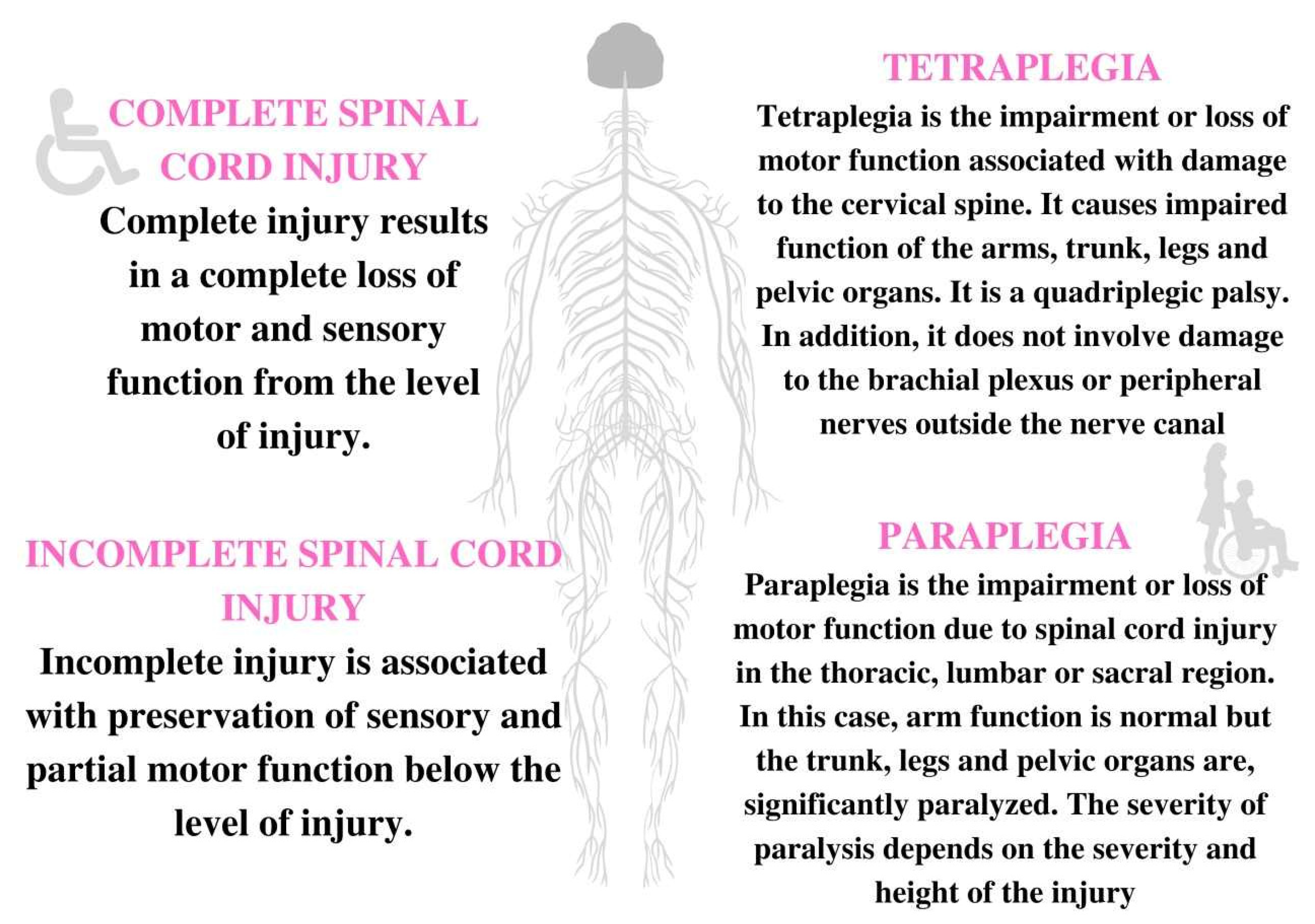
2. Etiology and Types of SCI
3. Inflammation in SCI
3.1. The Role of Microglia
3.2. Bioactive Mediators of Inflammation
3.3. Chemokine-Receptor Ligand System
3.4. Heme Oxygenases (HO)
3.5. Apelin-13
3.6. Nanotherapeutics
3.7. Ferulic Acid (FA)
3.8. Neuroinflammation
4. Oxidative Stress and Neuropathic Pain in SCI
4.1. The Causes of NP in SCI
4.2. Pharmacotherapy NP
4.3. Groups of Drugs with Potential Use in the Treatment/Control of NP in SCI
4.3.1. Coanalgesics
4.3.2. Cannabinoids
4.3.3. NMDA Receptor Antagonists
4.3.4. Antioxidants
4.3.5. Other Modification of Genetic Pathways
5. Neuro-Regeneration, Neuroprotection, and Spinal Cord Plasticity
5.1. Biological Therapy
5.2. Phosphodiesterase Inhibitors
5.3. Immunosuppressive Therapy
5.4. RhoA/ROCK Inhibitors
5.5. Therapy Based on the mTOR Pathway
5.6. Therapy at the Sigma-1 Receptor
5.7. Other Possible Drugs
6. Spasticity and Muscle Loss
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| ABRT-scES | Activity-based recovery training epidural stimulation of the spinal cord |
| BTX-A | Botulinum toxin A |
| ChABC | Chondroitinase ABC enzyme |
| CNS | Central nervous system |
| CSPG | Chondroitin sulfate proteoglycans |
| EAS | External anal sphincter |
| ENSC | Intestinal neutral stem cells |
| hIL6 | Fusion protein of the cytokine interleukin-6 and the soluble interleukin-6 receptor, which are covalently linked by a flexible peptide linker |
| LUT | Functional impairment of the lower urinary tract |
| LV | Lentiwirus |
| mTOR kinase | Threonine-serine protein kinase, whose function is to regulate cell growth, proliferation, and movement, as well as translation and transcription processes |
| NDO | Detrusor neurogenic hyperactivity |
| OAB | Overactive bladder |
| onaBoNTA | OnabotulinumtoxinA |
| PC protein | Biochemically serine proteinase is a physiological blood anticoagulant, inactivating active factors VIIIa and Va and thus inhibiting thrombin and clot fibril formation |
| PTEN | The PTEN gene belongs to the suppressor genes, which means that its protein product is involved in cell cycle regulation |
| SCC | Spinal cord crush |
| scES | Epidural stimulation of the spinal cord |
| SCI | Spinal cord injury |
| TNF-alpha | Tumor necrosis factor alpha |
| TrkB, 29D7 | Antibody that binds to the TRKB receptor, matched by a key-clamp model |
| TRKD receptor | Tropomyosin B receptor kinase |
References
- O’Shea, T.M.; Burda, J.E.; Sofroniew, M.V. Cell biology of spinal cord injury and repair. J. Clin. Investig. 2017, 127, 3259–3270. [Google Scholar] [CrossRef] [PubMed]
- Anjum, A.; Yazid, M.D.; Daud, M.F.; Idris, J.; Ng, A.M.H.; Naicker, A.S.; Ismail, O.H.R.; Kumar, R.K.A.; Lokanathan, Y. Spinal Cord Injury: Pathophysiology, Multimolecular Interactions, and Underlying Recovery Mechanisms. Int. J. Mol. Sci. 2020, 21, 7533. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tang, Y.; Vogel, L.C.; DeVivo, M.J. Causes of spinal cord injury. Top. Spinal Cord Inj. Rehabil. 2013, 19, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef] [Green Version]
- Devilee, R.; Sanders, R.; de Lange, S. Treatment of fractures and dislocations of the thoracic and lumbar spine by fusion and Harrington instrumentation. Arch. Orthop. Trauma Surg. 1995, 114, 100–102. [Google Scholar] [CrossRef]
- Zhao, L.; Shen, J.; Jia, K.; Shi, F.; Hao, Q.; Gao, F. MicroRNA-24-3p Inhibits Microglia Inflammation by Regulating MK2 Following Spinal Cord Injury. Neurochem. Res. 2020, 46, 843–852. [Google Scholar] [CrossRef]
- Torg, J.S.; Corcoran, T.A.; Thibault, L.E.; Pavlov, H.; Sennett, B.J.; Naranja, R.J., Jr.; Priano, S. Cervical cord neurapraxia: Classification, pathomechanics, morbidity, and management guidelines. J. Neurosurg. 1997, 87, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.J.; Auguste, K.I.; Sun, P.P. Cervical spinal stenosis and sports-related cervical cord neurapraxia. Neurosurg. Focus. 2011, 5, E7. [Google Scholar] [CrossRef] [Green Version]
- Aarabi, B.; Alibaii, E.; Taghipur, M.; Kamgarpur, A. A comparative study of functional recovery after surgically explored and conservatively treated rocket spinal cord injuries. Neurosurgery 1996, 39, 1133–1140. [Google Scholar] [CrossRef]
- Sidhu, G.S.; Ghag, A.; Prokuski, V.; Vaccaro, A.R.; Radcliff, K.Y. Spinal cord injuries from civilian gunshots: A systematic review of the current literature. Clin. Orthop. Relat. Res. 2013, 471, 3945–3955. [Google Scholar] [CrossRef]
- LaPlaca, M.; Simon, C.; Prado, G.; Cullen, D. CNS injury biomechanics and experimental models. Wallach. Brain Res. 2007, 161, 13–26. [Google Scholar]
- Hall, E.D.; Wolf, D.L. A pharmacological analysis of the pathological mechanisms of post traumatic spinal cord ischemia. J. Neurosurg. 1986, 64, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, K.L. Care of the patient living with spinal cord injury. Nursing 2003, 33, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Nas, K.; Yazmalar, L.; Şah, V.; Aydın, A.; Öneş, K. Rehabilitation after spinal cord injury. World J. Orthop. 2015, 6, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chen, W.; Liu, K.; Ma, P.; Qiu, P.; Zheng, C.; Zhang, X.; Tan, P.; Xi, X.; He, X. Mitigation of Microglia-mediated Acute Neuroinflammation and Tissue Damage by Heme Oxygenase 1 in a Rat Spinal Cord Injury Model. Neuroscience 2021, 457, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Pelisch, N.; Rosas Almanza, J.; Stehlik, K.E.; Aperi, B.V.; Kroner, A. Use of a Self-Delivering Anti-CCL3 FANA Oligonucleotide as an Innovative Approach to Target Inflammation after Spinal Cord Injury. eNeuro 2021, 8, ENEURO.0338-20.2021. [Google Scholar] [CrossRef]
- Shen, K.G.; Chan, L.; He, L.; Li, X.; Yang, S.; Wang, B.; Zhang, H.; Huang, J.; Chang, M.; Li, Z.; et al. Anti-Inflammatory Nanotherapeutics by Targeting Matrix Metalloproteinases for Immunotherapy of Spinal Cord Injury. Small 2021, 17, 41. [Google Scholar] [CrossRef]
- David, S.; López-Vales, R. Bioactive Lipid Mediators in the Initiation and Resolution of Inflammation after Spinal Cord Injury. Neuroscience 2021, 466, 273–297. [Google Scholar] [CrossRef]
- Vafaei-Nezhad, S.; Niknazar, S.; Norouzian, M.; Abdollahifar, M.A.; Aliaghaei, A.; Abbaszadeh, H.A. Therapeutics effects of [Pyr1] apelin-13 on rat contusion model of spinal cord injury: An experimental study. J. Chem. Neuroanat. 2021, 113, 101924. [Google Scholar] [CrossRef]
- Borgonetti, V.; Galeotti, N. Combined inhibition of histone deacetylases and BET family proteins as epigenetic therapy for nerve injury-induced neuropathic pain. Pharmacol. Res. 2021, 165, 105431. [Google Scholar] [CrossRef]
- Jiang, X.; Yu, X.; Chen, Y.; Jing, C.; Xu, L.; Chen, Z.; Liu, F.; Chen, L. Ferulic Acid Improves Motor Function Induced by Spinal Cord Injury in Rats via Inhibiting Neuroinflammation and Apoptosis. Acta Cir. Bras. 2021, 36, 7. [Google Scholar] [CrossRef] [PubMed]
- Cheung, V.; Hoshide, R.; Bansal, V.; Kasper, E.; Chen, C.C. Methylprednisolone in the management of spinal cord injuries: Lessons from randomized, controlled trials. Surg. Neurol. Int. 2015, 24, 142. [Google Scholar] [CrossRef]
- Sauerland, S.; Nagelschmidt, M.; Mallmann, P.; Neugebauer, E.A. Risks and benefits of preoperative high dose methylprednisolone in surgical patients: A systematic review. Drug Saf. 2000, 23, 449–461. [Google Scholar] [CrossRef]
- Hugenholtz, H.; Cass, D.E.; Dvorak, M.F.; Fewer, D.H.; Fox, R.J.; Izukawa, D.M.; Lexchin, J.; Tuli, S.; Bharatwal, N.; Short, C. High-dose methylprednisolone for acute closed spinal cord injury—Only a treatment option. Can. J. Neurol. Sci. 2002, 29, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, A.M.; Zhao, P.; Waxman, S.G.; Hains, B.C. Early microglial inhibition preemptively mitigates chronic pain development after experimental spinal cord injury. J. Rehab. Res. Dev. 2009, 46, 123–133. [Google Scholar] [CrossRef]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation: Emergence of the pro-resolving superfamily of mediators. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Vales, R.; Ghasemlou, N.; Redensek, A.; Kerr, B.J.; Barbayianni, E.; Antonopoulou, G.; Baskakis, C.; Rathore, K.I.; Constantinou-Kokotou, V.; Stephens, D.; et al. Phospholipase A2 superfamily members play divergent roles after spinal cord injury. FASEB J. 2011, 25, 4240–4252. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.N.; Huang, W.; Hall, J.C.; Michael-Titus, A.T.; Priestley, J.V. Improved outcome after spinal cord compression injury in mice treated with docosahexaenoic acid. Exp. Neurol. 2013, 239, 13–27. [Google Scholar] [CrossRef]
- Paterniti, I.; Impellizzeri, D.; Di Paola, R.; Esposito, E.; Gladman, S.; Yip, P.; Priestley, J.V.; Michael-Titus, A.T.; Cuzzocrea, S. Docosahexaenoic acid attenuates the early inflammatory response following spinal cord injury in mice: In-vivo and in-vitro studies. J. Neuroinflamm. 2014, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.L.; King, V.R.; Curran, O.E.; Dyall, S.C.; Ward, R.E.; Lal, N.; Priestley, J.V.; Michael-Titus, A.T. A combination of intravenous and dietary docosahexaenoic acid significantly improves outcome after spinal cord injury. Brain 2007, 130, 3004–3019. [Google Scholar] [CrossRef] [Green Version]
- Banisor, I.; Leist, T.P.; Kalman, B. Involvement of beta-chemokines in the development of inflammatory demyelination. J. Neuroinflamm. 2005, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Osamu, Y. Chemokines: A new classification system and their role in immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Martín-Pintado, N.; Yahyaee-Anzahaee, M.; Campos-Olivas, R.; Noronha, A.M.; Wilds, C.J.; Damha, M.J.; González, C. The solution structure of double helical arabino nucleic acids (ANA and 2’F-ANA): Effect of arabinoses in duplex-hairpin interconversion. Nucleic Acids Res. 2012, 40, 9329–9339. [Google Scholar] [CrossRef] [PubMed]
- Denisov, A.Y.; Noronha, A.M.; Wilds, C.J.; Trempe, J.F.; Pon, R.T.; Gehring, K.; Damha, M.J. Solution structure of an arabinonucleic acid (ANA)/RNA duplex in a chimeric hairpin: Comparison with 2’-fluoro-ANA/RNA and DNA/RNA hybrids. Nucleic Acids Res. 2001, 29, 4284–4293. [Google Scholar] [CrossRef] [Green Version]
- Paine, A.; Eiz-Vesper, B.; Blasczyk, R.; Immenschuh, S. Signaling to heme oxygenase-1 and its anti-inflammatory therapeutic potential. Biochem. Pharmacol. 2010, 80, 1895–1903. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Sánchez, J.; Chánez-Cárdenas, M.E. A review on hemeoxygenase-2: Focus on cellular protection and oxygen response. Oxid. Med. Cell. Long. 2014, 2014, 604981. [Google Scholar] [CrossRef] [Green Version]
- Haghparast, E.; Sheibani, V.; Abbasnejad, M.; Esmaeili-Mahani, S. Apelin-13 attenuates motor impairments and prevents the changes in synaptic plasticity-related molecules in the striatum of Parkinsonism rats. Peptides 2019, 117, 170091. [Google Scholar] [CrossRef]
- Masoumi, J.; Abbasloui, M.; Parvan, R.; Mohammadnejad, D.; Pavon-Djavid, G.; Barzegari, A.; Abdolalizadeh, J. Apelin, a promising target for Alzheimer disease prevention and treatment. Neuropeptides 2018, 70, 76–86. [Google Scholar] [CrossRef]
- Xin, Q.; Cheng, B.; Pan, Y.; Liu, H.; Yang, C.; Chen, Y.; Bai, B. Neuroprotective effects of apelin-13 on experimental ischemic stroke through suppression of inflammation. Peptides 2015, 63, 55–62. [Google Scholar] [CrossRef]
- Chan, L.; Huang, Y.; Chen, T. Cancer-targeted tri-block copolymer nanoparticles as payloads of metal complexes to achieve enhanced cancer theranosis. J. Mater. Chem. B 2016, 4, 4517–4525. [Google Scholar] [CrossRef]
- Sosnik, A.; Mühlebach, S. Drug Nanoparticles and Nano-Cocrystals: From Production and Characterization to Clinical Translation. Adv. Drug Deliv. Rev. 2018, 131, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Haijian, X.; Dan, C.; Zhengbu, L.; Yi, Y. Mechanisms of Autophagy and Apoptosis Mediated by JAK2 Signaling Pathway after Spinal Cord Injury of Rats. Exp. Ther. Med. 2017, 14, 1589–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Yao, X.; Sun, B.; Jiang, W.; Liao, C.; Dai, X.; Chen, Y.; Chen, J.; Ding, R. Pretreatment with Kaempferol Attenuates Microglia-Mediate Neuroinflammation by Inhibiting MAPKs-NF-ΚB Signaling Pathway and Pyroptosis after Secondary Spinal Cord Injury. Free Radic. Biol. Med. 2021, 168, 142–154. [Google Scholar] [CrossRef]
- Zhou, H.J.; Wang, L.Q.; Xu, Q.S.; Fan, Z.X.; Zhu, Y.; Jiang, H.; Zheng, X.J.; Ma, Z.Y.; Zhan, R.Y. Downregulation of miR-199b promotes the acute spinal cord injury through IKKbeta-NF-kappaB signaling pathway activating microglial cells exp. Cell Res. 2016, 349, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Karpińska, A.; Gromadzka, G. Oxidative Stress and Natural Antioxidant Mechanisms: The Role in Neurodegeneration. From Molecular Mechanisms to Therapeutic Strategies. Postepy Hig. I Med. Dosw. 2013, 67, 45–53. [Google Scholar] [CrossRef]
- Mata-Bermudez, A.; Diaz-Ruiz, A.; Burelo, M.; García-Martínez, B.A.; Jardon-Guadarrama, G.; Calderón-Estrella, F.; Rangel-Hernández, A.; Pérez-González, C.; Camilo, R. Dapsone Prevents Allodynia and Hyperalgesia and Decreased Oxidative Stress After Spinal Cord Injury in Rats. Spine 2021, 46, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Mata-Bermudez, A.; Ríos, C.; Burelo, M.; Pérez-González, C.; García-Martínez, B.A.; Jardon-Guadarrama, G.; Calderón-Estrella, F.; Manning-Balpuesta, N.; Diaz-Ruiz, A. Amantadine Prevented Hypersensitivity and Decreased Oxidative Stress by NMDA Receptor Antagonism after Spinal Cord Injury in Rats. Eur. J. Pain 2021, 25, 1839–1851. [Google Scholar] [CrossRef]
- Santos, M.C.Q.; da Silva, T.C.B.; da Silva, F.B.O.; Siebert, C.; Kroth, A.; Silveira, E.M.S.; Wyse, A.T.S.; Partata, W.A. Effects of Vitamin D Administration on Nociception and Spinal Cord Pro-Oxidant and Antioxidant Markers in a Rat Model of Neuropathic Pain. Braz. J. Med. Biol. Res. 2021, 54, e11207. [Google Scholar] [CrossRef]
- Davari, M.; Amani, B.; Amani, B.; Khanijahani, A.; Akbarzadeh, A.; Shabestan, R. Pregabalin and gabapentin in neuropathic pain management after spinal cord injury: A systematic review and meta-analysis. Korean J. Pain 2020, 33, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Alles Sascha, R.A.; Cain Stuart, M.; Snutch, T. Pregabalin as a pain therapeutic: Beyond calcium channels. Front. Cell. Neurosci. 2020, 14, 83. [Google Scholar] [CrossRef] [Green Version]
- Tong, C.; Zhengyao, Z.; Mei, L.; Dongpo, S.; Qian, H.; Fengqun, M. Pregabalin and Gabapentin in Patients with Spinal Cord Injury-Related Neuropathic Pain: A Network Meta-Analysis. Pain Ther. 2021, 10, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Evoy, K.E.; Sadrameli, S.; Contreras, J.; Covvey, J.R.; Peckham, A.M.; Morrison, M.D. Abuse and Misuse of Pregabalin and Gabapentin: A Systematic Review Update. Drugs 2021, 81, 125–156. [Google Scholar] [CrossRef] [PubMed]
- Kukkar, A.; Bali, A.; Singh, N.; Jaggi, A.S. Implications and mechanism of action of gabapentin in neuropathic pain. Arch. Pharm. Res. 2013, 36, 237–251. [Google Scholar] [CrossRef]
- Wiffen, P.J.; Derry, S.; Bell, R.F.; Rice, A.S.; Tölle, T.R.; Phillips, T.; Moore, R.A. Gabapentin for chronic neuropathic pain in adults. Cochrane Database Syst Rev. 2017, 9, CD007938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.D.; Chaplan, S.R.; Higuera, E.S.; Sorkin, L.S.; Stauderman, K.A.; Williams, M.E.; Yaksh, T.L. Upregulation of dorsal root ganglion (alpha)2(delta) calcium channel subunit and its correlation with allodynia in spinal nerve-injured rats. J. Neurosci. 2001, 21, 1868–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnerup, N.B.; Attal, N. Pharmacotherapy of neuropathic pain: Time to rewrite the rulebook? Pain Manag. 2016, 6, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.; et al. Neuropathic pain. Nat. Rev. Dis. Primers. 2017, 16, 17002. [Google Scholar] [CrossRef] [Green Version]
- Ghanavatian, S.; Derian, A. Tizanidine; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK519505/ (accessed on 5 September 2022).
- Semenchuk, M.R.; Sherman, S. Effectiveness of tizanidine in neuropathic pain: An open-label study. J. Pain 2000, 1, 285–292. [Google Scholar] [CrossRef]
- Nabata, K.J.; Tse, E.K.; Nightingale, T.E. The Therapeutic potential and usage patterns of cannabinoid use in patients with spinal cord injury: A systematic review. Curr. Neuropharmacol. 2021, 19, 402–432. [Google Scholar] [CrossRef]
- Huestis, M.A.; Solimini, R.; Pichini, S.; Pacifici, R.; Carlier, J.; Busardò, F.P. Cannabidiol Adverse Effects and Toxicity. Curr. Neuropharmacol. 2019, 17, 974–989. [Google Scholar] [CrossRef] [PubMed]
- Neale, M. Efficacy and safety of cannabis for treating children with refractory epilepsy. Nurs. Child Young People 2017, 11, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Thiele, E.A.; Marsh, E.D.; French, J.A.; Mazurkiewicz-Beldzinska, M.; Benbadis, S.R.; Joshi, C.; Lyons, P.D.; Taylor, A.; Roberts, C.; Sommerville, K. Cannabidiol in patients with seizures associated with Lennox-Gastaut syndrome (GWPCARE4): A randomised, double-blind, placebo-controlled phase 3 trial. Lancet 2018, 17, 1085–1096. [Google Scholar] [CrossRef]
- Norberg, M.M.; Kavanagh, D.J.; Olivier, J.; Lyras, S. Craving cannabis: A meta-analysis of self-report and psychophysiological cue-reactivity studies. Addiction 2016, 111, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- Filbey, F.M.; Dunlop, J.; Ketcherside, A.; Baine, J.; Rhinehardt, T.; Kuhn, B.; DeWitt, S.; Alvi, T. fMRI study of neural sensitization to hedonic stimuli in long-term, daily cannabis users. Hum. Brain Mapp. 2016, 37, 3431–3443. [Google Scholar] [CrossRef]
- Banafshe, H.R.; Khoshnoud, M.J.; Abed, A.; Saghazadeh, M.; Mesdaghinia, A. Vitamin D supplementation attenuates the behavioral scores of neuropathic pain in rats. Nutr. Neurosci. 2019, 10, 700–705. [Google Scholar] [CrossRef]
- Zang, C.; Ma, S.; Zhao, X.; Wen, B.; Sun, P.; Fu, Z. Upregulation of Antioxidant and Autophagy Pathways via NRF2 Activation Protects Spinal Cord Neurons from Ozone Damage. Mol. Med. Rep. 2021, 23, 6. [Google Scholar]
- Ercan, S.; Ayfer, A.; Mustafa, S.K. Antioxidative Effects of Alpha-Lipoic Acid in Spinal Cord Injury an Experimental Rat Model. Annali Ital. Chirurgia 2021, 92, 98–102. [Google Scholar]
- Wang, P.; Zhang, Y.; Xia, Y.; Xu, D.; Wang, H.; Liu, D.; Xu, S.; Sun, Y. MicroRNA-139-5p Promotes Functional Recovery and Reduces Pain Hypersensitivity in Mice with Spinal Cord Injury by Targeting Mammalian Sterile 20-like Kinase 1. Neurochem. Res. 2021, 46, 349–357. [Google Scholar] [CrossRef]
- Zhang, P.; Sun, H.; Ji, Z. Downregulating lncRNA PVT1 Relieves Astrocyte Overactivation Induced Neuropathic Pain Through Targeting miR-186-5p/CXCL13/CXCR5 Axis. Neurochem. Res. 2020, 46, 1457–1469. [Google Scholar] [CrossRef]
- Bao, Y.-N.; Dai, W.-L.; Fan, J.-F.; Bin Ma, B.; Li, S.-S.; Zhao, W.-L.; Yu, B.-Y.; Liu, J.-H. The dopamine D1-D2DR complex in the rat spinal cord promotes neuropathic pain by increasing neuronal excitability after chronic constriction injury. Exp. Mol. Med. 2021, 53, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Kang, J.; Kim, H.; Gwak, Y. The Roles of Superoxide on At-Level Spinal Cord Injury Pain in Rats. Int. J. Mol. Sci. 2021, 22, 2672. [Google Scholar] [CrossRef] [PubMed]
- Kishima, K.; Tachibana, T.; Yamanaka, H.; Kobayashi, K.; Okubo, M.; Maruo, K.; Noguchi, K. Role of Rho-associated coiled-coil containing protein kinase in the spinal cord injury induced neuropathic pain. Spine J. 2021, 21, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Dimitrijevic, M.R.; Danner, S.M.; Mayr, W. Neurocontrol of movement in humans with spinal cord injury. Art. Org. 2017, 39, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Turtle, J.D.; Henwood, M.K.; Strain, M.M.; Huang, Y.J.; Miranda, R.C.; Grau, J.W. Pain fiber involvement after spinal cord injury promotes hemorrhage and expands the area of secondary injury. Exp. Neurol. 2019, 311, 115–124. [Google Scholar] [CrossRef]
- Leibinger, M.; Zeitler, C.; Gobrecht, P.; Andreadaki, A.; Gisselmann, G.; Fischer, D. Transneuronal delivery of hyper-interleukin-6 enables functional recovery after severe spinal cord injury in mice. Nat. Commun. 2021, 12, 391. [Google Scholar] [CrossRef]
- Nakamura, M.; Okada, S.; Toyama, Y.; Okano, H. Role of IL-6 in spinal cord injury in a mouse model. Clin. Rev. Allergy Immunol. 2005, 28, 197–204. [Google Scholar] [CrossRef]
- Fouad, K.; Vavrek, R.; Cho, S. A TrkB Antibody Agonist Promotes Plasticity after Cervical Spinal Cord Injury in Adult Rats. J. Neurotrauma 2021, 38, 1338–1348. [Google Scholar] [CrossRef]
- Guan, B.; Jiang, C. Design and development of 1, 3, 5-triazine derivatives as protective agent against spinal cord injury in rat via inhibition of NF-ĸB. Bioorg. Med. Chem. Lett. 2021, 41, 127964. [Google Scholar] [CrossRef]
- Yoshizaki, S.; Tamaru, T.; Hara, M.; Kijima, K.; Tanaka, M.; Konno, D.J.; Matsumoto, Y.; Nakashima, Y.; Okada, S. Microglial inflammation after chronic spinal cord injury is enhanced by reactive astrocytes via the fibronectin/β1 integrin pathway. J. Neuroinflamm. 2021, 18, 12. [Google Scholar] [CrossRef]
- Abdallah, A.N.; Shamaa, A.A.; El-Tookhy, O.S.; Bahr, M.M. Effect of combined intrathecal/intravenous injection of bone marrow derived stromal cells in platelet-rich plasma on spinal cord injury in companion animals. Open Vet. J. 2021, 11, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Arac, D.; Erdi, M.F.; Keskin, F.; Kenan, M.; Cuce, G.; Aydemir, F.H.Y.; Guney, O.; Kocaogullar, Y. Neuroprotective Effects of Milrinone on Experimental Acute Spinal Cord Injury: Rat Model. World Neurosurg. 2020, 147, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yin, Y.; Li, L.; Zhang, Y.; He, W.; Shi, Y. Milrinone Ameliorates the Neuroinflammation and Memory Function of Alzheimer’s Disease in an APP/PS1 Mouse Model. Neuropsychiatr. Dis. Treat. 2021, 17, 2129–2139. [Google Scholar] [CrossRef]
- Bahadir, S.; Onal, M.B.; Narin, F.; Basar, I.; Yigitkanli, K.; Bavbek, M.; Ziyal, M.I. Azathioprine as a Neuroprotective Agent in Experimental Traumatic Spinal Cord Injury. Turk. Neurosurg. 2021, 31, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Horikoshi, Y.; Kuriyagawa, C.; Niiyama, Y. Rho/ROCK Pathway and non-coding RNA: Consequences of ischemic stroke and spinal cord injury inside. J. Mol. Sci. 2021, 22, 11573. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Li, Y.Q.; Lu, Y.F.; Wu, Y.; Liu, R.; Zheng, Y.R.; Yin, M. Exploring the potential of RhoA inhibitors to improve exercise-recoverable spinal cord injury: A systematic review and meta-analysis. J. Chem. Neuroanat. 2021, 111, 101879. [Google Scholar] [CrossRef]
- Roy, A.; Pathak, Z.; Kumar, H. Strategies to neutralize RhoA/ROCK pathway after spinal cord injury. Exp. Neurol. 2021, 343, 113794. [Google Scholar] [CrossRef]
- Jia, X.; Huang, G.; Wang, S.; Long, M.; Tang, X.; Feng, D.; Zhou, Q. Extracellular vesicles derived from mesenchymal stem cells containing microRNA-381 protect against spinal cord injury in a rat model via the BRD4/WNT5A axis. Bone Jt. Res. 2021, 10, 328–339. [Google Scholar] [CrossRef]
- Li, X.; Peng, Z.; Long, L.; Lu, X.; Zhu, K.; Tuo, Y.; Chen, N.; Zhao, X.; Wang, L.; Wan, Y. Transplantation of Wnt5a-modified NSCs promotes tissue repair and locomotor functional recovery after spinal cord injury. Exp. Mol. Med. 2020, 52, 2020–2033. [Google Scholar] [CrossRef]
- Stern, S.; Hilton, B.J.; Burnside, E.R.; Dupraz, S.; Handley, E.E.; Gonyer, J.M.; Brakebusch, C.; Bradke, F. RhoA drives actin compaction to restrict axon regeneration and astrocyte reactivity after CNS injury. Neuron 2021, 3, 3436–3455.e9. [Google Scholar] [CrossRef]
- Dupraz, S.; Hilton, B.J.; Husch, A.; Santos, T.E.; Coles, C.H.; Stern, S.; Brakebusch, C.; Bradke, F. RhoA Controls Axon Extension Independent of Specification in the Developing Brain. Curr. Biol. 2019, 29, 3874–3886.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Chen, Q. mTOR pathway: A potential therapeutic target for spinal cord injury. Biomed. Pharmacother. 2022, 145, 112430. [Google Scholar] [CrossRef]
- Ravikumar, B.; Vacher, C.; Berger, Z.; Davies, J.E.; Luo, S.; Oroz, L.G.; Scaravilli, F.; Easton, D.F.; Duden, R.; O’Kane, C.J.; et al. Inhibition of mTOR induces autophagy and reduces toxicity of polyglutamine expansions in fly and mouse models of Huntington disease. Nat. Genet. 2004, 36, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Malagelada, C.; Jin, Z.H.; Jackson-Lewis, V.; Przedborski, S.; Greene, L.A. Rapamycin protects against neuron death in in vitro and in vivo models of Parkinson’s disease. J. Neurosci. 2010, 30, 1166–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Rong, Y.; Ji, C.; Lv, C.; Jiang, D.; Ge, X.; Gong, F.; Tang, P.; Cai, W.; Liu, W.; et al. MicroRNA-421-3p-abundant small extracellular vesicles derived from M2 bone marrow-derived macrophages attenuate apoptosis and promote motor function recovery via inhibition of mTOR in spinal cord injury. J. Nanobiotechol. 2020, 18, 72. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Uezono, N.; Yasui, T.; Nakajo, M.; Nagai, T.; Wang, D.; Nishibori, M.; Nakashima, K. Combinatrial treatment of anti-High Mobility Group Box-1 monoclonal antibody and epothilone B improves functional recovery after spinal cord contusion injury. Neurosci. Res. 2021, 172, 13–25. [Google Scholar] [CrossRef]
- Ruschel, J.; Hellal, F.; Flynn, K.C.; Dupraz, S.; Elliott, D.A.; Tedeschi, A.; Bates, M.; Sliwinski, C.; Brook, G.; Dobrindt, K.; et al. Axonal regeneration. Systemic administration of epothilone B promotes axon regeneration after spinal cord injury. Science 2015, 348, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Bhowmick, S.; Abdul-Muneer, P.M. PTEN Blocking Stimulates Corticospinal and Raphespinal Axonal Regeneration and Promotes Functional Recovery After Spinal Cord Injury. J. Neuropathol. Exp. Neurol. 2021, 80, 169–181. [Google Scholar] [CrossRef]
- Ding, Z.; Dai, C.; Zhong, L.; Liu, R.; Gao, W.; Zhang, H.; Yin, Z. Neuregulin-1 converts reactive astrocytes toward oligodendrocyte lineage cells via upregulating the PI3K-AKT-mTOR pathway to repair spinal cord injury. Biomed. Pharmacother. 2021, 134, 111168. [Google Scholar] [CrossRef]
- Gaja-Capdevila, N.; Hernández, N.; Zamanillo, D.; Vela, J.M.; Merlos, M.; Navarro, X.; Herrando-Grabulosa, M. Neuroprotective Effects of Sigma 1 Receptor Ligands on Motoneuron Death after Spinal Root Injury in Mice. Int. J. Mol. Sci. 2021, 22, 6956. [Google Scholar] [CrossRef]
- Tanji, C.; Hashimoto, M.; Furuya, T.; Saito, J.; Miyamoto, T.; Koda, M. Sigma 1 receptor agonist cutamesine promotes plasticity of serotonergic boutons in lumbar enlargement in spinal cord injured rats. Neurosci. Lett. 2021, 759, 135971. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ji, J.; Wang, Z.; Xu, T. Effects of minocycline on motor function recovery and expression of glial fibrillary acidic protein and brain-derived neurotrophic factor after spinal cord injury in rats. J. Pharm. Pharmacol. 2021, 73, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Zavvarian, M.M.; Hong, J.; Khazaei, M.; Chio, J.C.T.; Wang, J.; Badner, A.; Fehlings, M.G. The Protein Kinase Inhibitor Midostaurin Improves Functional Neurological Recovery and Attenuates Inflammatory Changes Following Traumatic Cervical Spinal Cord Injury. Biomolecules 2021, 11, 972. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Joshi, H.P.; Sheen, S.H.; Kim, K.T.; Kyung, J.W.; Choi, H.; Kim, Y.W.; Kwon, S.Y.; Roh, E.J.; Choi, U.Y.; et al. Resolvin D3 Promotes Inflammatory Resolution, Neuroprotection, and Functional Recovery After Spinal Cord Injury. Mol. Neurobiol. 2020, 58, 424–438. [Google Scholar] [CrossRef]
- He, X.-J.; Wang, F.; Zhao, Y.-J.; Qiao, H.; Liu, D.-F.; Li, J.; Li, J.-X.; Chang, S.-E.; Lu, T.; Li, F.-T.; et al. Lithium promotes recovery after spinal cord injury. Neural. Regen. Res. 2022, 17, 1324–1333. [Google Scholar] [CrossRef]
- Ito, S.; Ozaki, T.; Morozumi, M.; Imagama, S.; Kadomatsu, K.; Sakamoto, K. Enoxaparin promotes functional recovery after spinal cord injury by antagonizing PTPRσ. Exp. Neurol. 2021, 340, 113679. [Google Scholar] [CrossRef]
- Invernizzi, M.; de Sire, A.; Fusco, N. Rethinking the clinical management of volumetric muscle loss in patients with spinal cord injury: Synergy among nutritional supplementation, pharmacotherapy, and rehabilitation. Curr. Opin. Pharmacol. 2021, 57, 132–139. [Google Scholar] [CrossRef]
- Otzel, D.M.; Kok, H.J.; Graham, Z.A.; Barton, E.R.; Yarrow, J.F. Pharmacologic approaches to prevent skeletal muscle atrophy after spinal cord injury. Curr. Opin. Pharmacol. 2021, 60, 193–199. [Google Scholar] [CrossRef]
- Weaver, F.M.; Etingen, B.; Guihan, M.; Ray, C.; Priebe, M.; Burns, S.; Carbone, L. Spinal cord injury providers’ perspectives on managing sublesional osteoporosis. J. Spinal Cord Med. 2020, 43, 428–434. [Google Scholar] [CrossRef]
- Abdelrahman, S.; Ireland, A.; Winter, E.M.; Purcell, M.; Coupaud, S. Osteoporosis after spinal cord injury: Aetiology, effects and therapeutic approaches. J. Musculoskelet. Neuronal. Interact. 2021, 21, 26–50. [Google Scholar]
- Holman, M.; Chang, G.; Ghatas, M.; Saha, P.; Zhang, X.; Khan, M.; Sima, A.; Adler, R.; Gorgey, A.S. Bone and non-contractile soft tissue changes following open kinetic chain resistance training and testosterone treatment in spinal cord injury: An exploratory study. Osteoporos. Int. 2021, 32, 1321–1332. [Google Scholar] [CrossRef]
- Ryu, Y.; Ogata, T.; Nagao, M.; Sawada, Y.; Nishimura, R.; Fujita, N. Early escitalopram administration as a preemptive treatment strategy against spasticity after contusive spinal cord injury in rats. Sci. Rep. 2021, 11, 7120. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Huang, Y.; Chen, R.; Liu, N.; Fang, S. Efficacy and safety of tolperisone versus baclofen among Chinese patients with spasticity associated with spinal cord injury: A non-randomized retrospective study. Braz. J. Med. Biol. Res. 2021, 54, 11293. [Google Scholar] [CrossRef] [PubMed]
- Štětkářová, I.; Krámská, L.; Keller, J. Improvement of Memory Functions in Chronic Spinal Cord Injury After Long-Term Intrathecal Baclofen Delivery for Spasticity Relief. Neuromodulation 2021, 24, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Delhaas, E.M.; Frankema, S.P.G.; Huygen, F.J.P.M. Intrathecal baclofen as emergency treatment alleviates severe intractable autonomic dysreflexia in cervical spinal cord injury. J. Spinal Cord Med. 2021, 44, 617–620. [Google Scholar] [CrossRef] [Green Version]
- Palazón-García, R.; Benavente-Valdepeñas, A.M. Botulinum Toxin: From Poison to Possible Treatment for Spasticity in Spinal Cord Injury. Int. J. Mol. Sci. 2021, 22, 4886. [Google Scholar] [CrossRef]
- Li, S.; Francisco, G.E. The Use of Botulinum Toxin for Treatment of Spasticity; Handbook of Experimental Pharmacology Book Series; Springer: Berlin/Heidelberg, Germany, 2021; Volume 263, p. 281. [Google Scholar]
- Krebs, J.; Pannek, J.; Rademacher, F.; Wöllner, J. Are 200 units of onabotulinumtoxin A sufficient for the suppression of neurogenic detrusor overactivity in individuals with established 300-unit botulinum toxin treatment? A retrospective cohort study. World J. Urol. 2021, 39, 543–547. [Google Scholar] [CrossRef]
- Heriseanu, R.E.; Chari, P. A severe case of non-infective myositis six weeks post intramuscular injections of Onabotulinum toxin A (Botox) in a young man with tetraplegia: Case report. Spinal Cord Ser. Cases 2021, 7, 76. [Google Scholar] [CrossRef]
- Albayrak, O.; Coşkun, Ö.K.; Giray, E.; Karadağ-Saygı, E. Botulinum toxin application to the internal and external oblique muscles for abdominal spasms in spinal cord injury. Spinal Cord Ser. Cases 2021, 7, 75. [Google Scholar] [CrossRef]
- Gong, C.; Zheng, X.; Guo, F.; Wang, Y.; Zhang, S.; Chen, J.; Sun, X.; Shah, S.Z.A.; Zheng, Y.; Li, X.; et al. Human spinal GABA neurons alleviate spasticity and improve locomotion in rats with spinal cord injury. Cell Rep. 2021, 34, 108889. [Google Scholar] [CrossRef]
- Sun, W.M.; Ma, C.L.; Xu, J.; He, J.P. Reduction in post-spinal cord injury spasticity by combination of peripheral nerve grafting and acidic fibroblast growth factor infusion in monkeys. J. Int. Med. Res. 2021, 49, 3000605211022294. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mech, D.; Korgol, K.; Kurowska, A.; Adamski, B.; Miazga, M.; Biala, G.; Kruk-Slomka, M. Promising Advances in Pharmacotherapy for Patients with Spinal Cord Injury—A Review of Studies Performed In Vivo with Modern Drugs. J. Clin. Med. 2022, 11, 6685. https://doi.org/10.3390/jcm11226685
Mech D, Korgol K, Kurowska A, Adamski B, Miazga M, Biala G, Kruk-Slomka M. Promising Advances in Pharmacotherapy for Patients with Spinal Cord Injury—A Review of Studies Performed In Vivo with Modern Drugs. Journal of Clinical Medicine. 2022; 11(22):6685. https://doi.org/10.3390/jcm11226685
Chicago/Turabian StyleMech, Dominika, Katarzyna Korgol, Antonina Kurowska, Bartlomiej Adamski, Malgorzata Miazga, Grazyna Biala, and Marta Kruk-Slomka. 2022. "Promising Advances in Pharmacotherapy for Patients with Spinal Cord Injury—A Review of Studies Performed In Vivo with Modern Drugs" Journal of Clinical Medicine 11, no. 22: 6685. https://doi.org/10.3390/jcm11226685